
Abstract
Triggering receptor expressed on myeloid cells-2 (TREM-2), a transmembrane receptor expressed by macrophages, microglia, and osteoclasts (OCs), plays a protective role in late-onset Alzheimer Disease (AD). To validate TREM-2 as a therapeutic target in AD, its potential secondary parallel effect on bone homeostasis should be clarified. However, animal models and monolayer cultures of human cells were shown poorly predictive of TREM-2 function in human. Therefore, this study aimed to engineer a tridimensional in vitro model using human progenitors differentiated into osteoblasts and OCs, recapitulating physiological bone homeostasis. Human bone marrow-derived mesenchymal cells were seeded and cultured under perfusion inside a collagen type I scaffold for 3 weeks, generating osteoblasts and mineralized matrix. Human peripheral blood-derived CD14+ monocytes were subsequently seeded through the generated tissue, thanks to perfusion flow, and further cultured for up to 3 weeks with an inductive medium, generating mature OCs. This culture system supported collagenous matrix deposition and resorption, allowing for the investigation of kinetic of soluble TREM-2 over the coculture time. Agonistic activation of TREM-2 in this model had no effect on OC activity or on mineralized matrix turnover. In conclusion, the engineered culture system represents a tridimensional, in vitro human bone model for drug testing and suggested no effect of TREM-2 agonist on bone resorption.
Introduction
T
TREM-2 is also a regulator of osteoclastogenesis and bone homeostasis. 12 Complete loss-of-function of TREM-2 results in a rare syndrome called Nasu-Hakola disease, characterized by bone cysts and presenile dementia. 13 In such patients, osteoclasts (OCs) differentiation is arrested, resulting in large aggregates of immature OCs with impaired bone resorption activity. 14 Knock-down of TREM-2 impairs proliferation and beta-catenin activation in OC precursors in vitro. 12 Through a cross-talk between osteoblastic and OC cells, osteoclastogenesis requires functional TREM-2, which cooperates with the Receptor Activator for κB factor (RANK).
In contrast to the numerous loss-of-function studies available, it is currently unknown whether and how gain-of-function of TREM-2 affects bone homeostasis. This is likely related to the fact that the nature of endogenous TREM-2 ligand(s) is still elusive5,15 and the physiological role of its shed form (sTREM-2) still needs to be elucidated.16,17 However, considering the therapeutic strategy to agonize TREM-2-mediated signaling for neuroprotection, it would be of highest relevance to identify TREM-2 ligands and assess the effect of TREM-2 overactivation on the differentiation of myeloid precursors into functional multinucleated OCs and on bone homeostasis. While targeting TREM-2 may provide new opportunities for AD diagnosis and treatment, it is relevant to assess its possible effect on bone homeostasis. On the contrary, the study of TREM-2 pathway is mainly relying on in vivo mouse model or 2D cell culture, 6 which hampers a deeper understanding of its role in human bone homeostasis.
To investigate the underlying mechanisms of physiological and pathological cellular processes in bone, animal models show limitations. In addition to the high costs and large degree of systemic complexity, they do not faithfully recapitulate the biological programs specific of the human species and are poorly predictive of a clinical outcome.3,12,18 In fact, while human TREM-2 activates osteoclastogenesis, mouse TREM-2 has an inhibitory effect on the formation of OCs. 15
Three dimensional organotypic culture models based on human cells may provide an alternative solution to this issue, reducing the use of complex and costly animal models, while providing a more physiological outcome. Several in vitro models have been proposed to recapitulate the process of bone turnover by using human progenitors19,20 and mimic relevant pathological states with the ultimate goal of providing systems for drug testing. 21
We thus aimed at establishing a perfusion-based three-dimensional (3D) model using human progenitors differentiated into OBs and OCs. Bone marrow-derived mesenchymal cells (BMCs) were cultured and differentiated into OBs under perfusion in collagen type I scaffolds to achieve bone-like matrix deposition. In a second phase, human monocytes were added to the system and differentiated into OCs to establish a coculture system. Extracellular matrix (ECM) turnover was monitored over time together with OCs activity in a homeostatic state. We then tested the hypothesis that the effect of TREM-2 agonistic stimulation on bone turnover and OC activity could be tested in this model.
