
Abstract
Conditioned medium (CM) and microvesicles (MVs) are produced using different protocols: CM is collected following 12–96 h of cell culture without renewal of tissue culture medium, while MVs are collected after overnight cell culture. For future comparative studies in regenerative medicine looking at the efficacy of CM and MVs, it is important to understand how the quality of cell secretions is affected by culture. The aim of this study was to evaluate whether the duration of culturing influences the micro-RNAs (miRNAs) cargo of equine amniotic mesenchymal cells (AMCs) and their MVs. The analysis identified 990 miRNAs. After one night, there were 347 differently expressed (DE)-miRNAs between MVs and cells, whereas after four nights there were 359. About 58.3% of the DE-miRNAs were shared between samples produced under the two conditions. The comparison between miRNA content in AMC cells cultured for one night versus four nights showed eight DE-Equus caballus (eca)-miRNAs, which target genes were involved in immune response to external stimulus, inflammatory response, and production of reactive oxygen species. Comparing MVs isolated from one or four nights, four DE-miRNAs that target genes regulating cell cycle progression and production of reactive oxygen species were found, but only eca-miR-214 was enriched in the MVs after four nights. In conclusion, after 4 days of cell culture, the profile of AMC miRNAs was altered, indicating a probable phenotypic transition versus a new cell culture environment and aging. After this time, MVs accumulated eca-miR-214, which may help cells survive or adapt to new culture conditions.
Impact Statement
Future comparative studies of regenerative medicine using conditioned medium or microvesicles will be difficult to interpret because there is no information about the quality of cell secretions and of the cargo of miRNAs. Microvesicles usually are isolated after one night of culturing, while conditioned medium is collected after a longer culture time. Results show that after four nights of culturing, cells select miRNAs that may reflect a mechanism to promote cell survival or adaptation to the new culture condition (exhaustive medium). Therefore, potentially, the stress of four nights of culturing could make the cells more responsive to an adverse environment.
Introduction
I
Materials and Methods
Chemical reagents, cell culture media, and supplements were obtained from Sigma unless otherwise specified, and tissue culture dishes were purchased from Euroclone.
Tissue collection and amniotic mesenchymal cell isolation
Three allanto-amniotic membranes were collected at the end of mare pregnancies and kept at 4°C in sterile saline solution (0.9% NaCl) supplemented with 4 mg/mL amphotericin (Euroclone, Milan, Italy), 100 UI/mL penicillin or 100 mg/mL streptomycin (Euroclone), and then processed within 12 h. Amniotic membranes were mechanically separated from allantois, and amniotic cells (AMCs) were isolated as described by Lange-Consiglio et al.
24
Briefly, amniotic membrane samples were fragmented and incubated for 9 min at 38.5°C in PBS supplemented with 2.4 UI/mL dispase (Becton Dickinson, Milan, Italy). After 5–10 min at room temperature in high-glucose Dulbecco's modified Eagle's medium (HG-DMEM; EuroClone), supplemented with 10% heat-inactivated fetal bovine serum (FBS) and 2 mM
Enzymatic digestion product was separated from undigested product using a 100 μm cell strainer. Amniotic cells were obtained by centrifugation at 200 g for 10 min and cultured in HG-DMEM supplemented with 10% FBS, penicillin (100 UI/mL), streptomycin (100 mg/mL), 0.25 mg/mL amphotericin B, 2 mM
CM production and MV isolation
CM was obtained by culturing cells in ultra-culture serum-free medium (Euroclone) under two different conditions: AMCs were cultured for either one night or four nights without replacement of medium. Then CM was centrifuged at 2000 g for 20 min to remove any debris. Subsequently, the supernatant obtained was ultra-centrifuged at 100,000 g (Beckman Coulter Optima L-100K ultracentrifuge) at 4°C for 1 h. Serum-free medium 199 containing 25 mM N-2-hydroxyethylpiperazine-N-2-ethanesulfonic acid (HEPES) was used to wash the pellet obtained, and this was centrifuged again using the same conditions. Then the final pellet was resuspended in HG-DMEM. A portion of pellet was stored at −20°C to perform molecular biology studies. Five microliters of each sample was diluted in sterile saline solution (0.9% NaCl) to a final volume of 1 mL and analyzed using Nanosight LM10 instrument (Nanoparticle Tracking Analysis; Nano-Sight Ltd., Amesbuty, United Kingdom) to determine the size and concentration of MVs.
