
Abstract
Skeletal muscle stem cell (SMSC) transplantation has shown great therapeutical potential in repairing muscle loss and dysfunction, but the muscle acquisition is usually a traumatic procedure causing pain and morbidity to the donor. In this study, we investigated the feasibility of isolating SMSCs from human orbicularis oculi muscle (OOM), which is routinely removed and discarded during ophthalmic cosmetic surgeries. OOM fragments were harvested from 18 female healthy donors undergoing upper eyelid plasties. Plastic-adherent cells were isolated from the muscles using a two-step plating method combined with collagenase digestion. A total of 15 cell cultures were successfully established from the muscle samples. These adherent cells were positive for the specific markers of SMSCs and could be directed toward the osteogenic, adipogenic, chondrogenic, and myogenic phenotypes in the presence of lineage-specific inductive media. Moreover, after cultured in the myogenic inductive medium for 3 weeks, the muscle cells were injected into the tibialis anterior muscles of nude mice and the cell fate was detected using a DiI-labeling technique. In vivo myogenesis was evidenced by the expression of DiI fluorescence after cell transplantation. The donor cells could be found in the satellite cell position and incorporated into the host myofibers. Our results demonstrated that human OOM represents a novel source of myogenic precursors with stem cell-like properties, which may provide a foundation for the SMSC-based therapeutics of skeletal muscle diseases.
Impact Statement
In the present study we obtained an adherent cell population of skeletal muscle stem cells (SMSCs) from human orbicularis oculi muscle (OOM) samples using a two-step plating method combined with collagenase digestion, which possess the self-renewal and multipotential capabilities in vitro. Moreover, these cells could differentiate into myoblasts and fuse into the host myofibers when injected into a nude mouse model of muscle regeneration, suggesting that human OOM tissue may represent a novel SMSC source for the cellular therapeutics of skeletal muscle regeneration.
Introduction
T
However, there is a progressive depletion of the SMSC population in degenerative muscle diseases, and thus, SC recovery is low from dystrophic muscles. 11 Moreover, the muscle biopsy acquisition is an invasive procedure usually causing pain, bleeding, infection, and morbidity to the donor site, limiting the availability of muscles that can be obtained. Therefore, it is difficult to isolate sufficient SMSCs from donors and in some studies these cells had to be harvested from human traumatized muscle tissues.12,13
Upper eyelid plasty is the most performed cosmetic and ophthalmic surgery in China. During this operation, some orbicularis oculi muscle (OOM) tissue will be normally removed and discarded as waste. OOM is the main sphincter muscle of human eyelids. Its function is to modulate the facial expression and close the palpebral fissure. 14 As a highly specialized skeletal muscle, OOM might represent a valuable source for SMSCs due to its rich vascularization and easy accessibility. Compared with the limb and trunk muscles, OOM has two distinct advantages. First, it can be spared in the DMD patients due to different excitation–contraction coupling mechanisms and low expression levels of dystrophin. 15 Second, it is more easily harvested with less morbidity and discomfort to donors. To date, however, few literature have reported the existence of SMSCs in human OOM.
In the present study, we isolated an adherent cell population from human OOM using a two-step plating method combined with collagenase digestion. In vitro, these muscle cells could differentiate into the osteogenic, chondrogenic, adipogenic, and myogenic lineages. After introduced into muscles of nude mice, they were found to be able to participate in myofiber regeneration. Our data demonstrated that human OOM is an easy accessible and less invasive source of muscle stem cells, which may have potential application for the cellular therapeutics of muscle dysfunction.
Materials and Methods
SMSC isolation and expansion
Experimental protocols for our study were approved by the Research Ethics Committee of Tongji University and conformed to the principles outlined in the Declaration of Helsinki. A total of 18 female healthy patients (18–51 years old, mean age 28.6 ± 10.4 years) who underwent upper eyelid blepharoplasty were enrolled in this study. They all had provided written informed consent and did not present any eyelid or orbital diseases. Muscle fragments were surgically removed from the pretarsal portion of OOM during the procedure. After histological and immunohistochemical evaluation, OOM samples were processed according to methods previously reported, with modification.2,6 First, muscle tissues were extensively washed in phosphate-buffered solution (PBS; Sigma, Shanghai, China), minced into a coarse slurry using sterile scissors, and digested with 0.1% collagenase type I solution (Worthington Biochemical Corp., Lakewood, NJ) at 37°C for 4 h under constant shaking. The enzymatic digestion was neutralized with low-glucose Dulbecco's modified Eagle's medium (LG-DMEM; Gibco, Grand Island, NY) containing 10% fetal bovine serum (FBS; Hyclone Laboratories, Logan, UT).