Materials and Methods
Cell source and expansion
Bone marrow aspirates were obtained from the iliac crest of healthy donors during routine orthopedic surgical procedures, after informed consent from the patient and following protocol approval by the local Ethics Committee (Ethical Kommission Beider Basel #78/07). Nucleated cells were isolated from aspirates by means of red blood cells lysis buffer (pH 7.2) containing 0.15 M NH4Cl, 1 mM KHCO3 (Sigma Aldrich), and 0.1 mM Na2EDTA (Fluka, Switzerland). Freshly isolated cells were plated at a density of 1 × 104 nucleated cells/cm2. Cell expansion was carried out in DAF medium, which consisted of α-modified Eagle's medium (α-MEM), 10% fetal bovine serum, 100 mM HEPES buffer solution, 1 mM sodium pyruvate, 100 U/mL penicillin, 100 mg/mL streptomycin, and 292 mg/mL L-glutamine (GIBCO, Switzerland), supplemented with 100 nM dexamethasone, 0.1 mM L-ascorbic acid-2-phosphate (both from Sigma Aldrich), and 5 ng/mL fibroblast growth factor-2 (FGF-2; R&D systems). DAF medium was used to increase human bone marrow mesenchymal cells (hBMCs) proliferation and promote osteogenic commitment. 19 Medium was changed twice a week. At confluence, hBMCs were replated for expansion (seeding density of 3 × 103 cells/cm2). Upon confluence, cells at passage 2 were enzymatically retrieved and counted for use in the 3D coculture experiments.
Peripheral blood-derived CD14+ monocytes
Mononuclear cells were isolated from human peripheral blood buffy coats from healthy donors (different from bone marrow donors) by gradient centrifugation (Ficoll, Histopaque 1077; Sigma-Aldrich, St. Louis, MO). Subsequently, CD14+ monocytes were isolated using anti-CD14-coated magnetic beads (Miltenyi Biotec, Auburn, CA), according to the manufacturer's instructions. The freshly isolated CD14+ monocytes from different donors have been used for the 2D and 3D experiments.
3D coculture system
3D cocultures were established by using a perfusion bioreactor system (CELLEC Biotek AG), previously tested for cell seeding and cultivation into 3D scaffolds,19,22 following a two-phase strategy, as shown in Figure 1. In the first phase, 3 × 106 hBMCs were suspended in 8 mL of a-MEM complete medium (CM) supplemented with 10% fetal bovine serum, 100 mM HEPES buffer solution, 1 mM sodium pyruvate, 100 U/mL penicillin, 100 mg/mL streptomycin, and 292 mg/mL L-glutamine, and perfused through 6 mm diameter, 1.5 mm thick disks of porous, cross-linked collagen type I scaffolds (Optimax®; CELLEC Biotec AG). hBMCs were perfused overnight through the scaffolds at a superficial velocity of 1 mm/s. After 24 h (cell seeding phase), the superficial velocity was reduced to 100 μm/s for perfusion culture. hBMCs were cultured for 3 weeks with an osteoinductive medium (OM), which consists in a-MEM CM supplemented with 10 nM dexamethasone, 0.1 mM L-ascorbic acid-2-phosphate, and 10 mM β-glycerophosphate. In the second phase, 3 × 106 freshly isolated CD14+ monocytes were seeded into each osteogenic construct at a superficial velocity of 400 μm/s. Cells were then cultured for 3 additional weeks in the presence of CM supplemented with 25 ng/mL macrophage colony-stimulating factor (MCSF) to promote adhesion and proliferation of monocytes and 50 ng/mL receptor activator of nuclear factor kappa-B ligand (RANKL) (both from R&D systems, Minneapolis, MN) to promote OCs differentiation (osteoclastogenic medium). Medium was refreshed every 4 days and cells were cultured for an additional period of 21 days at a superficial velocity of 100 μm/s (the total culture time for phases 1 and 2 was 42 days).
