
Abstract
Bioactive scaffolds are used in cartilage regeneration for providing an environment conducive to chondrogenic differentiation and proliferation of stem cells in vitro and in vivo. This study aimed to develop a composite scaffold comprising silk fibroin (SF) and cartilage extracellular matrix (ECM), which could not only preserve the biological activity of the original ECM, which promotes chondrogenic differentiation of bone marrow-derived mesenchymal stem cells (BMSCs), but also could provide great mechanical strength. To this end, we manufactured scaffolds containing different ratios of SF and ECM (1:4, 1:2, and 1:1 SF-ECM) by lyophilization with a novel crosslinking method. The results showed improved mechanical strength with increasing SF concentrations. Compared to the ECM control, the SF-ECM scaffolds allowed a greater increase in glycosaminoglycan and collagen concentrations in vitro. According to histological findings, the SF-ECM scaffolds exhibited significant biocompatibility and provided an environment for cartilage regeneration after transplantation in vivo. The SF-ECM scaffolds also possessed good biomechanical properties and the ability to promote chondrogenic differentiation of BMSCs. These findings open up new possibilities for evaluating SF-ECM scaffolds as potential candidates for cartilage regeneration.
Impact Statement
This study presented a new method to fabricate SF-ECM scaffolds that potentially promote chondrogenesis of BMSCs, and open up new possibilities for using SF-ECM scaffolds as an off-the-shelf strategy for joint cartilage regeneration. It is worthy of further investigation in knee joints of animals, and beyond knee cartilage, this scaffold may also serve as an ideal biomaterial for the regeneration of other joint cartilages.
Introduction
A
Extracellular matrix (ECM), a fast emerging scaffold biomaterial derived from native tissues, has been used to promote cell differentiation and proliferation and matrix formation in many tissue-engineering applications.12–21 Accordingly, a promising bioscaffold has been proposed based on ECM derived from decellularized cartilage tissue (primarily consisting of proteoglycans and type-II collagen) in which growth factors are incorporated for providing cues to enhance cartilage regeneration.22–26 Devitalization and decellularization of cartilage ECM produce scaffolds which preserve the bioactivity of materials for application in cartilage tissue engineering.27–30 Scaffolds of this type have been shown to be chondroinductive in previous studies.24–26 Nevertheless, owing to the inadequate strength of ECM-derived scaffolds, the present study additionally used silk fibroin (SF) as one of the components of the structural matrix to meet the requirements for cartilage-tissue engineering.
SF, derived from degummed raw silk, is widely used in tissue engineering as a biomaterial owing to its remarkable mechanical strength, biodegradable nature, biological compatibility, and abundant sources. Previous studies have demonstrated that SF, as a degradable protein-based scaffolding biomaterial, could be a preferential choice for constructing bone marrow-derived mesenchymal stem cell (BMSC)-based scaffolds for cartilage regeneration.31,32
Contraction of constructs is an impediment to the success of chondrogenesis of BMSCs during in vitro culture and implantation into a defect.33–35 In an attempt to overcome this problem, different crosslinking techniques were investigated to prevent the contraction of ECM scaffolds in previous studies. Genipin, a natural crosslinking agent derived from gardenia fruit extract, bridges the amino groups of hydroxylysine and lysine residues in silk protein and ECM.36,37 In several studies, genipin has proved less toxic than other crosslinkers such as glutaraldehyde and 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride, which have been widely used for crosslinking various biomaterials, including SF and ECM.33,38 Methanol and ethanol can induce the formation of β-sheet structures in SF.38,39 The properties of SF, such as its mechanical strength and biodegradable nature, can be controlled by regulating the content of β-sheets.38,40,41 In the present study, both genipin and ethanol were used for SF-ECM scaffold fabrication.
The main aim of our study was to develop a SF-ECM scaffold that combined the capability of promoting robust chondrogenic differentiation of BMSCs with appropriate mechanical properties. To this end, we first fabricated scaffolds containing different ratios of SF-ECM and compared their structural and mechanical properties. Next, we sought to determine whether altering the ratio of SF and ECM would have an effect on the cytotoxic and chondroinductive properties of the constructs in vitro. Finally, we compared the efficacy of chondrogenesis of BMSCs among different scaffolds in vivo.
Materials and Methods
All experimental procedures involving animals were approved and conducted in compliance with the guidelines on animal use outlined by the Nanjing Medical University School of Medicine. All animal subjects received care in accordance with the “Guide for Care of Laboratory Animals,” as detailed by the National Institutes of Health.
Preparation of ECM suspension
Cartilage segments were harvested from the costal cartilage of female goats (n = 12) within 6 h of sacrifice and transported to the laboratory in sterile conditions. The costal cartilage was first cut into small pieces (1 mm 3 ) using eye scissors. To manufacture a cartilage slurry, small pieces of cartilage were blended in phosphate-buffered saline (PBS) using a homogenizer (IKAT10; IKA). The homogenized cartilage tissue was centrifuged, and the supernatant was removed. The remaining material was freeze-dried overnight and pulverized by milling (JXFSTPRP-24; Shanghai Jingxin) to produce powder-like cartilage, which was then subjected to a three-step decellularization procedure, as described by Courtman et al. 42 To inhibit protease activity, the powder-like cartilage was first blended in PBS containing 0.1% (w/v) ethylenediaminetetraacetic acid (EDTA; Sigma-Aldrich) and 3.5% (w/v) phenylmethyl sulfonylfluoride (Beyotime, Shanghai, China) using a homogenizer (IKAT10; IKA) and stirred constantly for 24 h at 4°C. In the second stage, the mixture was treated with a 1% solution of Triton X-100 in Tris-HCl and gently agitated for protease inhibition for 24 h at 4°C. The samples were thoroughly rinsed in PBS before the third stage, which involved digestion with 50 U/mL deoxyribonuclease and 1 U/mL ribonuclease (Sigma-Aldrich) in 10 mM Tris-HCl (pH 7.5), with agitation for 12 h at 37°C. The decellularized cartilage matrix was then washed with sterile PBS and made up to a 6.9% (w/v) suspension.
Preparation of silk solution
Cocoons (15 g) were boiled for 30 min in a degumming solution comprising 0.02 M Na2CO3 and then washed thoroughly in double-distilled water to remove the sericin protein. After drying, the degummed silk was dissolved in a 9.3 M LiBr solution at 60°C for 4 h. This solution was dialyzed in a 3500 kD dialysis bag against double-distilled water for 3 days to remove LiBr. Then, the solution was centrifuged at 9000 rpm for 20 min at 4°C to remove silk aggregates that had formed during dialysis. The supernatant was freeze-dried overnight and made up to a 6.9% (w/v) silk solution.
Scaffold preparation
We designed different kinds of scaffolds which were cylindrical shaped 6 mm in diameter and 2 mm thick by mixing the 6.9% (w/v) SF solution and ECM suspension at mass ratios of 1:1, 1:2, and 1:4. The mixture was carefully poured into a mold and freeze-dried overnight. The scaffold was immersed in 0.5% genipin-ethanol solution for 3 days to allow β-sheet formation and crosslinking. Then, the scaffolds were washed in double-distilled water for 48 h and dehydrated for use. All scaffolds were sterilized by 60 Coγ irradiation.