Digested muscles were centrifuged at 250 g for 10 min and the resulting cell pellets were resuspended in the growth medium (GM, containing LG-DMEM, 10% FBS, 2 mM
Cell identification and in vitro multilineage differentiation
Flow cytometric analysis was performed as previously reported to characterize the phenotype of the OOM-derived adherent cells. 16 A total of 105 cells were digested with trypsin-EDTA, washed with PBS, and incubated at 4°C for 30 min with the following phycoerythrin (PE)-conjugated or fluorescein isothiocyanate (FITC)-conjugated monoclonal anti-human antibodies: CD45-PE, CD73-PE, HLA-ABC-PE, HLA-DR-PE, CD31-FITC, CD29-FITC, CD34-FITC, CD56-FITC, CD90-FITC, and CD105-FITC (antibodies from Abcam, Cambridge, MA). Immunolabeled cells were analyzed by flow cytometry (FACSCalibur; Becton Dickson, San Jose, CA), and the cells incubated with PE- or FITC-conjugated nonspecific immunoglobulin G were used as isotype controls.
The multipotential differentiation capabilities of the muscle cells were evaluated as follows with methods previously reported.16,17 To induce osteogenic differentiation, cells were plated at a density of 2 × 104/cm2 in 35-mm dishes and induced for 2 weeks in the osteogenic medium (consisting of GM, 0.1 μM dexamethasone, 50 μM ascorbic acid, and 10 mM β-phosphoglycerol [all from Sigma]). Half media were replaced twice a week and the osteogenic differentiation potential was assessed by Alizarin red staining (Sigma). For adipogenic differentiation, cells were similarly replated and maintained for 2 weeks in the adipogenic medium (consisting of GM, 0.5 mM isobutyl-methylxanthine, 1 μM dexamethasone, 100 μM indomethacin, and10 μg/mL insulin [all from Sigma]). Medium was replaced every 3 days, and intracellular lipid accumulation was analyzed by Oil red staining (Sigma) after 2 weeks. For chondrogenic differentiation, cells were collected and spun to pellets (2.5 × 106 per pellet) in 15-mL polypropylene tubes (Falcon) and cultured in the chondrogenic medium (containing high-glucose DMEM, 10% FBS, 100 ng/mL insulin-like growth factor, 10 ng/mL transforming growth factor-β1, 6.25 mg/mL transferrin, and 40 ng/mL dexamethasone [all from Sigma except for FBS from HyClone]). Half media were changed twice weekly. After 3 weeks, the chondrogenic differentiation was determined by Toluidine blue staining (Sigma) to detect the extracellular accumulation of chondrocyte matrix.
In vitro myogenic differentiation
The myogenic differentiation of OOM-derived cells was performed as described previously.1,16 Briefly, cells were seeded at 2 × 104/cm2 in 35-mm dishes containing glass coverslips, which had been pretreated with 1:200 PBS-diluted (50 μL/cm2) Matrigel™ (BD Biosciences, Franklin Lakes, NJ) at 37°C overnight before cell plating, and maintained in fresh myogenic medium (containing GM, 5% horse serum, 50 μM hydrocortisone, and 0.1 μM dexamethasone [all from Sigma]) with changes twice weekly. After 2 weeks, myogenic differentiation was evaluated by fluorescent immunocytostaining with Pax3, Pax7, Myf5, and MyoD (early myogenic markers, antibodies from BD Sciences). At 4 weeks after induction, formation of myotubes, Desmin and Myogenin (late myogenic markers, antibodies from Santa Cruz Biotechnology, Santa Cruz, CA), was evaluated by microscopy, immunofluorescence, and immunocytochemical staining separately. The number of positively stained cells was expressed as a percentage of the total cells counted within the images. Cells incubated in GM were used as controls.