RNA extraction and library preparation
Total RNA was extracted from cells and MVs obtained under the two experimental conditions (one night and four nights) using a NucleoSpin® miRNA kit (Macherey-Nagel, Germany) in combination with TRIzol® protocol and isolation of small and large RNA in one fraction (total RNA). To define the quality and concentration of RNA, Agilent 2100 Bioanalyzer (Santa Clara, CA) was used. Then RNA was preserved at −80°C.
RNA libraries were obtained using a TruSeq Small RNA Library Preparation Kit (Illumina). Libraries from AMCs and MVs from each experimental condition were then united and purified with a Pippin Prep system (Sage Science, MA) to separate the fraction containing miRNAs. Quality and yield of libraries was determined with High Sensitivity DNA Chip (Agilent 2100 Bioanalyzer).
Sequencing and bioinformatics analysis
Sequencing of the final library was run on an Illumina Hiseq 2000 (San Diego, CA) 50 bp paired-end lane. The Mirdeep2 software was used to identify known and novel miRNAs, and the MirBase (
Results
AMC isolation
Cells were able to adhere to the plate (Fig. 1). After AMC isolation, cell vitality was 90% as confirmed by Trypan blue exclusion. Previous molecular biological analyses showed that this cell line exhibits fibroblast-like morphology and stromal phenotype, with the expression of markers such as CD22, CD44, CD106, CD105, and MHC1, but not CD34 and MHC2. 24 Moreover, these cells demonstrated the ability to differentiate in mesodermic and ectodermic lines. 24

Morphology of AMCs. Magnification, 20 × . AMC, amniotic mesenchymal cell.
MV isolation and measurement