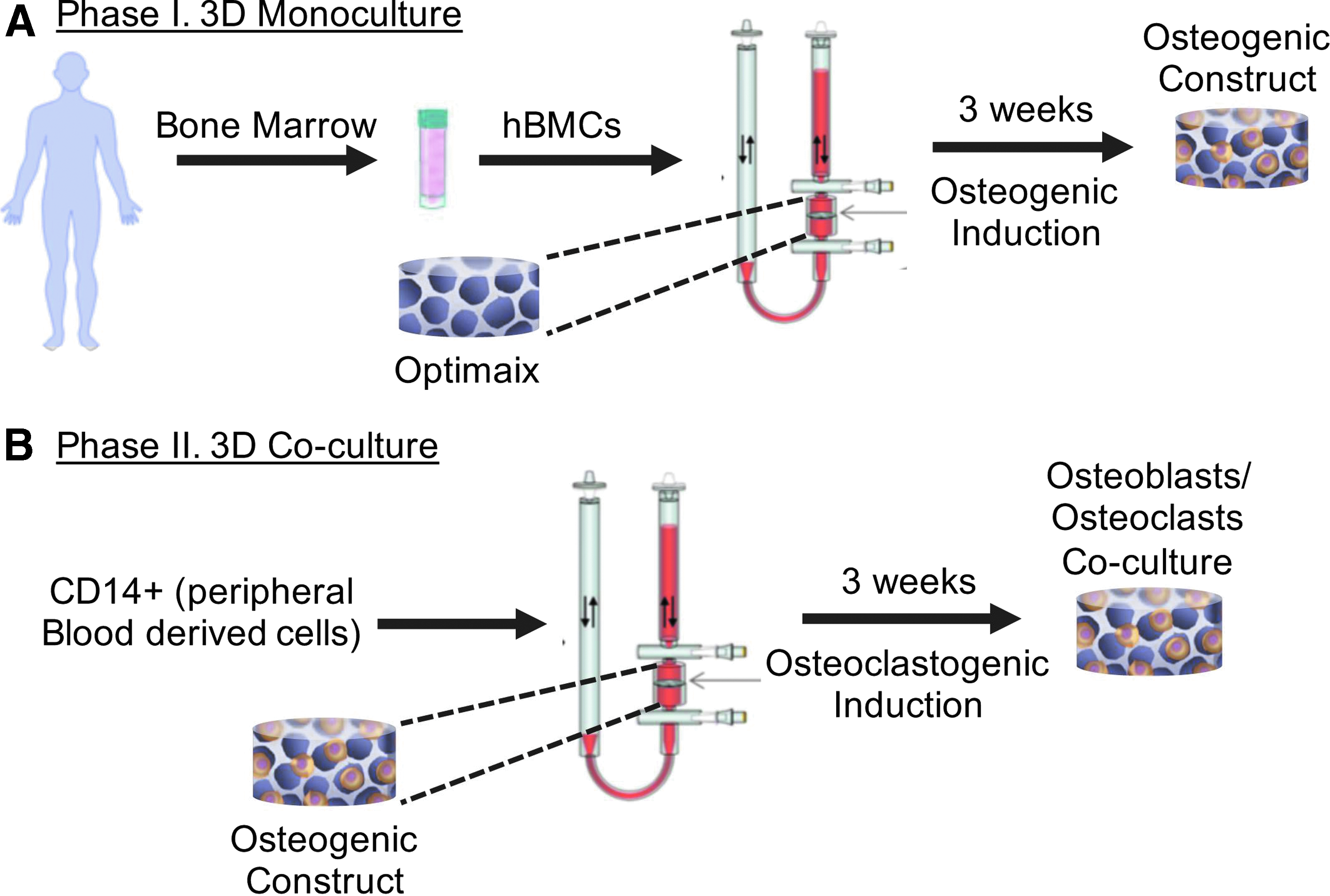
3D coculture system. hBMCs seeding and differentiation in osteogenic medium
In vitro functionality of the 3D coculture system
Histological staining and immunohistochemistry
After in vitro culture, constructs were fixed in 4% paraformaldehyde and embedded in paraffin. Eight micrometer-thick sections were stained with hematoxylin and eosin (H&E) (J.T. Baker/Avantor, Philipsburg, NJ), Alizarin red (AppliChem), Masson's trichrome (RAL Diagnostics), and for tartrate-resistant acid phosphatase (TRAP) activity by using a leukocyte acid phosphatase kit (Sigma-Aldrich). Expression of human bone sialoprotein (BSP) was assessed by immunohistochemistry using an anti-human BSP primary (Abcam; #52128) and biotinylated secondary antibodies (Dako; #E 0432). Samples were then incubated with a Vectastain ABC kits (Vector Labs, Burlingame, CA). The red signal was developed with the Fast Red kit (Dako Cytomation, Glostrup, Denmark) and sections counterstained by hematoxylin. Histological sections were analyzed with an Olympus BX-63 microscope (Olympus, Tokyo, Japan).
Cytofluorimetric analysis
At each time point of the secondary culture phase, cells were retrieved by perfusing the constructs with a solution of 0.3% collagenase (Worthington, NJ) for 40 min at 400 μm/s. Constructs were then perfused for an additional 15 min with a solution of 0.05% trypsin/0.53 mM EDTA (Gibco). The fraction of dead cells was assessed by Trypan blue exclusion (Sigma, Switzerland). After retrieval, cells were rinsed and incubated at 4°C for 30 min with antibodies against human CD14 and TREM-2 (BD Bioscience). Cells were then analyzed with an Accuri flow cytometer (BD Bioscience).