Cell culture
BMSCs were isolated from the bone marrow cavities of bilateral femurs of Sprague Dawley rats and identified in accordance with our previously described method. Briefly, the marrow cavity was rinsed with serum-free, low glucose Dulbecco's modified Eagle's medium (L-DMEM). Cells were collected by centrifugation at 1200 rpm for 5 min, suspended in L-DMEM supplemented with 10% fetal bovine serum (FBS; HyClone, Logan, UT), and incubated at 37°C with 5% CO2. After 24 h, unattached cells were removed from the primary culture, and adherent cells were cultured further by changing the culture medium every 3 days. Cells that reached confluence were defined as passage 0. Experiments described in this study used BMSCs at passage 3.30,43
The BMSCs were counted with a hemocytometer. The cells were resuspended in a culture medium at 500,000–1000,000 cells per 50 μL and seeded by pipetting 50 μL directly onto the scaffolds. The cell-seeded constructs were placed into 24-well plates. After incubation at 37°C for 2 h, the cells were attached to the scaffolds, and culture medium (L-DMEM, 10% FBS, and 1% penicillin and streptomycin) was added in each well. The cell-seeded constructs were incubated in vitro at 37°C in 5% CO2, and the medium was changed every 2–3 days.
Water uptake and degradation of scaffolds
Several concentrations of SF-ECM scaffolds were initially lyophilized and weighed (W0). The lyophilized scaffolds were then incubated in PBS at 37°C and weighed (WW) to evaluate their water uptake ability and degradation rate. At each time point, the PBS was removed, and the scaffolds were lyophilized and weighed (Wd). The degradation rate of ECM-silk scaffolds was measured by the percentage of dry weight loss. Water uptake and degradation rate were defined using formulas (a) and (b) (n = 3).
Mechanical properties
To evaluate the compressive modulus of each scaffold, bare SF-ECM scaffolds were mechanically characterized using an Instron Tensile Force Tester (Instron Corporation) and performed in compression. Scaffolds were loaded with a constant compression speed of 10 mm/min to obtain stress–strain data. The linear region of the stress–strain curve was used to calculate Young's modulus. The scaffolds were then immersed in PBS for 12 h, and, likewise, the compression modulus of the rehydrated scaffolds was measured.
Fourier-transform infrared spectroscopy
Infrared spectra of scaffolds were analyzed using the Continuum FT-IR Microscope (Thermo NICOLET 6700). The scaffolds were washed with PBS and lyophilized overnight to prepare them for Fourier-transform infrared (FTIR) analysis. Scanning was performed in the spectral range of 400–4000 cm−1.
Scanning electron microscopy
The surface morphology of scaffolds was analyzed by scanning electron microscopy (SEM). The scaffolds were lyophilized, coated with gold-palladium, and observed for surface morphology using a SEM (S3400II; Hitachi Medical Corporation, Tokyo, Japan)
Laser confocal microscopy
Cell-scaffold constructs were fixed overnight at 4°C in 4% paraformaldehyde. Subsequently, the samples were incubated with phalloidin (Actin-Tracker Green; Beyotime), diluted (1:200) in 1% bovine serum albumin for 1 h at room temperature, and then washed thrice with PBS for DAPI (Beyotime) staining. Images were acquired using a laser confocal microscope (LSM710, Oberkochen, Germany).
Cell proliferation rate and viability
To measure the cell proliferation rate and viability offered by different scaffolds (n = 3), BMSCs seeded on different scaffolds were evaluated using the Cell Counting Kit-8 (CCK-8; Beyotime, Nanjing, China) in accordance with the manufacturer's protocol. Cells cultured for 1, 3, 5, and 7 days were treated with 100 μL CCK-8 solution and incubated for 1 h at 37°C. The culture medium was then transferred into 96-well plates and measured for optical density at 450 nm using a microplate reader (Thermo Fisher, Massachusetts).
For assessment of cell viability, BMSCs seeded on scaffolds were stained using the LIVE/DEAD assay (Molecular Probes, Eugene, OR) after 72 h of culture and observed under a fluorescence microscope (Zeiss, Nanjing, China). Briefly, the constructs were incubated in 2 × 10−6 M calcein and 4 × 10−6 M ethidium homodimer-1 for 45 min at room temperature. Calcein (green/live) and ethidium homodimer-1 (red/dead) were detected at excitation wavelengths of 495 and 560 nm.
Biochemical analysis
For biochemical analysis, BMSCs were seeded on different scaffolds and analyzed for sulfated-glycosaminoglycan (sGAG) and collagen content on days 0, 14, and 28. The constructs were dissolved by papain digestion (125 μL/mL papain, 100 mM EDTA, and 5 mM
mRNA expression
For gene-expression analysis, BMSCs were seeded on different scaffolds (n = 3) and cultured for 28 days. For mRNA extraction, the cells were treated with TRIzol reagent (Invitrogen) and centrifuged at 12,000 rpm at 4°C. The mRNA levels of SOX-9, type-2 collagen (COL II), type-1 collagen (COL I), and type-10 collagen (COL X) were determined by real-time quantitative polymerase chain reaction (qPCR, StepOne Real-Time PCR System; Applied Biosystems) using the SYBR Green PCR Master Mix (Toyobo, Osaka, Japan) as previously described.45,46 The PCR primer sequences used in this study are shown in Table 1. Target mRNA was quantified by normalizing their expression levels to that of a housekeeping control (glyceraldehyde 3-phosphate dehydrogenase mRNA) using the ΔΔCt method. 47
Primer Sequences
Immunohistochemical analysis: in vivo test
To evaluate the ability of SF-ECM scaffolds to promote cartilage regeneration, scaffolds loaded with BMSCs were implanted into 6-week-old nude mice (n = 24) in the subcutaneous region. The mice were anesthetized by intramuscular injection of pentobarbital sodium (40 mg/kg) and then sacrificed for harvesting the scaffolds on days 14 and 28 postsurgery. The harvested scaffolds were fixed overnight in 4% paraformaldehyde at 4°C, dehydrated using a graded series of ethanol, embedded in paraffin, and sectioned at a thickness of 10 μm. After deparaffinization, the samples were stained with hematoxylin and eosin (H&E), safranin O, and Masson's trichrome stain. Stained sections were observed and imaged with an Olympus microscope. We randomly selected six different areas in each sample. Images were thresholded and then analyzed for pixel density to quantify proteoglycan and collagen content using Image analysis software (ImageJ). Data were expressed as the percentage of the total amount of proteoglycan (red) and collagen (blue) in the region. All images were taken at the same exposure time and conditions.
To further identify the expression of chondrogenic proteins (COL I and II), immunohistochemical staining was performed as previously described. 48
Statistical analysis
Data are expressed as mean value ± standard deviation (X ± SD). Statistical analysis was performed using IBM SPSS Statistics 19.0. Intergroup differences were analyzed by one-way analysis of variance after testing for the homogeneity of variance and the data between any two groups by the Q test. Statistical significance was determined by analysis of variance with p < 0.05.