After one-night culturing, MVs ranged from 50 to 670 nm, with a mean of 123.2 ± 43.2 nm for the three samples (Fig. 2A). Dimensions of MVs obtained after four nights of culturing varied from 50 to 1000 nm with a mean of 242.8 ± 15 nm for each sample. After four nights of culturing, apoptotic bodies (∼1000 nm) were present (Fig. 2B). A previous transmission electron microscopy study demonstrated that MVs were mostly released by budding and thus classified as shedding vesicles. 12 The MV size confirms the validity of this classification.
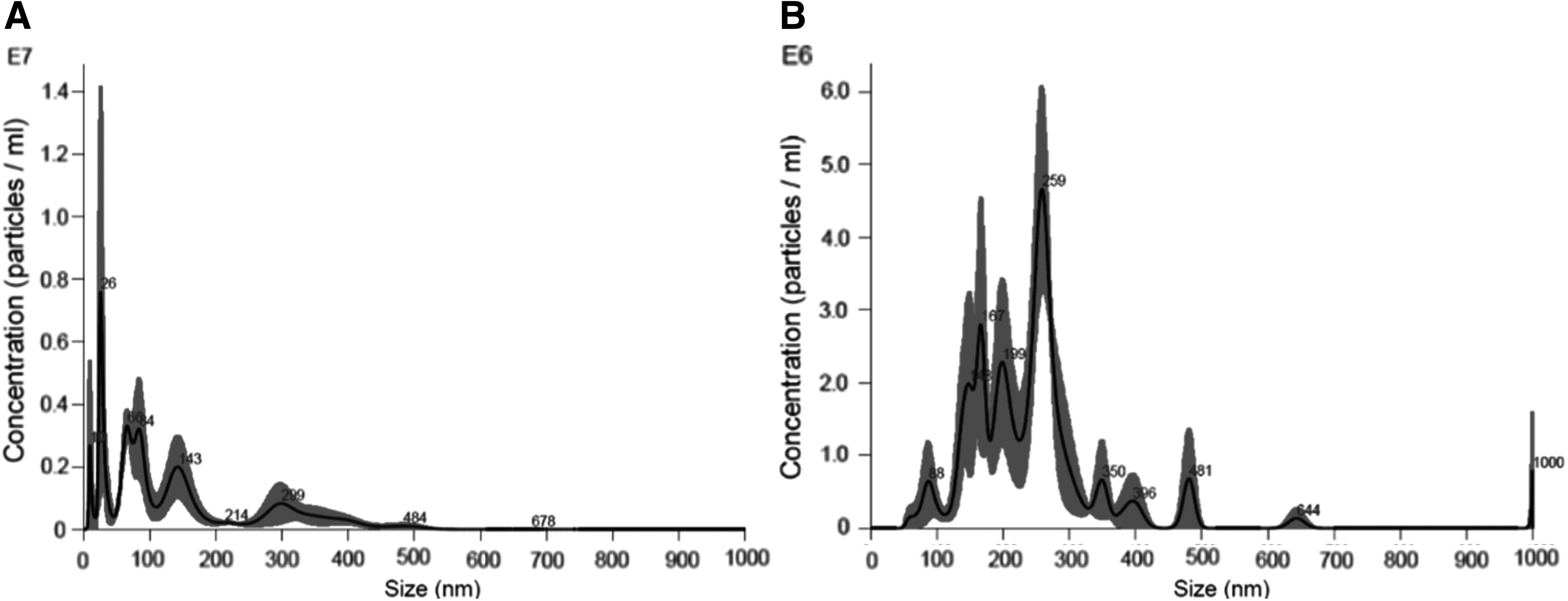
Results of Nanosight evaluation.
The concentration of MVs released after one or four nights of culturing was very similar (mean value of 406 × 10 6 and 448 × 10 6 particles per milliliter of medium for one night and four nights, respectively).
miRNA analysis: extraction, libraries and sequencing
miRNA sequencing provided about 8.4 and 37.6 million reads for cells and MV samples, respectively. About 41.2% and 1.2% of the total reads were identified as miRNAs using Mirdeep2 in cells and MVs, respectively. The small RNA profile according to read size confirmed that in MVs only a small proportion of reads were assigned to miRNAs, whereas a high percentage probably represented other non-coding RNAs, for both experimental conditions (one night and four nights; Supplementary Fig. S1; Supplementary Data are available online at
In total, 990 miRNAs were identified. Among these, 361 were classified as eca-miRNAs (with homology to Equus caballus-known miRNAs), while another 233 were found to be similar to miRNAs from related species, and 396 were considered novel. Principal component analysis (PCA) of all the expressed miRNAs showed that miRNAs from cells and MVs clearly clustered separately (PC1), and also, samples from one-night and four-night culturing showed a typical pattern of miRNA expression (PC2 and PC3; Fig. 3).