Supernatant analysis
Before each medium change, the media were collected, centrifuged at 400 g for 5 min and the resulting supernatant was stored at −20°C for enzyme-linked immunosorbent assay (ELISA) analysis. The samples were then thawed and assessed for C-terminus procollagen type I (CICP), cross-linked N-telopeptides of collagen type I (NTx-I, Osteomark® NTx Serum), (both from TECOmedical Group, Sissach, Switzerland), TRAP isoform 5b (TRAP5b, BoneTRAP® Immunodiagnostic Systems, Boldon Business Park, United Kingdom), shed TREM-2 (Roche Pharmaceuticals, Basel, Switzerland) and phosphate levels (BioVision). Each assay was performed according to the manufacturers' instructions.
Validation of TREM-2 activation in transfected HEK293 cells
Generation of TREM-2 expressing HEK293 cell clones
To verify TREM-2-activating effect of the tool antibody, HEK293 cells were stably transfected with human TREM-2 cDNA (NP_061838 VersaClone cDNA; R&D) and human Dap12 cDNA (Origene) using pBud CE4.1 double-gene expression vector (Invitrogen) and Zeocin as selection reagent. Positive clones were screened by measuring sTREM-2 from the supernatant and the best TREM-2-expressing clones were expanded and used for TREM-2 stimulation experiments.
HEK293 cell treatment with TREM-2 agonistic tool antibody and Western blot analysis
Cells were kept at a density of 80% confluency in growth medium (Iscove's Modified Dulbecco's Medium containing 10% fetal calf serum (FCS), 100 μg/mL Zeocin and 100 U/mL penicillin–streptomycin). Cells were treated with 5 μg/mL TREM-2 agonistic antibody for 1 h, subsequently washed and resuspended in RIPA buffer (Radioimmunoprecipitation assay buffer). Proteins were separated using NuPAGE Bis-Tris Gel (Life Technologies, 200 V/100 mA) and transferred onto nitrocellulose membrane. Non-specific binding was blocked by incubation in 5% non-fat dry milk diluted in TBST (0.01% Tween 20 in TBS) for 1 h at room temperature. Membranes were then incubated with anti-phospho-Zap-70 antibody (phospho-Syk Tyr352; Cell Signaling; 1:500 dilution) or Syk antibody (1:1000; Cell Signaling) overnight at 4°C. After washing steps, membranes were incubated with a 1:10,000 dilution of horseradish peroxidase-conjugated anti-rabbit antibodies for 1 h. Bands were developed with the ECL system (Amersham Biosciences) according to the manufacturer's protocols and analyzed using ImageJ software.
OCs stimulation with TREM-2 agonist
Peripheral blood-derived CD14+ monocytes were isolated, as reported above, and cultured as monolayer at the density of 0.5 × 106 cells/mL. In parallel, CD14+ monocytes were seeded on the osteogenic construct following the protocol reported before. Cells were cultured in osteoclastogenic medium for 6 days. Both 2D control and 3D coculture systems were then stimulated over night with TREM-2 tool antibody using the concentration of 5 μg/mL. Supernatants were collected and assessed for CICP, NTX-I, TRAP5b, sTREM-2, and phosphate levels in the 3D experiment. Supernatants from the 2D experiment were assessed for TRAP5b and cells were additionally stained for TRAP activity and analyzed with an Olympus BX-63 microscope.
Statistical analyses
The experiments have been performed three times using three different human donors, unless reported elsewhere. All values are presented as mean ± standard deviation. Differences between experimental groups were assessed with one-way ANOVA tests. p < 0.05 was considered to indicate statistically significant differences.
Results
Establishment of the 3D coculture system
The 3D coculture system was generated as illustrated in Figure 1. hBMCs were seeded and cultured in osteogenic medium for 3 weeks to obtain an osteogenic construct (Fig. 1A). In a second phase, the osteogenic construct was seeded with CD14+ cells from peripheral blood and cells were cultured in osteoclastogenic medium for 3 weeks (Fig. 1B).