Results
Physical characterization of scaffolds
FTIR spectroscopy
For analysis of scaffold components, the crystallization of SF was analyzed by FTIR spectroscopy (Fig. 1a). Noncrosslinked SF/ECM mixtures showed two main peaks at 1649 cm−1 (amide I) and 1543 cm−1 (amide II), demonstrating that the mixture was mainly composed of random coils.38,49 After crosslinking by ethanol, the major peak shifted to lower wavelengths at 1620–1622 and 1526–1528 cm−1 in the amide-I (C = O stretch) and amide-II (N-H bend) regions, respectively, assigned to the band of aromatic and aliphatic carbonyl stretch. This indicated that the crosslinked SF-ECM scaffolds comprised a greater proportion of β-sheets than random coils.

Surface characteristics of the scaffolds
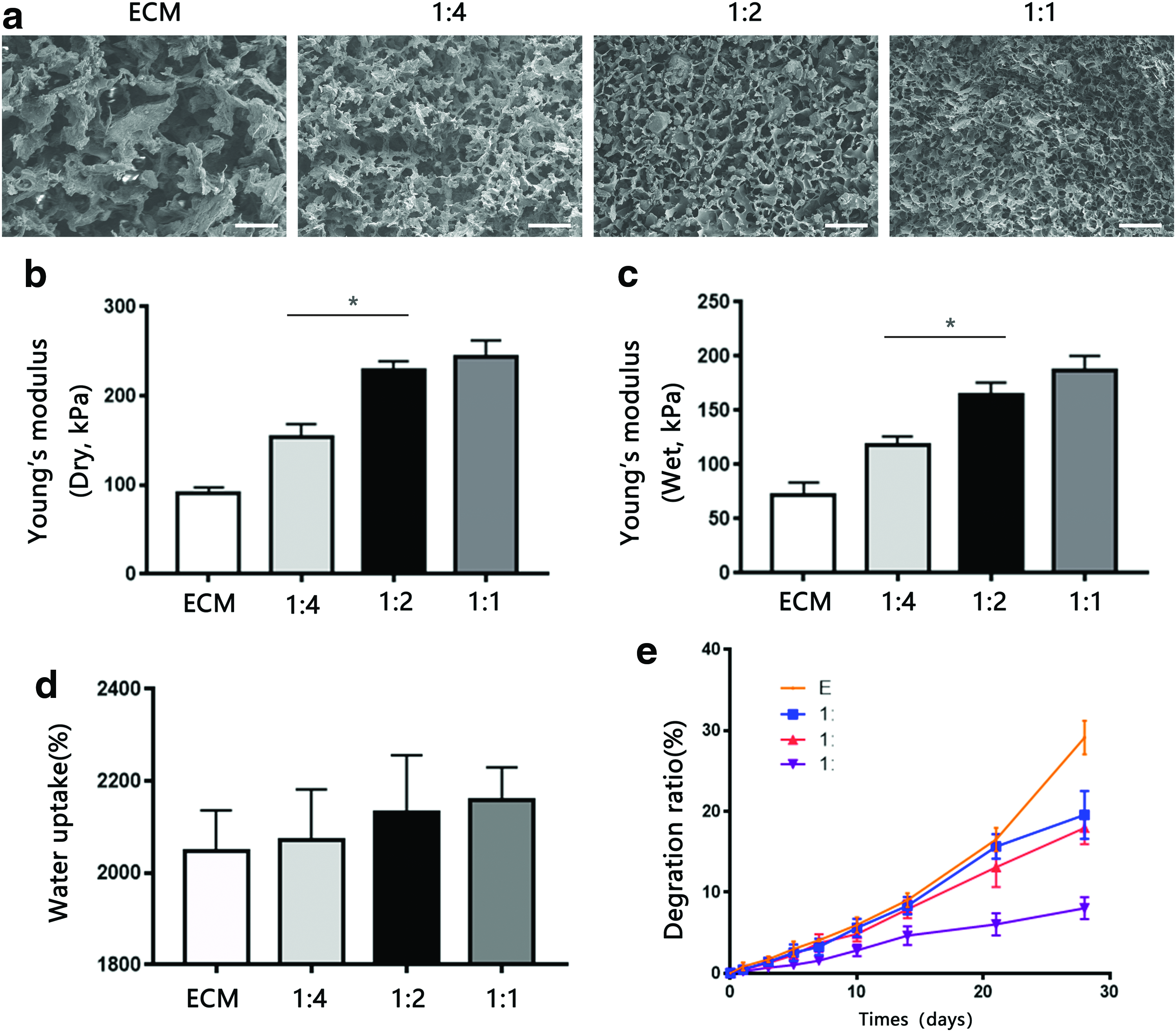
The surface characteristics of each scaffold in cross-section were observed by SEM (Fig. 2a). All scaffolds exhibited a porous structure with different pore sizes. The mean pore sizes of the scaffolds decreased with an increase in SF content (Table 2). The pore sizes of the ECM and 1:4, 1:2, and 1:1 SF-ECM scaffolds were 179.96 ± 12.62, 112.87 ± 12.12, 87.29 ± 14.21, and 55.79 ± 9.45 μm, respectively. The SF-ECM scaffolds exhibited a smoother surface than did the ECM scaffold. Moreover, scaffolds with higher concentrations of SF exhibited fewer uncovered ECM particles.
Pore Size and Porosity of Extracellular Matrix and Silk Fibroin–Extracellular Matrix Scaffolds
Data presented as mean ± standard deviation, n = 3.
ECM, extracellular matrix; SF, silk fibroin.
Compressive strength
Figure 2b and c shows the results of analysis of the compressive strength of scaffolds without cells. The compressive moduli of dry scaffolds without cells increased with increase in SF concentration; however, upon hydration, the scaffolds became noticeably softer. The Young's moduli of dry scaffolds increased in the following order: ECM (0.08995 ± 0.07229 MPa) <1:4 SF-ECM (0.1538 ± 0.01438 MPa) <1:2 SF-ECM (0.2290 ± 0.01001 MPa) <1:1 SF-ECM (0.2432 ± 0.01858 MPa). The Young's moduli of hydrated scaffolds increased in the following order: ECM (0.07219 ± 0.06429 MPa) <1:4 SF-ECM (0.1180 ± 0.04633 MPa) <1:2 SF-ECM (0.1638 ± 0.01161 MPa) <1:1 SF-ECM (0.1871 ± 0.01323 MPa).
Water uptake and degradation of scaffolds
Figure 2d shows the water absorption capacities of the scaffolds. Relative to the pure ECM scaffold (2046% ± 89.34%), ECM scaffolds containing SF exhibited slightly increased absorption, with the water uptake percentage increasing with increase in SF concentration (1:4, 1:2, and 1:1 SF-ECM: 2070% ± 112%, 2132% ± 124%, and 2142% ± 125.4%, respectively). Figure 2e shows the weight loss ratios of scaffolds at several time points over 28 days. The degradation rates of ECM scaffolds decreased with an increase in SF concentration.