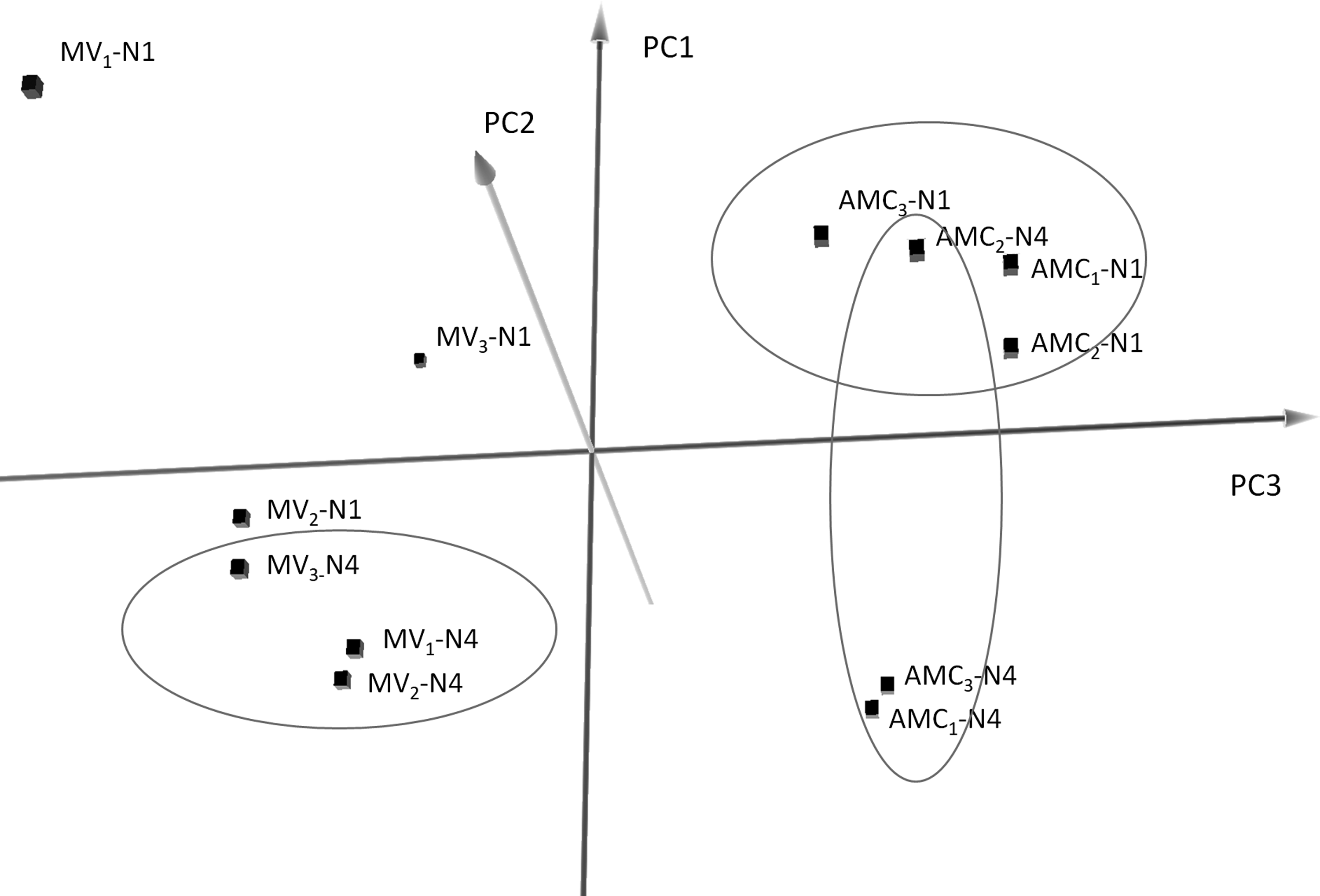
PCA considering all 990 miRNAs expressed in AMCs and MVs (isolates 1, 2, 3 subscripted) from one (N1) and four (N4) nights of culturing, for each replicate (H1, H2, H3). PCA, principal component analysis.
miRNA expression analysis and comparisons
AMCs versus MVs one night and AMCs versus MVs four nights
After one-night culturing, 347 DE-miRNAs were identified in MVs and cells comprising 122 eca-miRNAs, 88 miRNAs with similarity to miRNAs from similar species and 137 novel miRNAs. After four nights, 359 DE-miRNAs were identified, 124 of which were eca-miRNAs, 92 were miRNAs similar to miRNAs from related species, and 143 were novel miRNAs (Supplementary Table S1). About 58.3% of the DE-miRNAs were shared between samples produced under the different conditions. The 26 eca-miRNAs enriched at least twice in MVs compared with cells (log fold change [LogFC] >2), common to both conditions, are: eca-miR-1291a, eca-miR-141, eca-miR-142-5p, eca-miR-144, eca-miR-1468, eca-miR-146a, eca-miR-150, eca-miR-184, eca-miR-200b, eca-miR-200c, eca-miR-205, eca-miR-212, eca-miR-223, eca-miR-423-3p, eca-miR-423-5p, eca-miR-429, eca-miR-451, eca-miR-485-5p, eca-miR-486-3p, eca-miR-486-5p, eca-miR-507b, eca-miR-671-3p, eca-miR-8917, eca-miR-8928, eca-miR-8986a, and eca-miR-92b (Supplementary Table S2). Considering these 26 common DE-miRNAs between cells and MVs, regardless of the culturing time, 13 of these target 171 experimentally validated genes. Gene ontology analysis (p = 10–6) revealed that many of the targeted genes are involved in the regulation of anti-inflammatory response (Fig. 4). Novel miRNA precursors and mature fasta sequences are available in Supplementary Tables S3 and S4.