During monoculture, hBMCs deposited a dense ECM rich in collagen (Fig. 2A, E and Supplementary Fig. S1A, B; Supplementary Data are available online at
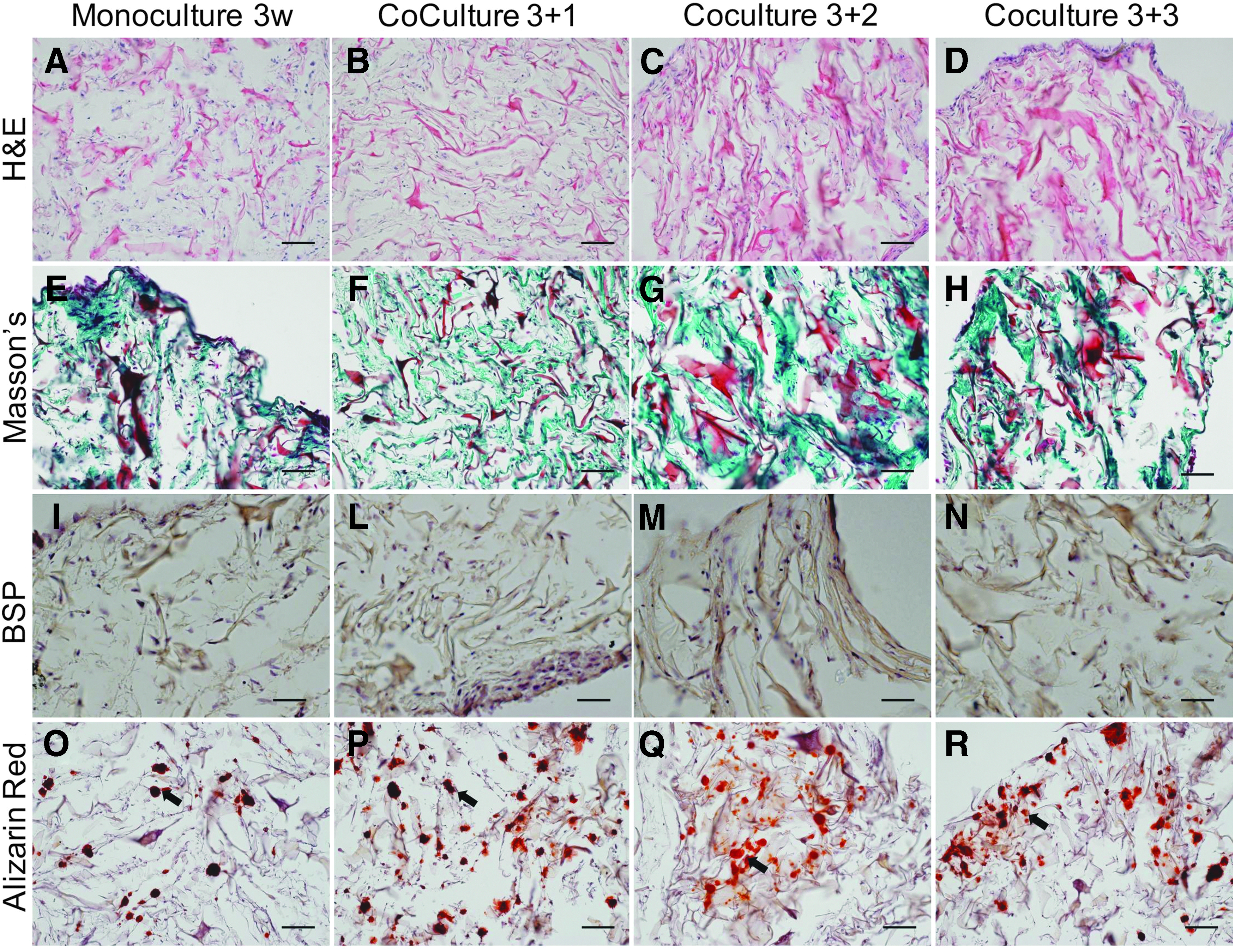
Histological analysis of the 3D system after 3 weeks of monoculture and 3 weeks of coculture. Analysis of the ECM deposition through H&E
Evaluation of OC differentiation and TREM-2 monitoring
As assessed by TRAP staining, CD14+ monocytes differentiated into the OC lineage in the 2D control (Supplementary Fig. S2A–C) and during coculture and were present mainly at the border of the construct (Fig. 3A–F). TREM-2 positive cells were detected by cytofluorimetry at weeks 1, 2, and 3 of coculture (Fig. 3G–I). Overall, these results supported the stability of the coculture system providing a tool for the detection of TREM-2 positive cells and distribution of TRAP positive OCs.

Analysis of OCs differentiation and TREM-2 levels. TRAP staining for OCs activity over time at two different magnifications
Assessment of ECM turnover using non-invasive tools
To non-invasively monitor the process of ECM deposition and resorption over time, typical markers of bone remodeling were assessed in the culture supernatant. Constant levels of CICP were measured at different time points, indicating sustained deposition of Col-I by mesenchymal/osteoblastic cells (Fig. 4A). The levels of NTx-I, increased with time indicating OC-driven degradation of Col-I (Fig. 4B). In parallel, TRAP5b increased with culture time up to 2 weeks of coculture, indicating an increase in the OCs activity (Fig. 4C). Levels of sTREM-2 followed the same trend as TRAP5b and NTx-I, suggesting a possible role of TREM-2 receptor in osteoclastogenic differentiation and ECM resorption. In addition, also cells cultured in 2D showed a sTREM-2 profile similar to the one of TRAP5b (Supplementary Fig. S2 D, E). A reduction in the levels of NTx-I, TRAP5b, and sTREM-2 was observed starting from week 2 of coculture (Fig. 4B–D).