In vivo myogenic differentiation
After myogenically induced for 3 weeks, the OOM-derived cells were trypsinized and labeled by the CM-DiI Staining Kit (Yeasen Biotech, Shanghai, China). Then, injections of 2.0 × 105 cells suspended in 20 μL of PBS were performed into the right tibialis anterior muscles of nude mice (8 weeks old). Noninduced DiI-stained cells were transplanted into the left muscles as the control group. After 1 week, the animals were killed and muscles that received injections were analyzed by the standard histology (hematoxylin and eosin staining) and the confocal laser scanning microscopy examination (Leica Microsystems, Mannheim, Germany) observation. The number of DiI-staining-positive myofibers was counted per vision field (200 × ) with an average value of four sections/sample. The donor cell recovery was defined as the number of DiI-positive myofibers per 105 transplanted cells.18,19 Animals used in this study were processed according to protocols approved by the Institutional Animal Care and Use Committee of Tongji University.
Results
SMSC isolation and expansion
Human OOM tissue can be divided into three parts: the pretarsal, preseptal, and orbital portions (Fig. 1a). On an average, 0.484 ± 0.266 g of muscle fragments was harvested from the pretarsal portion in each operation (n = 18). Histological evaluation demonstrated the typical structure of skeletal muscle myofibers accompanied by few fibrous tissues (Fig. 1b, c). A total of 15 cell cultures were successfully established from the 18 samples. Cells could be isolated from the muscle fibers on the basis of collagenase dissociation and slow adhesion characteristics. After 7 days in primary culture, these cells positively expressed the SC markers of Pax7 and Myf5 (Fig. 1d, e). The mean cell yield in primary culture was (2.68 ± 0.83) × 105 cells per gram of OOM (n = 15).
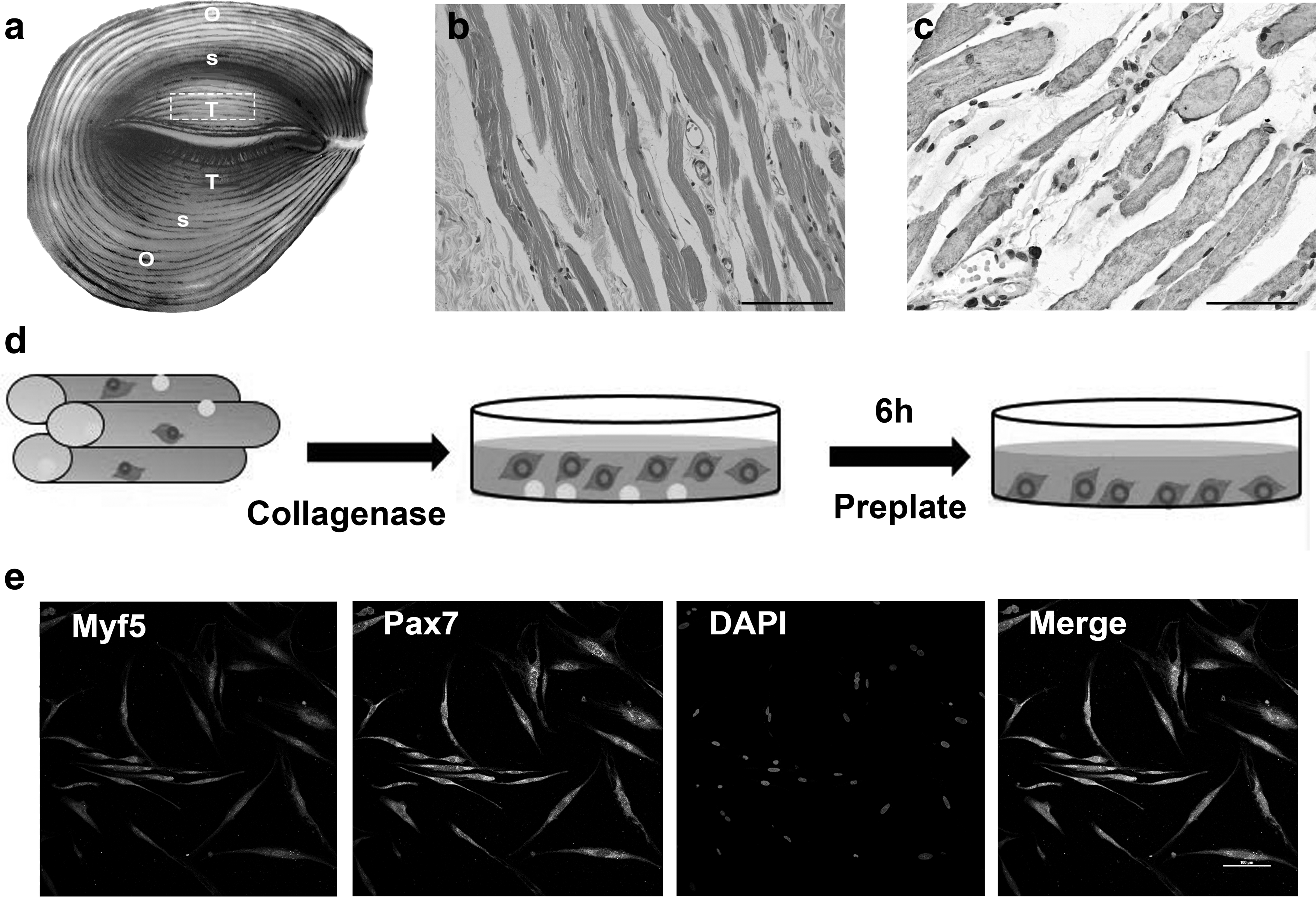
Isolation of OOM-derived SMSCs.
Cell identification and in vitro multilineage differentiation
The adherent cells primarily obtained from OOM with the two-step plating technique were of bipolar spindle shape (Fig. 2a) and could form big colonies when cultured in vitro for 2 weeks (Fig. 2b). After passaging, cells showed homogenously fibroblast-like morphology (Fig. 2c). The subcultured cells could be maintained in vitro for at least 10 passages with a mean population doubling time of 60–70 h (data not shown). Capabilities of multipotential differentiation were ascertained by the positive Alizarin red staining (osteogenic marker, Fig. 2d), Toluidine blue staining (chondrogenic marker, Fig. 2e), and Oil red staining (adipogenic marker, Fig. 2f) separately. No positive results were revealed in the noninduced control cells (data not shown).
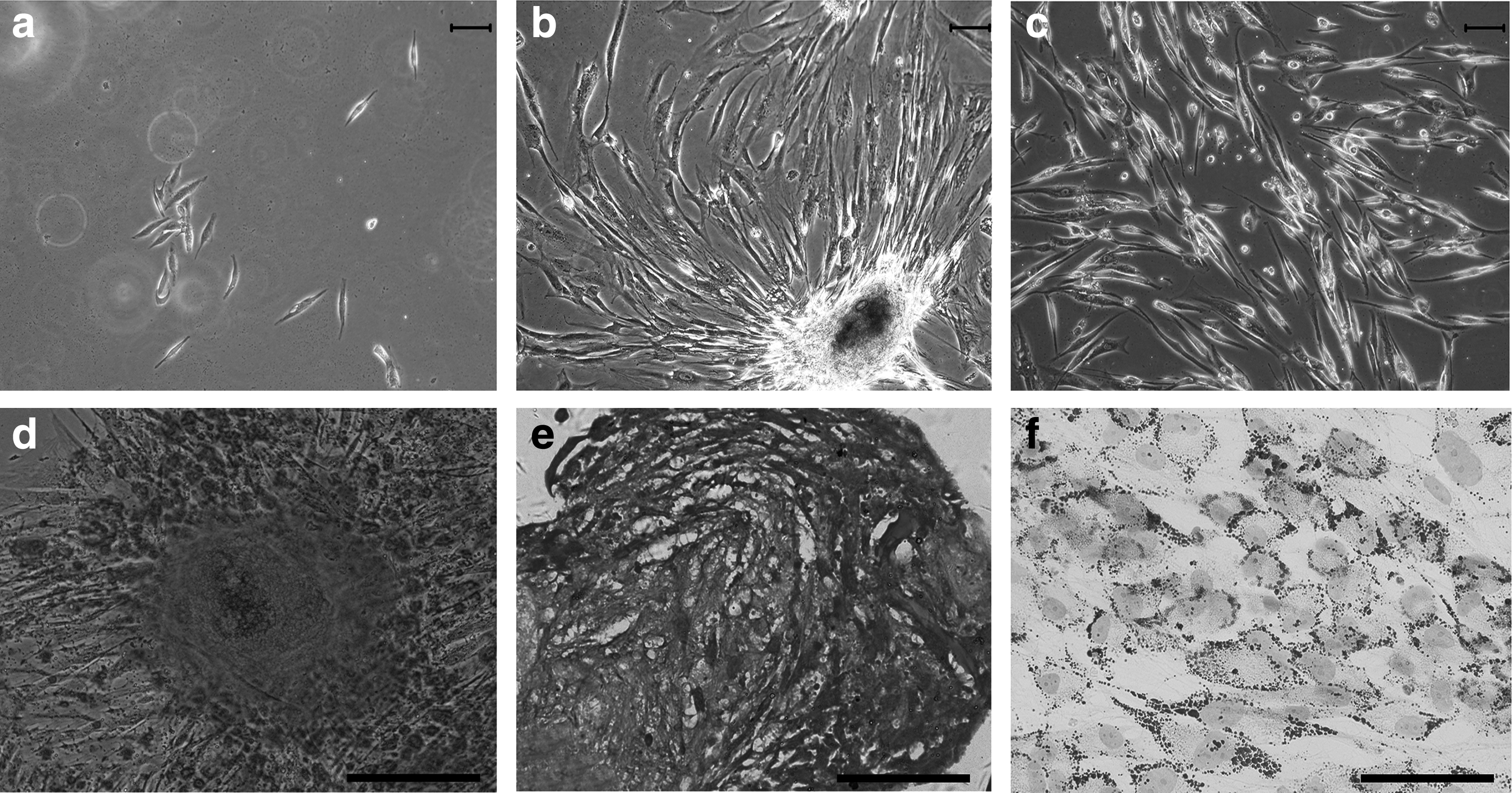