Pie chart showing the distribution of relative abundance of biological processes obtained from GO analysis for eca-miRNA target genes (19/26 eca-miRNA) found to be commonly overexpressed in MVs after one and four nights of culturing. **p < 0.001. GO, gene ontology.
AMCs one night versus AMCs four nights and MVs one night versus MVs four nights
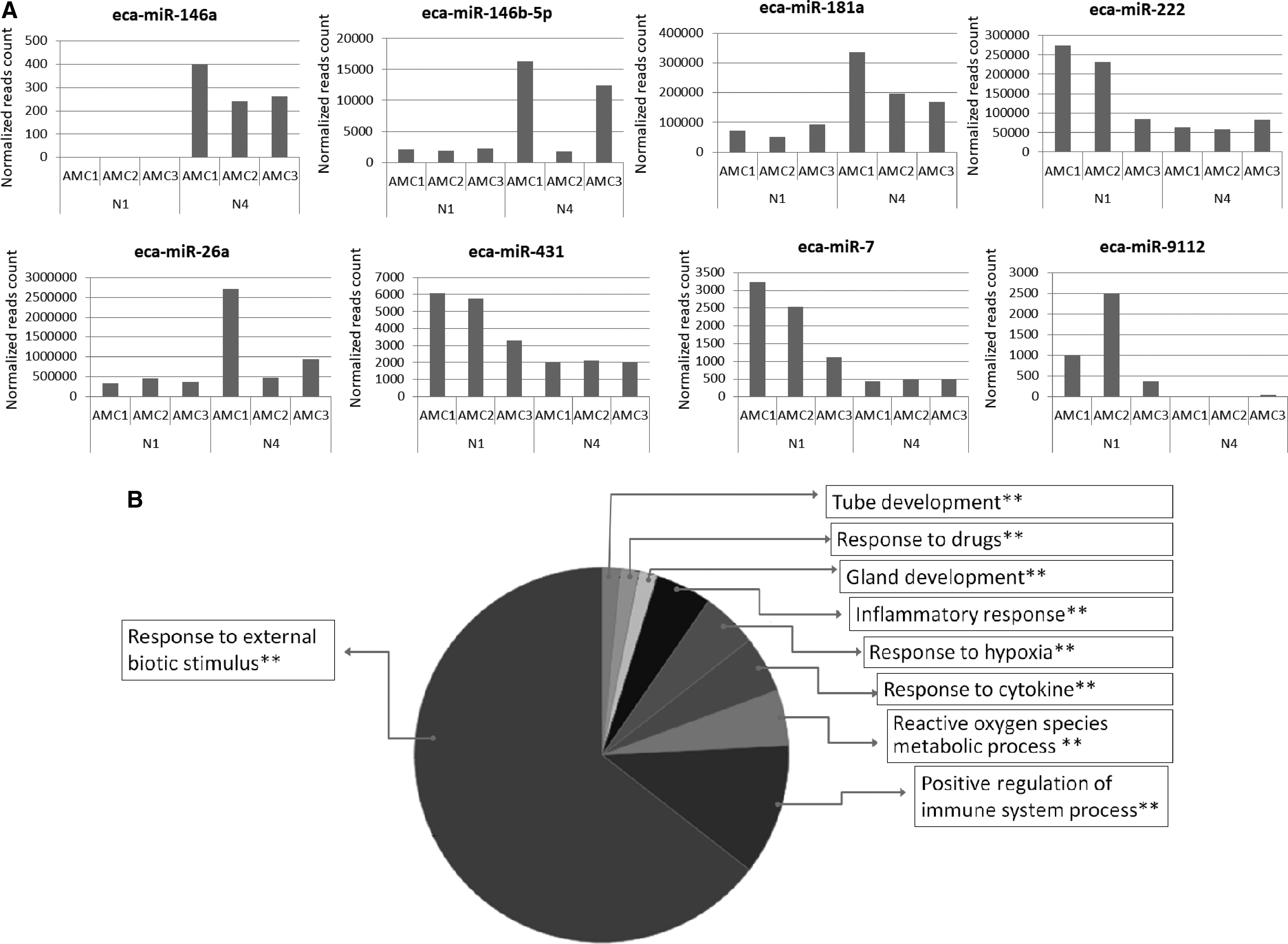
The comparison between miRNA content in AMC cells produced from one night versus four nights of culturing showed 21 DE-miRNAs. Among these, eight were eca-miRNAs (eca-miR-146a, eca-miR-146b-5p, eca-miR-181a, eca-miR-222, eca-miR-26a, eca-miR-431, eca-miR-7, eca-miR-9112) targeting 143 genes with functions related to the immune response to external stimulus, inflammatory response, and production of reactive oxygen species. eca-miR-146a, eca-miR-146b-5p, eca-miR-181a, and eca-miR-26a were overexpressed, whereas eca-miR-222, eca-miR-431, eca-miR-7, andn eca-miR-9112 were underexpressed after four nights of cell culturing (Fig. 5).