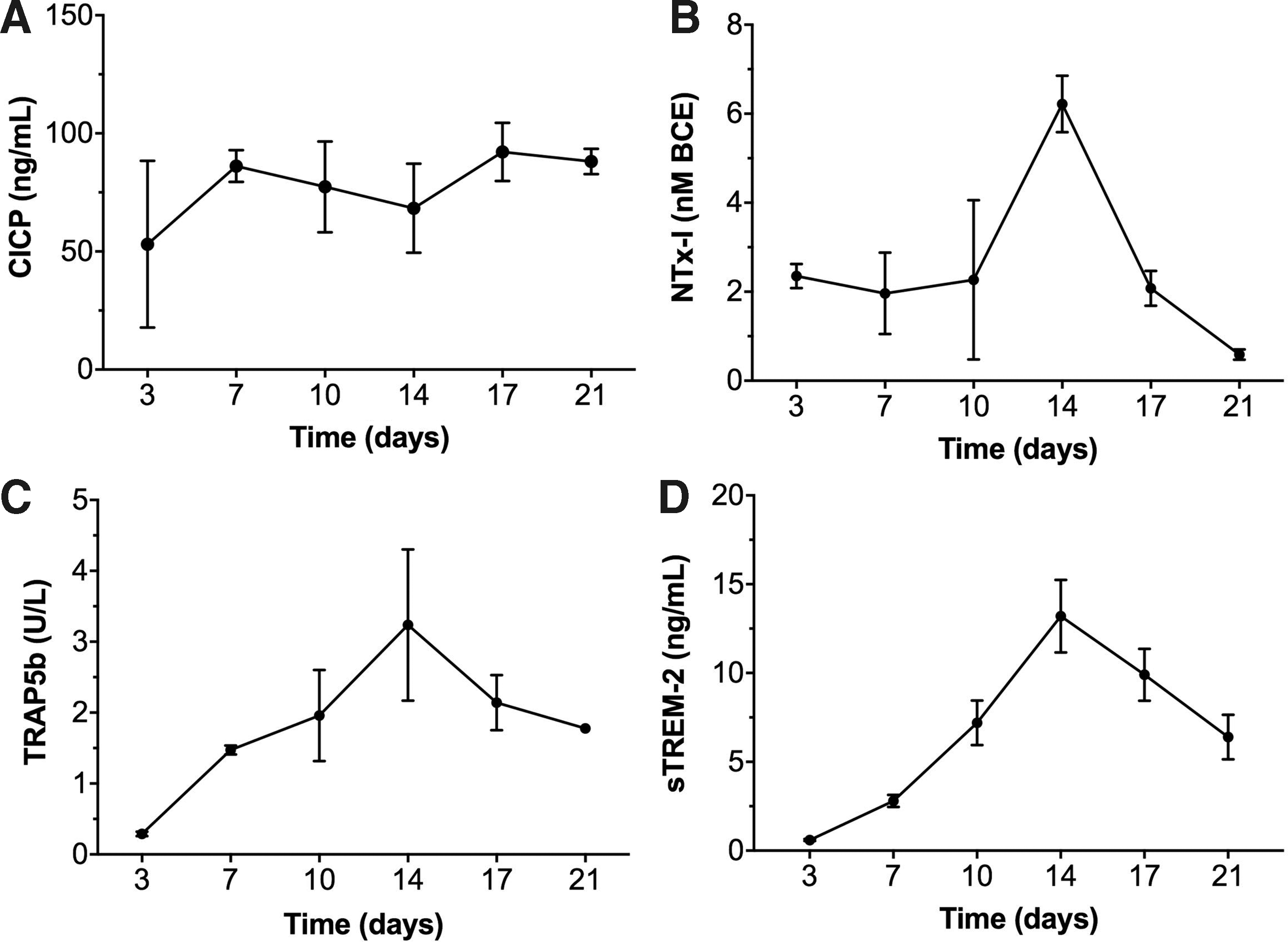
ECM turnover in the coculture system. Supernatant analysis for CICP
Effect of TREM-2 stimulation on OCs
The activation of the TREM-2 pathway in OCs by a TREM-2 agonistic tool was validated in HEK293 cells by using western blot analysis of Syk-phosphorylation. These cells do not spontaneously express TREM-2, therefore wild-type HEK293 cells were used as negative controls. For the investigation of TREM-2 agonistic antibody effects, HEK293 cells were stably transfected with human TREM-2 and its signaling adapter protein DNAX activation protein 12 (DAP12). TREM-2 activation by the tool antibody resulted in increased pSyk levels in both investigated TREM-2 expressing HEK cell clones, while wild-type cells did not show detectable pSyk signal (Fig. 5).