In vitro biological properties
Viability of BMSCs on SF-ECM scaffolds
The viability of cells grown on SF-ECM scaffolds was quantitatively determined by the CCK-8 assay on days 1, 3, 5, and 7 of incubation (Fig. 3b). Relative to the ECM scaffold, scaffolds containing SF showed lesser cell growth on days 1–3. However, by day 5, the cell proliferation rate in SF-ECM scaffolds increased and exhibited no significant difference with that in the ECM scaffold. The viability of BMSCs seeded on the scaffolds was evaluated by fluorescence microscopy after staining with the LIVE/DEAD assay (Fig. 3a). BMSCs were evenly distributed on the SF-ECM scaffolds. The results demonstrated that the SF-ECM scaffolds exhibited low cytotoxicity and were suitable for BMSC growth.
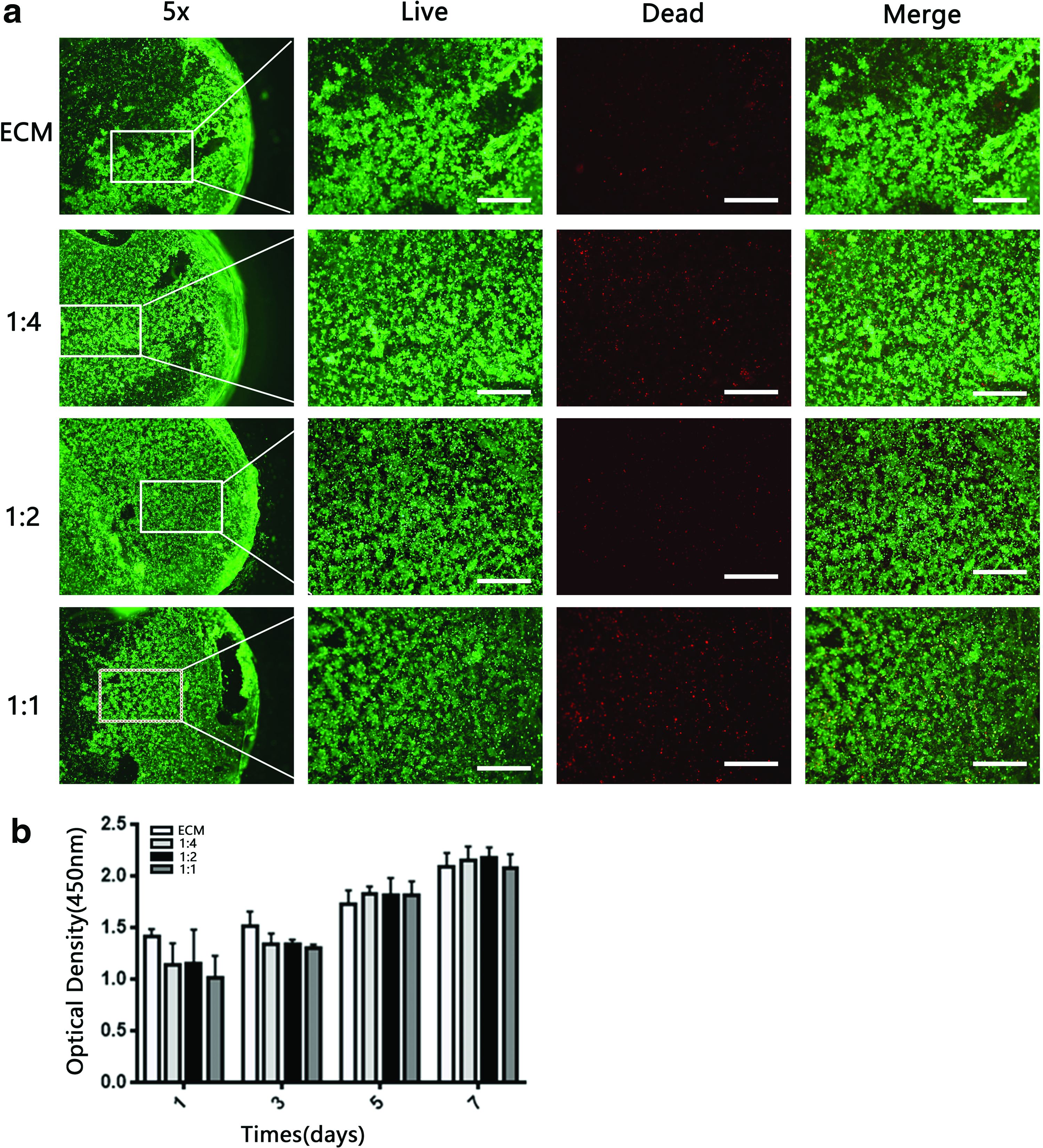
Viability of BMSCs seeded on SF-ECM scaffolds.
BMSC morphology on scaffolds
After incubation with different groups of scaffolds for 7 days, the morphology of BMSCs was observed by confocal microscopy (Fig. 4a). In the ECM scaffold, the BMSCs exhibited a typical fusiform morphology. Compared to BMSCs grown on ECM or other groups of scaffolds, those grown on the 1:2 SF-ECM scaffold showed a more round shape, which indicated a chondrogenic morphology.
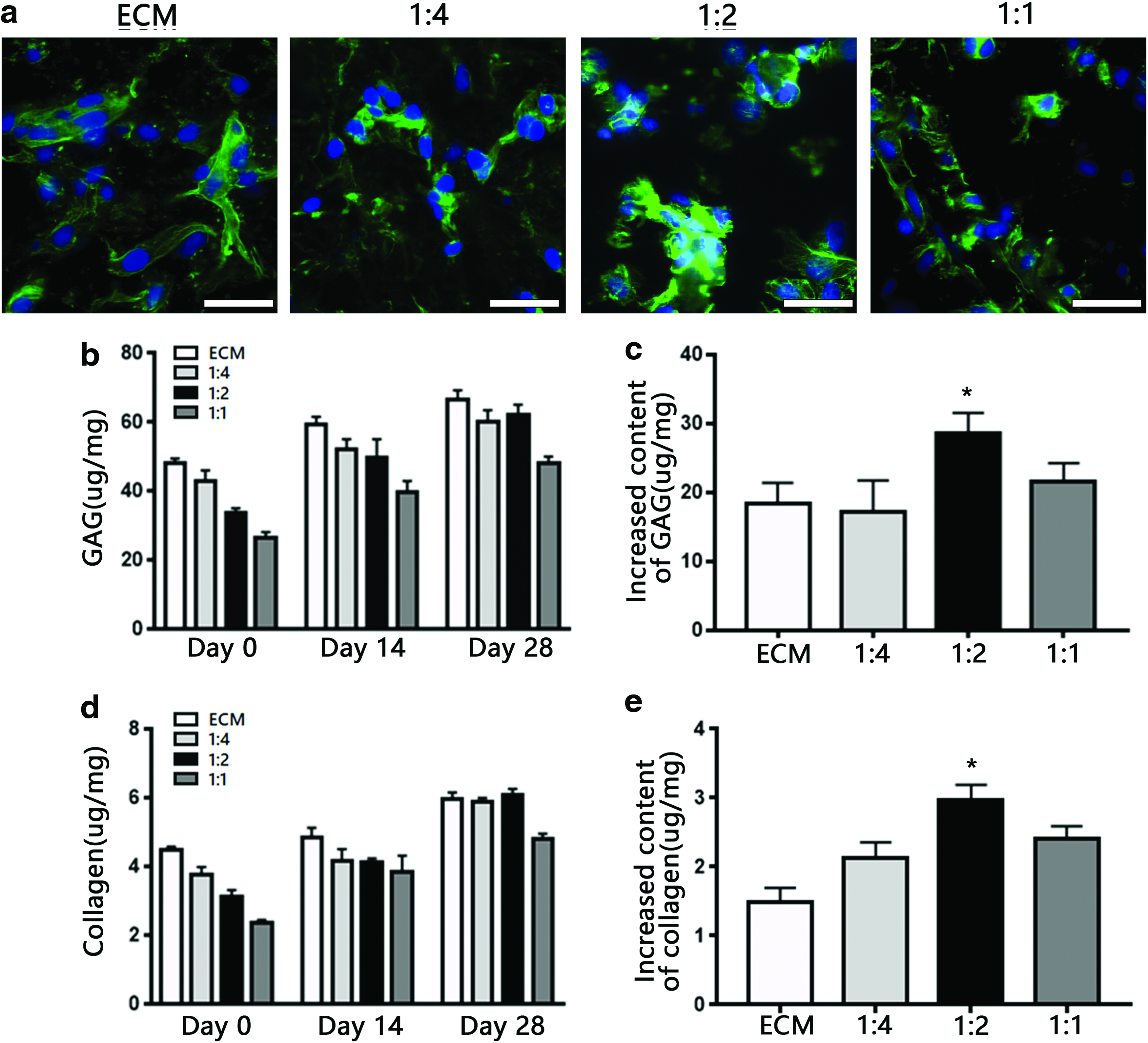
sGAG and collagen content
The total collagen and sGAG contents of the constructs were quantitatively analyzed at several time points (Fig. 4b, d). The collagen and sGAG concentrations in the 1:2 SF-ECM scaffold gradually increased and matched those in the pristine ECM scaffold. After 28 days of culture, relative to other constructs, the 1:2 SF-ECM construct exhibited the greatest increase in sGAG and collagen content (Fig. 4c, e).