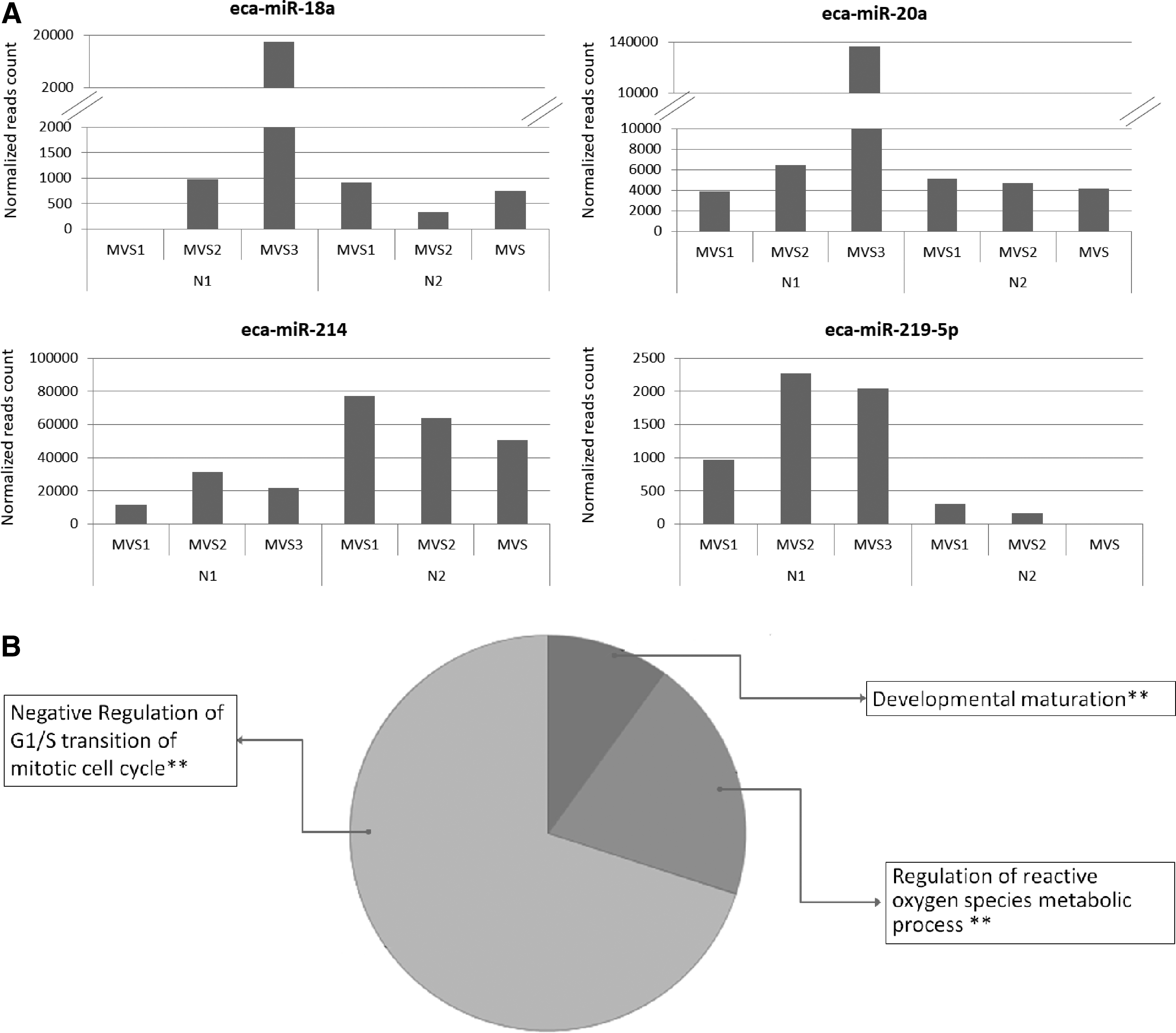
The MVs isolated from one or four nights of cell culturing also showed 13 DE-miRNAs. Among them, four eca-miRNAs (eca-miR-18a, eca-miR-20a, eca-miR-214, eca-miR-219-5p) targeted 64 genes whose main function is related to negative regulation of cell cycle progression and regulation of production of reactive oxygen species. Only eca-miR-214 was enriched in the MVs obtained from cells cultured for 4 days (Fig. 6).