In vitro proliferation and multilineage differentiation of SMSCs.
Flow cytometric characterization demonstrated that the muscle cells expressed the SMSC-specific markers of CD56 and CD34, as well as the mesenchymal stem cell (MSC) markers of CD73, CD90, CD29, and CD105. On the contrary, these cells were negative for the endothelial marker of CD31 or the monocyte/macrophage marker of CD45. 14 Furthermore, they were positive for HLA-ABC but not for HLA-DR (Fig. 3).

Immunophenotype of OOM-derived SMSCs. Cells were labeled with FITC- or PE-conjugated CD markers and analyzed by flow cytometry. Values represent the mean percentage ± standard deviation of the cells positively stained by the indicated markers (n = 3). PE, phycoerythrin; FITC, fluorescein isothiocyanate.
In vitro myogenic differentiation
After 14 days of in vitro myogenic culture, more than 80% of the induced cells were positive for the expression of Pax3, Pax7, Myf5, and MyoD, four early myogenic markers, exhibiting their differentiation capacity toward skeletal muscle precursor cells. However, the noninduced control cells did not show any positive results (Fig. 4a). At 4 weeks of induction, these cells fused into multinucleated myotubes and appeared immunoreactive for desmin and myogenin (terminal myogenic markers). In contrast, no formation of myotubes or positive expression of desmin and myogenin was found in the undifferentiated cells (Fig. 4b).