mRNA-expression profile of BMSCs on SF-ECM scaffolds
We investigated the chondrogenic differentiation of MSCs in scaffolds by quantifying the mRNA-expression levels of chondrogenic-related genes such as SOX-9, COL I, and COL II and hypertrophic-related genes COL X (Fig. 5) by real-time qPCR on day 28 of culture. SOX-9, a member of the SOX (Sry-type HMG box) gene family, is expressed during early mesenchymal condensation and activates COL II expression. 50 Relative to BMSCs grown on other kinds of scaffolds, those grown on the 1:2 SF-ECM scaffold expressed the highest levels of SOX-9. The expression levels of COL II mRNA in BMSCs grown on 1:2 and 1:1 SF-ECM scaffolds were similar and higher than those in cells grown on other groups of scaffolds, although the differences were not apparent. Because COL II expression indicates chondrogenic differentiation in cartilage, and COL I expression indicates chondrocyte dedifferentiation, we used the ratio of COL II to COL I mRNA expression as an index for differentiation of chondrocytes. 51 The COL II:COL I expression ratio in the 1:2 SF-ECM scaffold was approximately twofold higher than that in the ECM scaffold, which indicated predominant chondrogenic differentiation in the former. The expression level of COL I and COL X mRNA in cells grown on the 1:2 SF-ECM scaffold was lower than that in cells grown on the ECM scaffold.
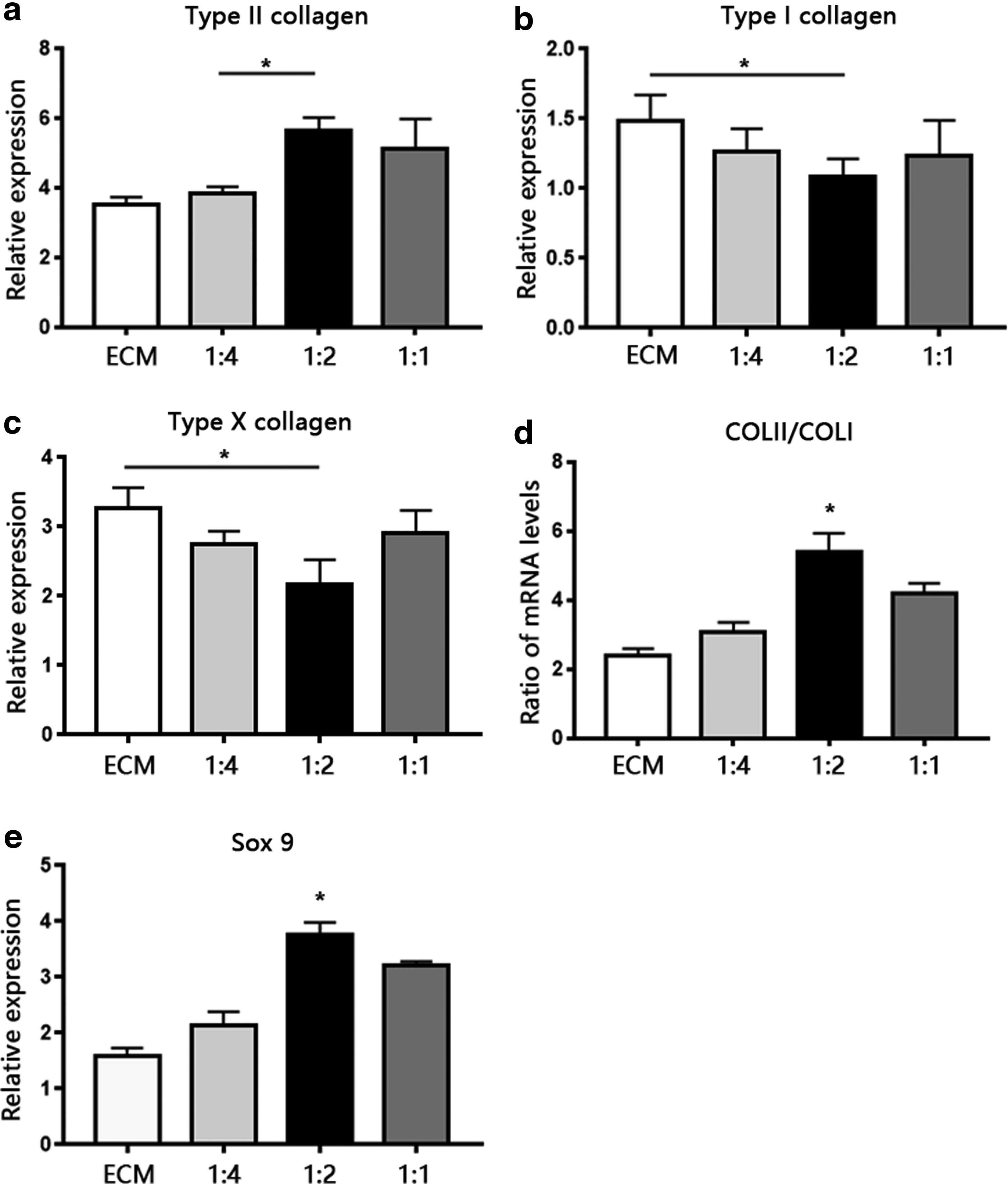
mRNA expression levels of fibrotic-marker gene
Histological evaluation: in vivo test