Discussion
It is well known that MVs are captured by the target cells transferring molecules (mainly miRNAs) that can modify the metabolism of target cells.12,27–29 In a previous article, Lange-Consiglio et al. 13 showed that MVs derived from AMCs shed vesicles, and, without sorting between shedding vesicles and exosomes, the expression profile of miRNAs showed a pattern of miRNAs in MVs similar to that of the cells of origin. However, selected miRNAs were detectable in higher concentrations in the released MVs, so it is conceivable that miRNAs are packaged in MVs by a selective mechanism before their secretion. Might the culture time influence this compartmentalization?
Our results showed that the MV size increased from 123.2 ± 43.2 nm after one night of culturing to 242.8 ± 15 nm after four nights for the presence of apoptotic bodies as revealed by the Nanosight Instrument. Some studies have shown that the apoptotic bodies carry some miRNAs regulating specific genes30,31; so in this study all MVs were studied for their miRNA content without any selection. During prolonged cell culturing (four nights), at which time cells consume the nutrients in the growth medium, one or more of the nutrients is depleted and the cells reduce their proliferation. Usually, mammalian cells in culture deteriorate after about two doubling times. 32 The AMCs at P3 have high plating efficiency with an initial lag phase of 22 h, and a prolonged log phase (period where the cells proliferate exponentially) of 11 days. 24 Moreover, equine AMCs kept in high-density culture develop a spheroid structure with no signs of cell contact inhibition. 24 It is conceivable, therefore, that after four nights of culturing without renovation of medium, AMCs continue proliferating even after they reach confluence, but they can suffer generating apoptotic bodies.
The concentration of MVs released after one night or four nights of culturing was very similar. It is possible that during four nights of culturing: (i) the secreted MVs degenerate in the exhaustive medium and are replaced by “new” secreted MVs; (ii) a balance between “old” endocitate MVs and “new” secreted MVs is established.
In the end, the number of vesicles present after four nights is similar to those after one night, and four nights of culturing did not result in a larger quantity of MVs.
Interestingly, comparing AMCs versus MVs one night and AMCs versus MVs four nights
Notwithstanding, PCA separated AMCs cultured for one and four nights and MVs isolated after one and four nights, showing specific miRNA profiles in these two time-points, which were non-overlapping but depended on culturing time. Our analysis revealed differences in miRNA content between AMCs cultured for one and four nights with changes that could influence the response to inflammation and to external stimulus such as hypoxia. Four eca-miRNAs (eca-miR-146a, eca-miR-146b-5p, eca-miR-181a, and eca-miR-26a) were overexpressed, whereas another four eca-miRNAs (eca-miR-222, eca-miR-431, eca-miR-7, eca-miR-9112) were underexpressed after four nights of cell culturing. Some of the miRNAs that have overexpressed in AMCs cultured for four nights (eca-miR146a, eca-miR146b, and eca-miR181a) have previously been observed to be related to altered cell homeostasis. miRNA-146a and miRNA-146b overexpression has been previously reported to induce dendritic cell apoptosis and cytokine production by targeting TNF Receptor Associated Factor 6 (TRAF6) and Interleukin 1 Receptor Associated Kinase 1 (IRAK1) in man. 33 miR-181 upregulation inhibited proliferation and promoted apoptosis of chondrocytes by targeting Phosphatase and tensin homolog (PTEN). 34 Changes in miRNA profile were previously observed in endothelial cells (ECs) with differences related to tissue of origin, cultured cells, and number of passages during sub-culturing, with alteration in senescence-associated miRNAs, probably reflecting cell exposition to changes in culture environment. 35
When the content of miRNAs in MVs was compared between one and four nights, four eca-miRNAs that regulate cell cycle progression and production of reactive oxygen species were observed to change their expression levels.