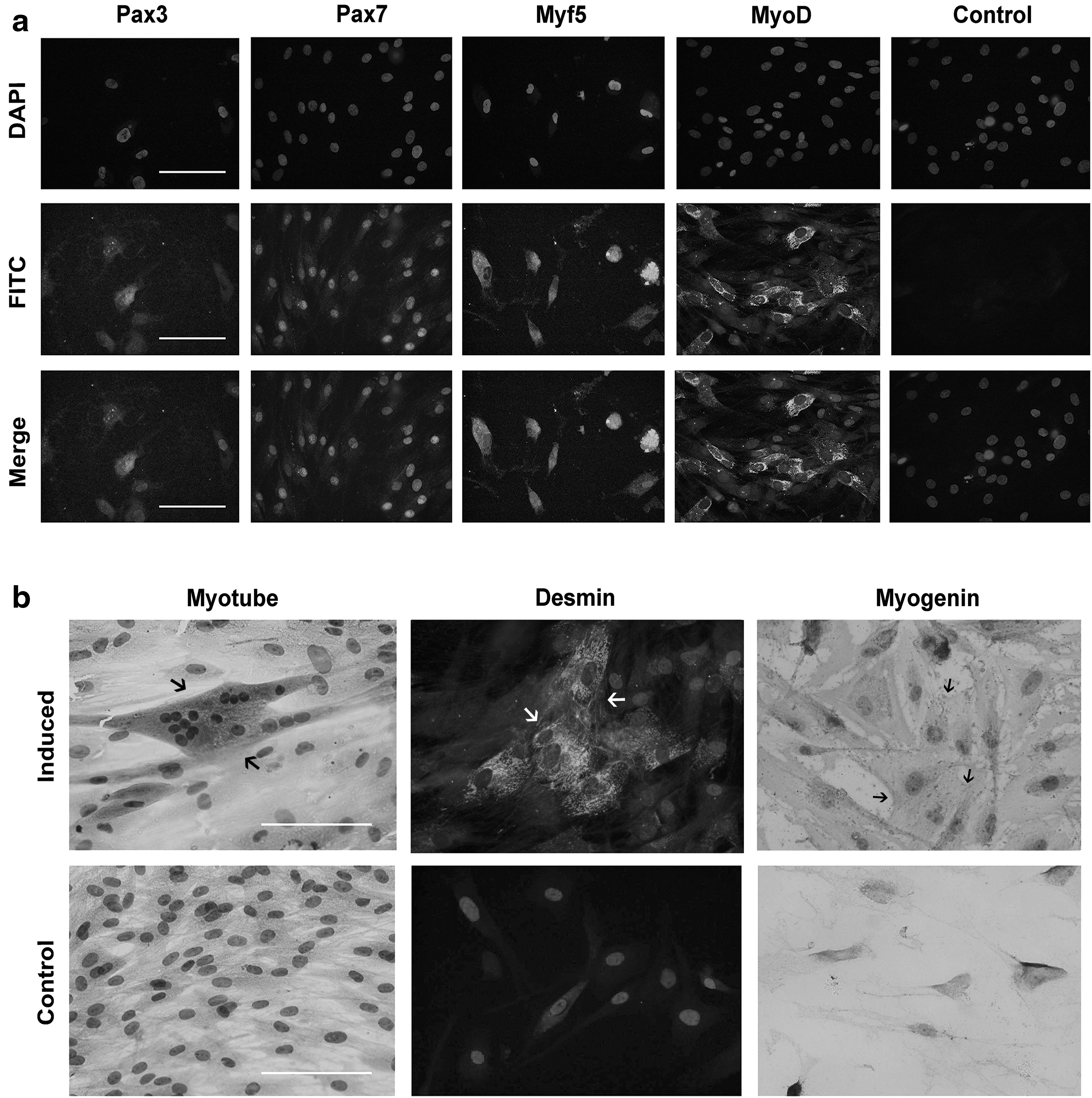
In vitro myogenic differentiation of SMSCs.
In vivo myogenic differentiation
The OOM-derived cells were myogenically induced for 3 weeks in vitro, labeled with CM-DiI stain, and injected into a nude mouse model of muscle regeneration. At 1 week posttransplantation, cells expressing the red fluorescent marker were found beneath the myofiber membrane, a position of SCs, and fused with the host muscular fascicles, indicating that the transplanted donor cells differentiated into myoblasts and incorporated into the pre-existing recipient myofibers undergoing regeneration (Fig. 5). And the in vivo cell recovery, obtained by counting the number of DiI-positive myofibers per 1 × 105 injected cells, was about 20.6 ± 4.3 (n = 3). In the control group, formation and regeneration of new myofibers were also observed in the muscles that received the cell injection, but no evidence of red fluorescence was detected.