To investigate the ability to promote chondrogenesis in vivo, scaffolds loaded with BMSCs were transplanted into 6-week-old nude mice in the subcutaneous region. All scaffolds exhibited the capacity to promote chondrogenesis and cartilage regeneration. The results of H&E staining revealed that the BMSCs began to proliferate and secrete ECM, which gradually took over the scaffold starting from day 14 (Fig. 6). The results of safranin O and Masson's trichrome staining indicated the presence of proteoglycan and collagen, respectively, in all cartilaginous tissues (Fig. 7a, b). Positive red and blue staining in all scaffold sections indicated the presence of cartilaginous ECM. The proteoglycan and collagen density were quantified using ImageJ, and they were significantly elevated in 1:2 SF-ECM construct at day 28 (Supplementary Fig. S1; Supplementary Data are available online at
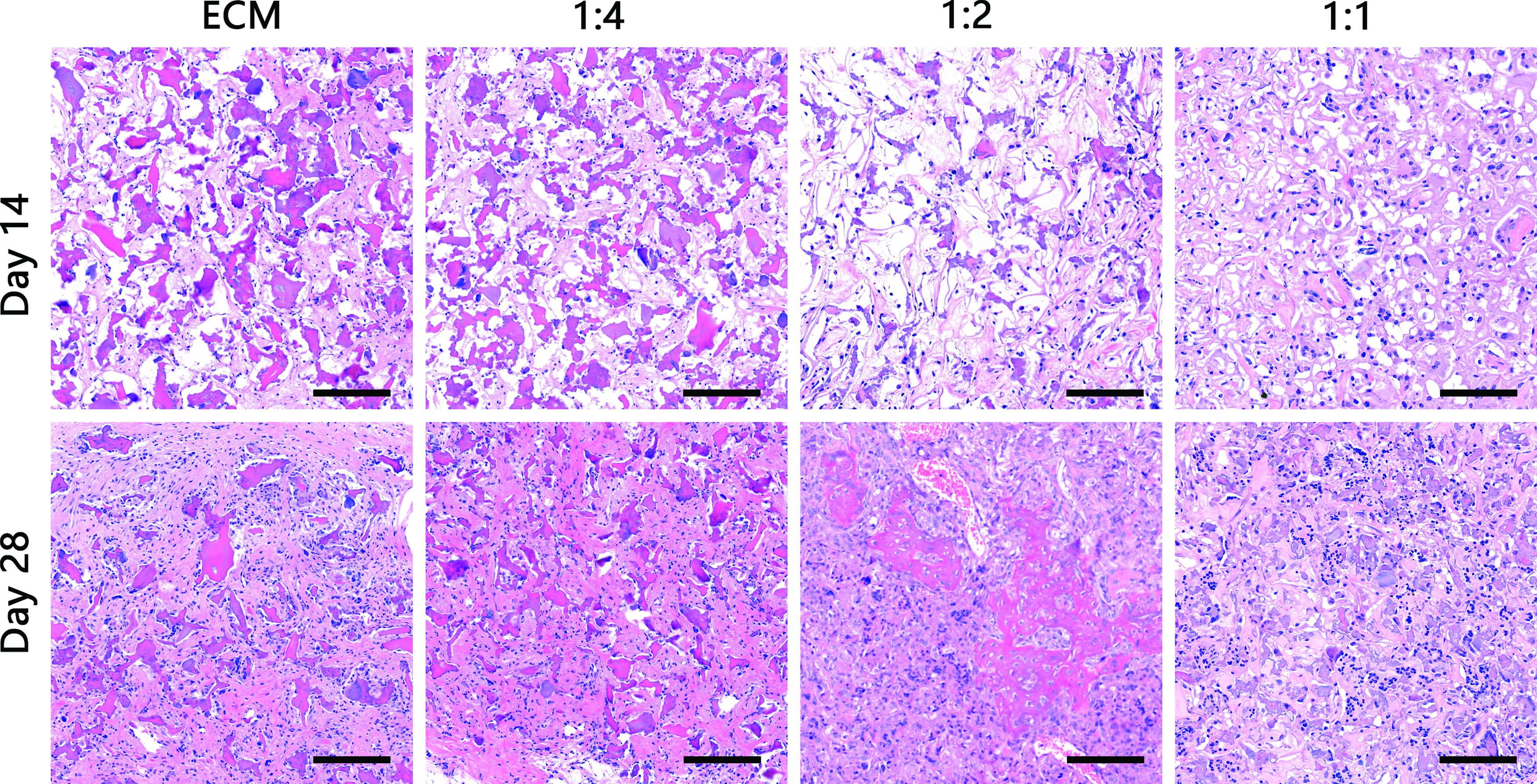
BMSCs were seeded on SF-ECM scaffolds and implanted into nude mice for 14 and 28 days. H&E-stained sections of implanted scaffolds (n = 6; magnification, 20 × ; scale bar = 100 μm). H&E, hematoxylin and eosin. Color images available online at
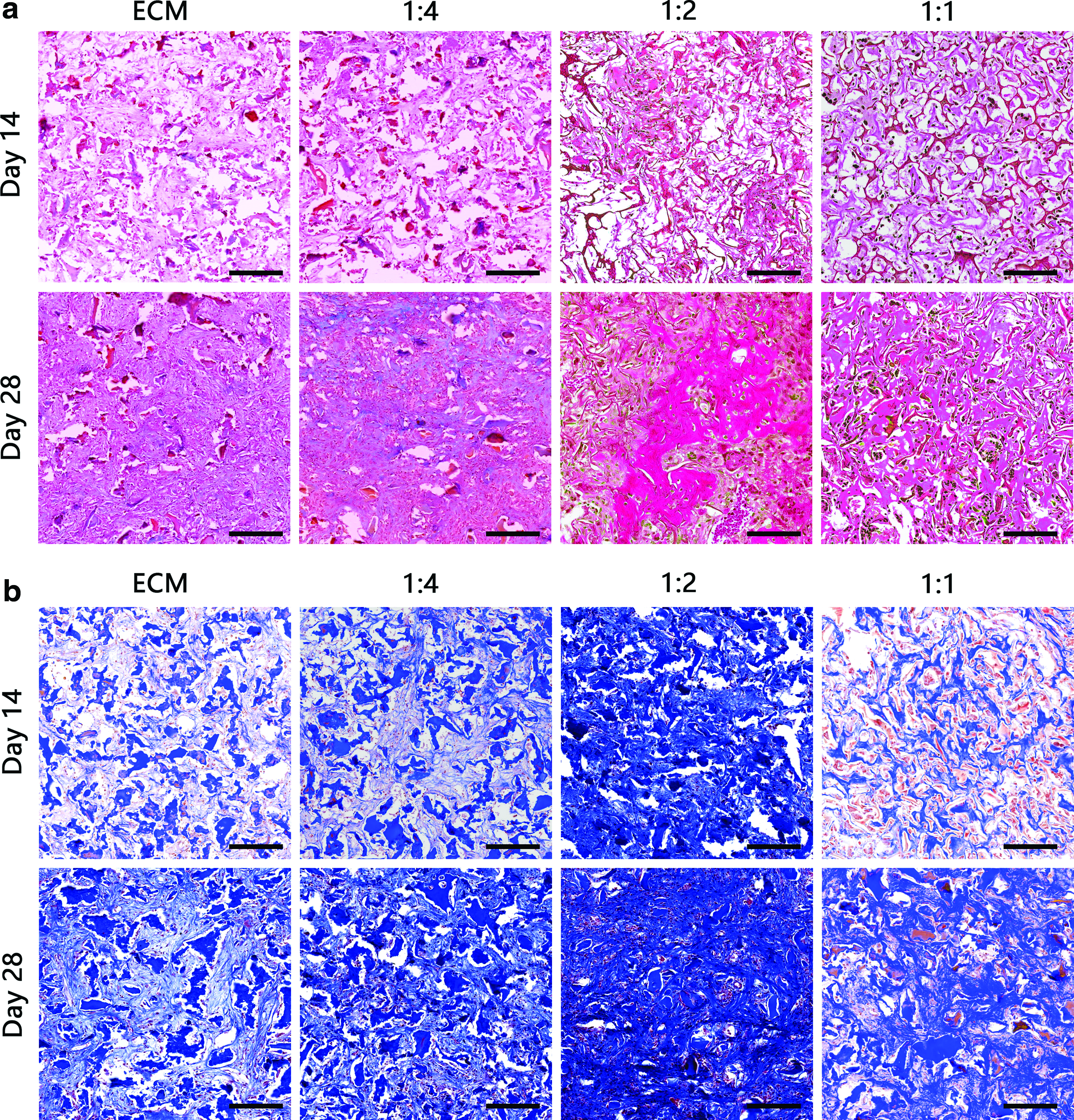
Histological sections of scaffolds implanted for 14 and 28 days in the subcutaneous region of athymic nude mice, stained with
The results of immunochemical staining helped determine the COL I and II content in the scaffolds (Fig. 8a, b). Relative to the staining intensity observed on day 14 in vivo, there was a significant increase in staining intensity in the samples on day 28. The 1:2 SF-ECM scaffold exhibited higher COL II and lower COL I content than other scaffolds.