To note, only a single eca-miRNA (eca-miR-214) was observed to accumulate in MVs after 4 days of culturing. Specific accumulation and shuttling of miR-214 through exosomes was observed between human hepatic stellate cells (HSCs) and human HepG2 hepatocytes, resulting in the suppression of genes related to fibrogenesis. 36 In addition, miR-214 was observed to be present in the EC-derived exosomes and to promote angiogenesis both in vitro and in vivo by repressing ataxia telangiectasia mutated (ATM) expression in neighboring target cells. 37 Expression of miR-214 was observed to increase with age in human bone tissue. 38 Recently, autophagy-related protein 12 (ATG12-mediated autophagy) was observed to be inhibited by miR-214 in colorectal cancer. 39 Extracellular vesicles released by melanoma cells evidence the unique nature of miRNA cargos, including miR-214 accumulation. 40 miR-214 seems to have many roles also in tumorogenesis by coordinating proliferation, angiogenesis, invasiveness, resistance to chemotherapy, and modulating microenvironment. 41 In contrast, in breast cancer cells, miR-214 promotes apoptosis and sensitizes cells to doxorubicin. 42
To summarize, the physical characteristic of MVs (presence of apoptotic bodies), the analysis of miRNA cargo in AMCs, and the enrichment of specific miRNA in the MVs, including eca-miR-214, suggests that these cells could undergo oxidative stress and alterations in cell cycle after 4 days of culturing. It has previously been observed that the miRNA cargo of cells change in function of aging, oxidative stress, and cellular microenvironmental changes.43,44
Potentially, the stress of four nights of culturing could make the cells more responsive to an adverse environment. During in vitro culturing, cells have an optimal environment for pH, nutrient availability, and controlled atmosphere, but the inhospitable microenvironment of injured or degenerating tissues may result in the death or apoptosis of a large proportion of implanted MSCs immediately post-transplantation. 45
Many recent studies have demonstrated that MSCs can be activated or conditioned by a variety of inflammatory stimuli (treatment with interferon-γ or tumor necrosis factor-α) that enhance their immunomodulatory function46–48 and, probably, their responsiveness and engraftment in injured tissue. Inflammatory priming before clinical application might improve the therapeutic effect of MSC treatments. 49 Since miRNAs incorporated into cells after four nights of culturing are involved in responses to external biotic stimuli, drugs, hypoxia, cytokines, and inflammatory response, it is conceivable that these cells may be better able to adapt to an inhospitable environment. The results of this conditioning could be a greater rate of engraftment and more responsiveness of cells to injured tissue. Obviously, these hypotheses require validation in future studies.
Conclusions
After 4 days of cell culturing, the miRNA profile of AMCs changed, indicating a probable phenotypic transition that underlines the ability of the cells to adapt versus a new cell culture environment (medium exhausted) and aging. The MVs released by AMCs change their size and accumulate specific miRNAs, including eca-miR-214, and this may reflect a mechanism to promote cell survival or adaptation to the new culture condition. These results do not allow us to make conclusions about which experimental condition is better for the isolation of MVs. Certainly, in vitro and/or in vivo comparative studies will be needed to evaluate their effectiveness.
Footnotes
Acknowledgments
The research was supported by grants from Università degli Studi di Milano (Milano, Italy) by MIUR GenHome project “Technological Resort for the Advancement of Animal Genomic Research” and by the Italian Ministry of Education, Universities, and Research for the project “Progetto Bandiera INTEROMICS—Sottoprogetto 1: Sviluppo di Infrastrutture Bioinformatiche per le applicazioni OMICS in Biomedicina.”
The authors wish to thank Dott.ssa Maria Chiara Deregibus and Prof. Giovanni Camussi (Department of Internal Medicine and Molecular Biotechnology Center, University of Torino, Torino, Italy) for their skilled assistance in Nanosight use.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.