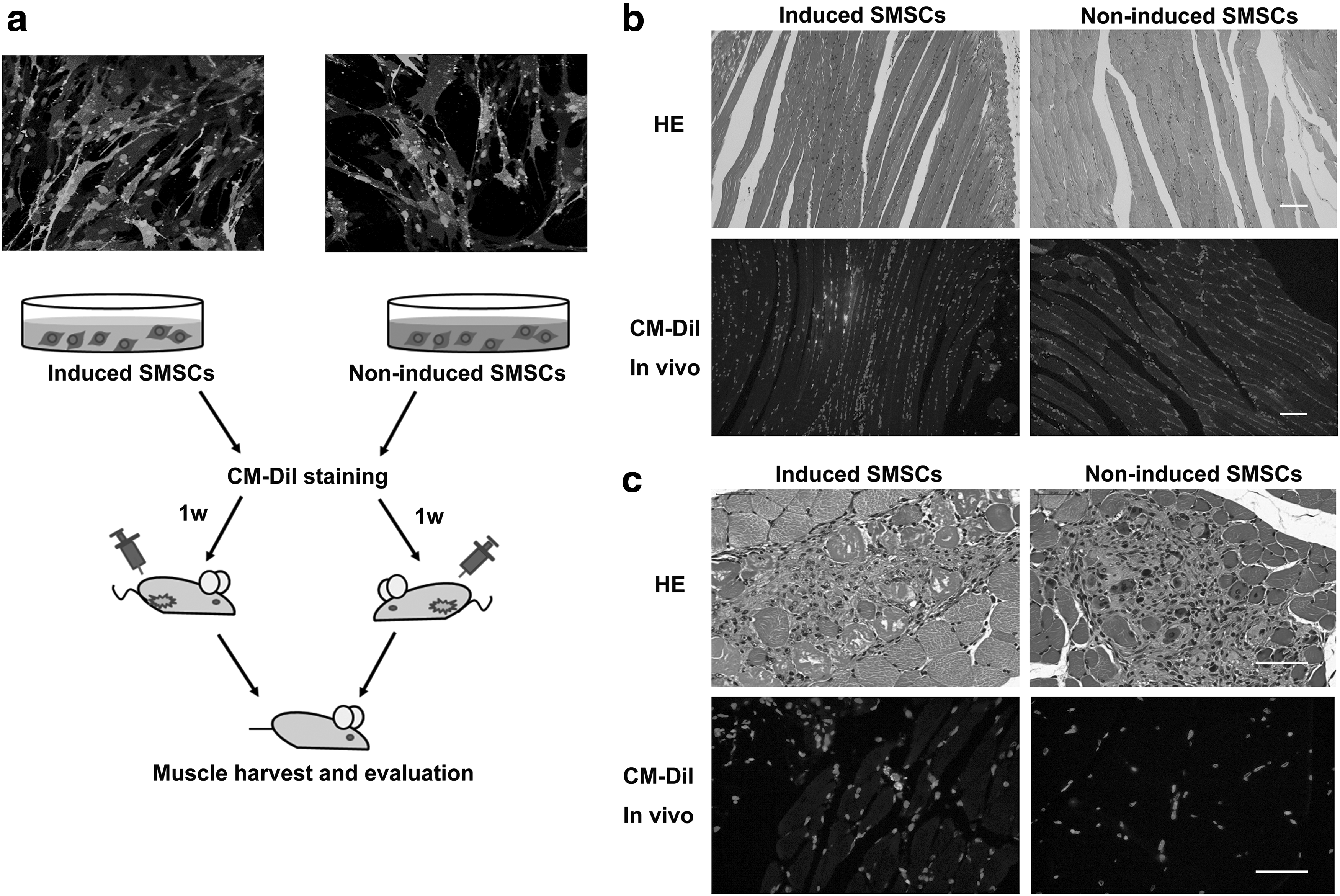
In vivo myogenic differentiation of SMSCs.