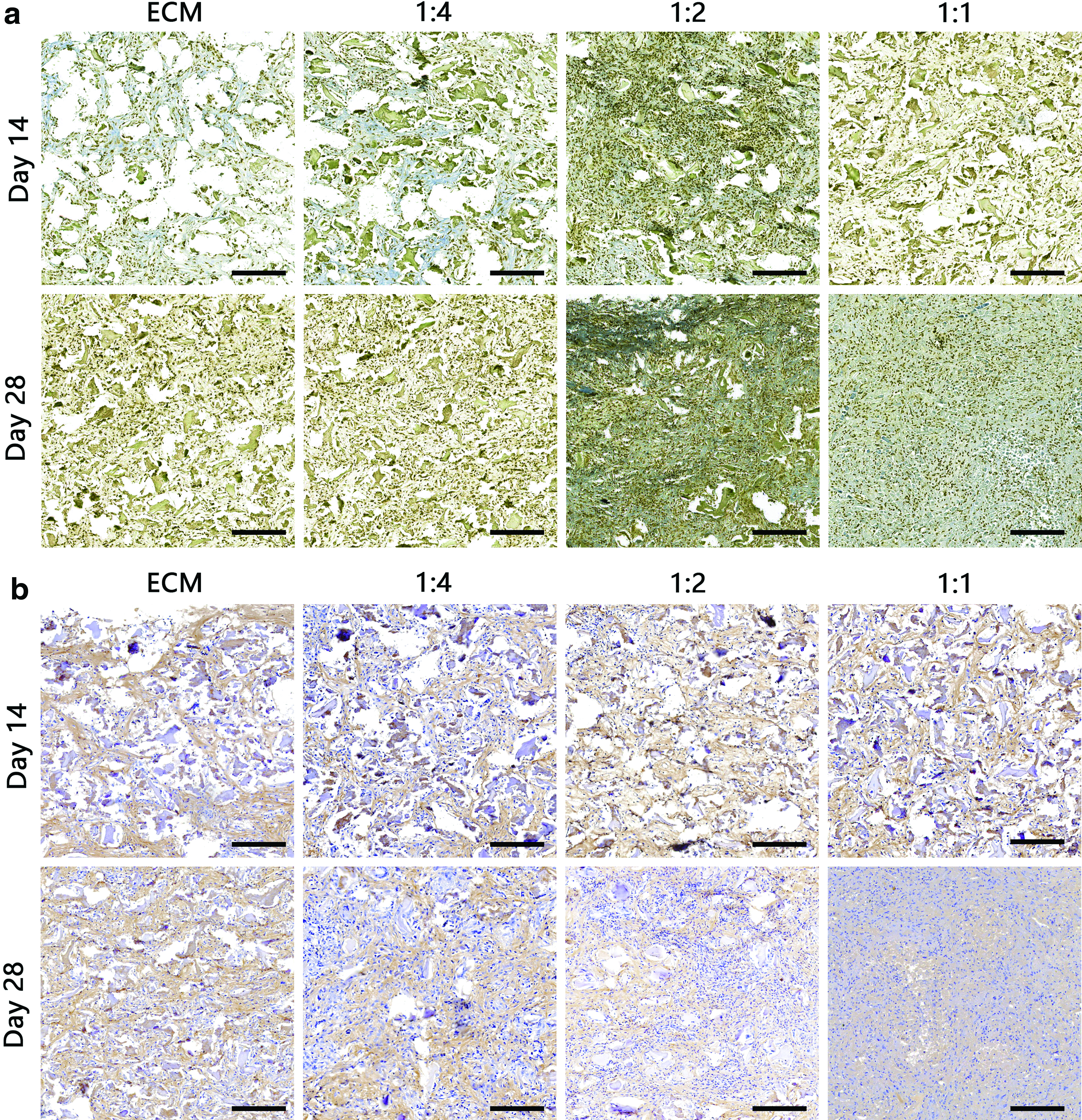
Results of immunohistochemical analysis of collagen
Discussion
The overall objective of this study was to develop a promising single stage or off-the-shelf therapy involving SF-ECM scaffolds with improved mechanical properties. By varying the component ratios and concentrations of ECM and SF, we were able to fabricate scaffolds with better mechanical properties than existing ECM scaffolds. To overcome the shortcomings of existing crosslinking agents, we used a novel crosslinking method that combined two crosslinkers (ethanol and genipin) to achieve less cytotoxicity. The present findings showed that increasing the SF content of scaffolds not only led to a several-fold increase in the modulus but also resulted in a longer degradation time and altered morphology (Fig. 2). We investigated the chondroinductive properties of different scaffolds loaded with BMSCs and found that BMSCs on the 1:2 SF-ECM scaffold exhibited a substantial proportion of chondrogenic-differentiation phenotypes. In vivo studies in athymic nude mice transplanted with different scaffolds seeded with BMSCs demonstrated the biocompatibility of SF-ECM scaffolds, as well as their ability to promote cartilaginous matrix formation in subcutaneous tissues. Together, these results demonstrated that the 1:2 SF-ECM scaffold supported cartilage regeneration in vitro and in vivo, which opens up the possibility of using ECM combined with SF as an off-the-shelf strategy for joint cartilage regeneration.