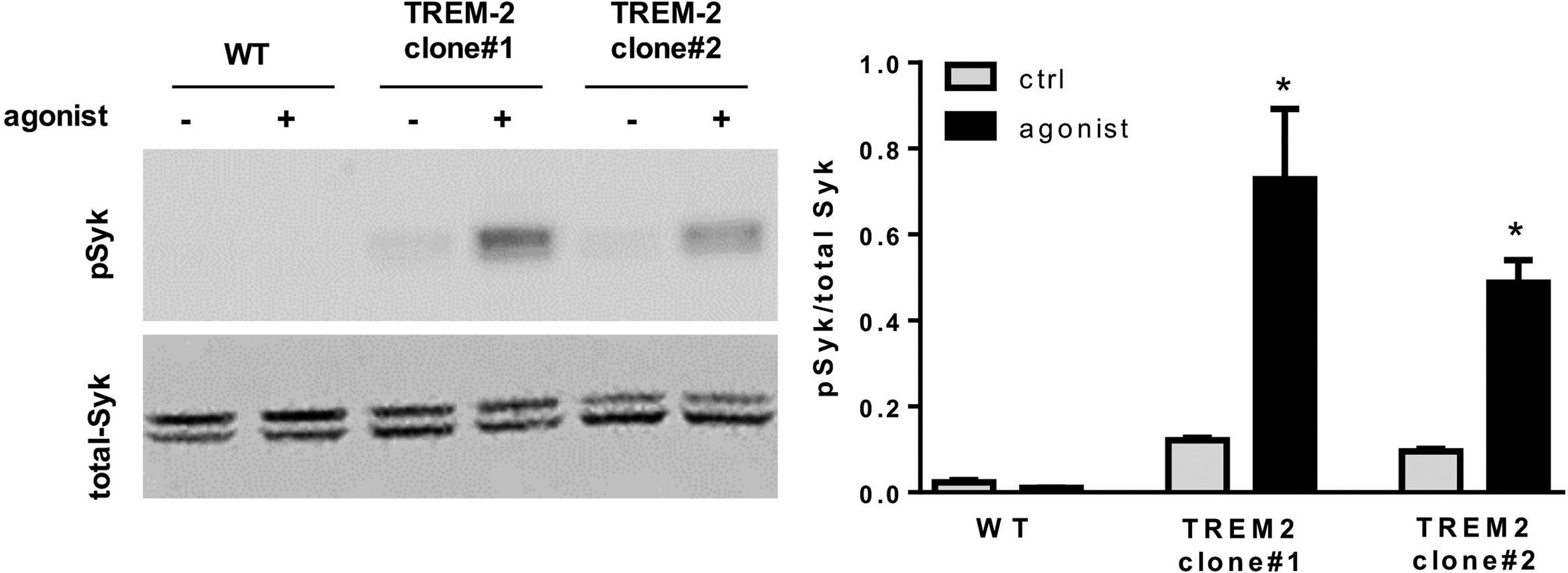
Syk-phosphorylation induced by TREM-2-agonist. Western-blot results revealed Syk-phosphorylation after stimulation with TREM-2 agonistic tool antibody only in HEK cells transfected with TREM-2 and DAP12. *p < 0.05.
Once the 3D culture model, the noninvasive tools for bone homeostasis analysis and the agonistic effect of the TREM-2 antibody were established, we investigated the TREM-2 agonist effect on ECM deposition and resorption together with OCs activity in 2D and 3D. Based on ELISA results for TRAP5b and sTREM-2 obtained in Figure 4, OCs were stimulated at day 6 of differentiation for a duration of 12 h, and OCs activity was subsequently analyzed in term of TRAP5b and ECM turnover at day 7. The agonist had no effect on cells morphology (Fig. 6A, B) or on TRAP5b levels (Fig. 6C) in monolayer cultures of OCs. In the 3D model, no relevant effect of the agonist on the interplay between OCs and osteoblastic cells was observed, since collagen deposition and resorption was similar to the control unstimulated condition (Fig. 6D, E). We also investigated the possible degradation of inorganic matrix components by phosphate level analysis. Phosphate levels in supernatants and TRAP5b activity over time (Fig. 6F, G) showed no difference between the agonist and the control unstimulated condition. In conclusion, TREM-2 agonistic stimulation had no effect on OCs activity in ECM turnover in the 3D coculture model.