Discussion
The trunk and limb muscles have been characterized as a reservoir of SMSCs with the therapeutical potential to repair muscle loss or treat muscle dysfunction. Recently, some studies have reported the presence of SMSCs in craniofacial skeletal muscles, such as the orbicularis oris muscle and the masseter muscle.17,20 In this study, we described for the first time the existence of SMSCs in human OOM tissue, which can differentiate into myogenic progenitors in vitro and participate in muscle regeneration in vivo.
The OOM-derived cells were selected on the basis of high adhesion properties after enzymatic dissociation. Their myogenic origin was revealed by the positive expression of Myf5 and Pax7, two specific SC markers. 21 This cell population exhibited the multipotential capabilities of differentiating into adipogenic, osteogenic, and chondrogenic cells when exposed to lineage-specific inductive medium. The myoblast differentiation was further evidenced by the expression of early myogenic markers and terminal myotube-like appearance in the presence of myogenic induction medium. Moreover, the myogenically induced cells could take part in the new myofiber formation when transplanted into muscles of a nude mouse model, indicating their potential application in myogenic regenerative therapies.
Muscle stem cells are heterogeneous in nature. To isolate and purify the cells, the preplate technique is normally recommended.2,22 In our present study, a two-step plating method was applied to obtain SMSCs from human OOM and it has proven to be very effective. Most muscle cells that could adhere to the tissue culture plastic rapidly were fibroblasts, while the slow-adhering cells were mainly myogenic progenitors with SMSC characteristics.22–24 Therefore, we harvested the OOM-derived cells that were able to adhere to the plastic substrate from 6 to 24 h after initial plating. These cells demonstrated homogenous fibroblast-like morphology and clonogenic growth behavior, and expressed the SMSCs' characteristic markers (CD56 and CD34). They were also positive for the MSC antigen markers (CD73, CD90, CD29, and CD105) and negative for CD31 and CD45, thus excluding the contamination of leukocytes or endothelial cells.
Webster et al. reported that there are 105 SMSCs in 1 g of skeletal muscle tissue. 25 In this study, we found that the stem cell number within OOM is more abundant. Approximately (2.68 ± 0.83) × 105 adherent cells with capacities of in vitro self-renewal and proliferation could be harvested from one gram of human OOM. They were able to expand to at least 10 passages in culture, yielding a cell number sufficient for the stem cell-based therapeutics for skeletal muscle diseases.26–28 Then, the functional myogenic capacity of SMSCs was verified in in vivo animal experiments. The mouse model used as human cell recipient in our study was immunodeficient, but the muscles were not predamaged with a myonecrotic agent or preirradiated before cell injection. It is reported that the muscle-injected myoblasts only fused with myofibers that underwent regeneration or repair.24,28 So, we speculate that the intramuscularly transplanted SMSCs mainly fuse with the myofibers injured by the injection needle. And our results confirmed the incorporation of donor cells with new myofibers by the presence of labeled fluorescence.
Adipose-derived stem cells (ASCs), a population of adult stem cells existing abundantly in human subcutaneous adipose tissue, also possess the myogenic differentiation capability.1,11 Therefore, it is necessary to take measures to avoid ASC interference when confirming the SMSC origin in this work. First, eyelid skin is the thinnest skin of the body and unique in having no subcutaneous fat layer. 29 Second, OOM is composed of three portions (the pretarsal, preseptal, and orbital parts), and each part varies in the histological structure indicating different physiological functions. 14 The pretarsal part of OOM tissue contains the highest percentage of skeletal muscle (83.5%) with no fatty tissue (0%). The preseptal portion consists of less muscle content (46.5%) and more adipose tissue (12.7%). The highest adiposity (32.7%) appears in the orbital orbicularis portion. 14 As the skin layer had been discarded when preparing OOM samples and the muscle samples used in our study were removed from the OOM pretarsal part where there is no fat tissue (Fig. 1), contamination of ASCs with SMSCs could be precluded.
In summary, our present study suggested that SMSCs can be easily accessible from OOM tissues, which are routinely removed and discarded during the eyelid cosmetic surgeries, allowing SMSC isolation without the need of additional surgical procedures and representing a less invasive source of muscle stem cells to be applied in myogenic regenerative studies.
Footnotes
Acknowledgment
This work was supported financially by a grant from the National Science Foundation of China (31271027).
Disclosure Statement
No competing financial interests exist.