Both costal cartilage and articular cartilage are hyaline cartilage. 52 Costal cartilage shows phenotypic and histological similarities to articular cartilage. 53 In previous studies, costal cartilage can be another source for autologous graft in the reconstruction of articular cartilage.53–57 Moreover, costal cartilage is abundant and widely available compared with articular cartilage. Hence, in this study, we chose costal cartilage as a raw material of scaffolds for articular cartilage repair.
Studies on development of cartilage tissue-engineered scaffolds to drive the differentiation process toward chondrocytes and promote cartilage regeneration have largely focused on altering the biochemical ingredients of scaffolds. To date, many natural derivatives have been used as biocompatible materials for tissue engineering and regeneration. 58 Cartilage-derived ECM proteins have been shown to be well-conserved among different species and might, therefore, be feasible for use as biocompatible scaffolds for MSC attachment, proliferation, and chondrogenic differentiation without exogenous growth factors.22,59 However, owing to their poor mechanical properties, ECM scaffolds might not satisfy the requirements for clinical application. In humans, the compressive modulus of articular cartilage is 0.5–7.7 MPa.60,61 Almeida et al. altered cartilage-ECM slurry concentrations for the purpose of elevating its mechanical modulus. 24 Zhang and colleagues introduced PLGA as one of the components to enhance the mechanical strength of an ECM scaffold. 62 In the present study, ECM scaffolds combined with SF exhibited remarkably high mechanical strength (Fig. 2b, c), which is in agreement with the findings of previous studies. 38 However, the degradation time increased correspondingly with increasing SF concentrations in the mixed scaffold. As show in Figure 2e, at an SF-ECM mass ratio of 1:1, the degradation time of the scaffold was too long to allow cartilage regeneration, while its compressive modulus exhibited little improvement.
Contrary to our expectations, increasing the SF:ECM ratio in the certain range promoted chondrogenic differentiation of BMSCs (Figs. 4 and 5). We assumed that not only the ECM content of a scaffold but also its mechanical strength would play an important role in the chondrogenic differentiation behavior of BMSCs. Even-Ram et al. showed that matrices with different mechanical strength properties favored the differentiation of MSCs into different lineages of cells. 63 In the present study, the efficacy of matrix production and chondrogenesis observed with the 1:2 SF-ECM scaffold was more ideal for cartilage tissue engineering than that observed with the ECM or other SF-ECM scaffolds (Figs. 7 and 8). In addition to their increased mechanical strength, the SF-ECM constructs exhibited significantly high secreted collagen and GAG content in vitro (Fig. 4c, e). However, owing to a marked decrease in ECM concentration, the 1:1 SF-ECM scaffold accumulated relatively less matrix content. In the present in vivo study, the ability for matrix production varied among the different types of scaffolds, with the 1:2 SF-ECM scaffold exhibiting greater matrix production than the 1:1 and 1:4 SF-ECM and pure-ECM scaffolds (Fig. 7). Immunohistochemical findings (Fig. 8) showed increased COL II and decreased COL I content in all scaffolds, especially in the 1:2 SF-ECM construct. The results of safranin O and Masson's trichrome staining, indicating the proportions of proteoglycan and collagen content, respectively, revealed the highest GAG and collagen synthesis activities in the 1:2 SF-ECM scaffold. These results suggested that differences in ECM concentration and mechanical strength might contribute to the varied yield of ECMs in different scaffolds.
To prevent the contraction of the SF-ECM scaffold, we incorporated a new crosslinking step, which instead of using toxic crosslinking methods, incorporated a combination of two crosslinkers (ethanol and genipin) into the scaffold fabrication procedure. Genipin, an alcohol soluble crosslinker, helped crosslink the amino groups of different silk and collagen chains.64–66 Ethanol treatment promoted the transformation of the scaffold structure from random coils to antiparallel β-sheets (Fig. 1a), which offered better mechanical stability to the construct. An optimum scaffold will correspond to specifications such as ideal pore size for cell attachment and suitable three-dimensional structure for nutrient transport and cell proliferation.67,68 By crosslinking different ratios of ECM and SF in a 0.5% genipin-ethanol solution for 3 days, we were able to produce different constructs (1:4, 1:2, and 1:1) that were microscopically diverse from each other. Similar to the ECM scaffold, the SF-ECM scaffolds were highly porous (94.23–97.54%). Relative to the original ECM scaffold, the scaffold with the higher SF concentration exhibited smaller pore sizes (Table 2).
These differences in morphology and pore size can be explained by the crystallization of ice during lyophilization. 69 The pore sizes of different scaffolds mainly depend on the distribution of water in the initial ECM suspension and SF solution. In the frozen ECM suspension and SF solution, ECM particles are relatively larger than SF particles, which might contribute to mass crystallization of ice in a frozen ECM suspension. Considering that the properties of this initial solution (6.9% each of ECM suspension and SF solution) contributed to the pore size of the scaffolds, it is feasible that an increase in SF content induced a smaller pore size. Scaffolds with smaller pores provide a larger surface area for MSC attachment after seeding.70,71
For extensive data on these ECM and SF-ECM scaffolds, in vivo experiments were performed to demonstrate the chondrogenesis ability in nude mice (Figs. 6–8). There exist possibilities that cells from nude mice migrate into scaffolds and implanted cells migrate outside the scaffolds which may be attributed to the inappropriate cell carrier. Previous studies have proved that both ECM and silk are suitable for cell reservation in nude mice. Yang et al. used fluorescent-labeled MSCs in ECM scaffolds and found that the cells in neotissues were derived from implanted BMSCs rather than cells from nude mice. 30 Kim et al. showed that cell penetration and growth in silk scaffolds did not occur during the entire implantation period in nude mice. 61 However, in our studies, it was difficult to identify whether cells from nude mice migrated into scaffolds. Regardless, all scaffolds showed the capacity of chondrogenesis and its biocompatibility in vivo.
Several limitations of the present study ought to be addressed. First, despite its relatively easy accessibility, costal cartilage slightly differs from articular cartilage in terms of ECM composition. Second, the reason behind the better chondrogenic-differentiation behavior of the 1:2 SF-ECM scaffold needs to be further investigated, because various factors in this study, including the scaffold ECM content, pore size, and mechanical strength, might have contributed to this phenomenon. Third, the fabrication and testing processes for SF-ECM scaffolds should be further developed in a large animal model to ensure better translational clinical application. Despite these limitations, the present in vivo experiments developed in a nude mouse model provided useful information for further investigation in other animals or humans.
Conclusions
In the present study, biomimetic composite scaffolds with different ratios of SF and ECM were fabricated by lyophilization. These SF-ECM scaffolds of different component ratios varied in size and structure and possessed ideal porosity and water-uptake capacities. They also possessed greater mechanical strength than the ECM scaffold. In vitro results demonstrated that these SF-ECM scaffolds possessed low toxicity and promoted chondrogenic differentiation of BMSCs. In vivo results demonstrated the biocompatibility of the SF-ECM scaffolds, as well as their ability to form cartilaginous matrix in subcutaneous regions in nude mice. In conclusion, according to our results, scaffolds containing optimal concentrations of ECM and SF are potential candidates for application in cartilage tissue engineering.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (No. 81472079, No. 81672169).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.