Two-dimensional control for monocytes cultured in osteoclastogenic medium for 7 days. TRAP staining for cells stimulated with the agonist
Discussion
In this work, we established a 3D coculture model starting from human progenitors differentiated into OCs and OBs. The goal was to create an in vitro model for the investigation of the role of TREM-2 in bone homeostasis to be used as a possible platform for drug testing. This model efficiently recapitulated the processes of bone matrix deposition and resorption, essential to study bone homeostasis, and has been used to test the effect of receptor modulation on peripheral myeloid cells.
Current models for the investigation of TREM-2 role on bone homeostasis are relying on in vitro 2D cell monolayer cultures of OCs and in vivo knockout mouse models.3,4,6,10 In vitro 2D cultures fail to recapitulate the ECM turnover occurring in vivo, while in vivo models are usually not suitable for the investigation of specific ligands and pathways.3,4,18,23 Human 3D organotypic models already proved to be effective in recapitulating the physiological and pathological state of bone. 21 In a previous work of our laboratory, we established a complex 3D coculture model starting from a heterogeneous set of populations, including endothelial, mesenchymal, and myeloid cells from the stromal vascular fraction of the adipose tissue. 19 Here, we simplified this model by using human mesenchymal cells from bone marrow differentiated in OBs and monocytes form peripheral blood differentiated in OCs. This optimization helped us to build a sensitive platform in which changes in the OBs and OCs homeostasis can be detected using noninvasive tools. This 3D bone-like model therefore provides a relevant system to investigate the role of TREM-2 in bone homeostasis, offering a platform for drug testing, possibly in a personalized setting (using donor-specific cells).
TREM-2 is composed of a single extracellular immunoglobulin-like domain, one transmembrane domain, and a short cytoplasmic tail. 5 The soluble form of TREM-2 can originate by shedding of the receptor extracellular portion or by alternative splicing of TREM-2 transcript leading to secretion.17,24 A recent study investigating the effect of a chimeric TREM-2 protein on mouse OCs reported interference in osteoclastogenesis. 16 Although the role of sTREM-2 is still unknown, a possible mechanism of action has been proposed, in which sTREM-2 would act to counter-regulate the receptor activity by binding the endogenous ligand.16,17 This hypothesis would explain also the sTREM-2 profile that we observed, with an initial progressive increase in response to OCs activity and a subsequent reduction. While the agonistic stimulation was efficient in activating TREM-2 pathway, inducing the syk-phosphorilation in TREM-2 expressing HEK cell, it did not affect OCs activity in 3D coculture system. As released sTREM-2 increased only after OCs differentiation, we presume that the soluble form might also be responsible for the absence of effect that we observed after the stimulation with the agonist. Further studies are needed to elucidate the possible mechanism underlying the role of sTREM-2 in OCs activity and ECM turnover, but our model provides a tool for the monitoring of TREM-2 in its transmembrane and shed form.
TREM-2 has emerged as a possible promising pathway to explore for diagnosis and treatment of AD. 5 We propose that this in vitro 3D model would provide a useful tool for the investigation of the endogenous ligands and the role of the soluble form taking into account the complex interplay between OCs and OBs. Moreover, this system could also be used to investigate the effect of other AD targets modulation on peripheral myeloid cells such as monocytes/OCs.
Conclusion
Before TREM-2 can be considered as a potential target for AD treatment, its role in bone homeostasis has to be elucidated. In this study, we present a 3D in vitro bone model based on the coculture of OBs and OCs in an engineered bone-like environment. Our model recapitulates the dynamic interplay of bone components, providing a tool to study the effect of the TREM-2 pathway on bone homeostasis. The model suggests absence of side effects on bone homeostasis of drugs modulating the activity of this receptor.
Footnotes
Acknowledgments
This work was funded by Roche Pharmaceuticals, Basel, Switzerland. Authors are grateful to Dr. Fabiana Gullotta and Dr. Atanas Torodov for the scientific advice and technical support.
Authors' Contribution
E.R., A.P., and A.S. designed the study with the help of I.K. and I.M.; A.M. supplied biological samples; E.R., N.A., and D.R. performed the experiments; E.R. analyzed the data; E.R., A.S., I.K., and E.M. interpreted the results; E.R. and A.S. wrote the article. All the authors discussed the results, commented, and revised the article.
Disclosure Statement
None of the authors has any conflicts of interest to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.