
Abstract
Despite recent studies on media additives, the low viability and dysplastic features of terminally mature erythroid cells are still major problems in enhancing erythroid cell yields in vitro. Moreover, research on enhancing terminal erythropoiesis has been focused on the immature stage of erythroid cells such as burst forming unit-erythroid and colony forming unit-erythroid. Here, we tested many commercially available serum-free culture media and developed a superior culture media formulation compared with the conventional control for higher expansion-fold, higher viability, and therefore enhanced red cell productivity. The addition of the specific medium to the previously known best media at a specific ratio, whose effects were not dose-dependent, enabled the generation of significantly higher erythrocyte products with over 1.3 million-fold proliferation of erythroid cells after maintenance for 21 days throughout the maturation stages from CD34+ cells. The cells cultured in this condition expressed maturation markers and were significantly superior in differentiation and enucleation. Comparative mRNA profiling revealed that erythroid cells in this medium showed more efficient maturation in mRNA levels. The cultured cells showed comparable erythroblast survival and also restored better red blood cell (RBC) functions of oxygen saturation profile with expression of adult globin up to 99%. However, to develop chemically defined media, the well-known supplements including hormones, cytokines, and serum-replacement reagents were not sufficient to replace the optimized media in producing mature RBCs. Taken together, our optimized medium formulation under serum-free culture conditions produced the best reproducible results on productivity and maturation in erythroid cells with economic benefits. These culture conditions may thus serve as a useful platform for further investigation of in vitro erythropoiesis and to develop defined serum-free media for clinical trials.
Impact Statement
This article describes a culture method for significantly high erythrocyte production by simply using a commercially available culture media mixture in serum-free and feeder-free conditions. The culture method enables up to 1.3 million-fold expansion of erythroid cells. With optimized medium conditions, erythrocytes showed proper red cell functions with adult hemoglobin at 99%. This simple and reproducible serum-free culture media formulation with economic advantages can be widely used in erythropoiesis research and red cell production.
Introduction
Increasing studies are investigating in vitro red blood cell (RBC) production and terminal erythropoiesis using stem cells to solve the lack of blood supply and donor-related side effects of transfusion.1–3 However, the low viability and productivity in suspension culture conditions without supporting cells such as macrophages and stromal cells need to be improved. Conventional culture media containing serum or plasma show high lot variations and can compromise the clinical applicability of in vitro produced RBC products as well as studies on in vitro erythropoiesis. 4
Hematopoietic stem cells (HSCs) are differentiated to RBCs through the progenitor and precursor stages. Before enucleation at the stage of polychromatic and orthochromatic erythroblasts, proliferation ability is limited by several doublings. 5 Also, the low viability and enucleation rate along with dysplastic changes at this stage have not been improved. 6 The high cost of widely used culture media and serum for culturing HSCs and blood cells is also problematic for the research and development of RBC products. 3 Moreover, erythrocyte yield with higher proliferation and viability has not been successful using serum-free conditions in previous studies. Therefore, there have been numerous reports on enhancing RBC production yields using various media additives ranging from single reagents to fetal bovine serum (FBS).4,7,8 Recently, Trakarnsanga et al. reported that conditioned media of OP9 mouse stromal cells could support erythropoiesis similar to the coculture with OP9 cells. 9 However, these culture systems are difficult to maintain and it is hard to get enough stromal cell numbers for mass erythroid cell culture. Furthermore, the feeder cells are of mouse origin and require FBS in culture with less than 80,000-fold expansion of erythroid cells. For reproducible and robust research, however, serum-free, feeder-free, and chemically defined culture media are required.
To solve these problems, various serum-free culture media have been developed for hematopoietic and blood cell culture including StemSpan™, Stemline®, X-VIVO™, and STEMPRO®-34. These media enable proliferation and maintenance of erythroid cells, even though these were not enough to obtain healthy RBC products for erythropoiesis research. Besides, during final maturation, most of the erythroblasts cannot enucleate and therefore die. Therefore, many laboratories have added human AB serum and plasma to media or have used feeder cells during the enucleation period.10–12 However, human AB serum and plasma are very expensive and unsuitable to produce RBCs in vitro.
Therefore, we aimed to develop a serum-free medium using commercial media with robust erythrocyte yields using frozen cord blood (CB) CD34+ cells for higher erythrocyte yield with better oxygen-carrying function, robustness, economics, and operability. In this study, we established an optimized mixed medium (OMM) in which erythroblasts reproducibly proliferate, maturate, and enucleate more efficiently than previously reported in any other serum-free conditions by easily mixing two commercial media. The described method provides high efficiency, allowing high-throughput RBC production for scientific purposes and can potentially be developed further for blood cell manufacture.
Materials and Methods
Isolation of CB CD34+ cells
Human umbilical CB was collected after obtaining written informed consent from healthy pregnant women. The study was approved by the Institutional Review Board of the Hanyang University in Korea (HYI-16-178-5). Mononuclear cells were collected using Ficoll-Paque. CD34+ cells were isolated through an EasySep Human CD34 Positive Selection Kit (Stemcell Technologies, Vancouver, Canada) according to the manufacturer's protocol. Isolated cells were cryopreserved in a liquid nitrogen tank.
Control culture methods for erythroid differentiation
CD34+ cells were cultured in a three-phase liquid culture system as described in a previous report. 6 Frozen cells were thawed and seeded to 0.5–1 × 105 cells/mL in serum-free Stemline II (Sigma–Aldrich, St Louis, MO). Supplemented reagents are listed in Supplementary Table S1. We designated these supplement reagents as a supplement set.
Establishment of OMM
The CD34+ cells and differentiated erythroid cells were cultured in Cellartis DEF-CS™ 500 Basal Medium (Takara, Kyoto, Japan), Cellartis DEF-CS 500 Xeno-Free GMP Grade Basal Medium (Takara), X-VIVO 15 (Lonza, Walkersville, MD), mTeSR1 (STEMCELL Technologies), Dulbecco's modified Eagle's medium, Nutrient mixture F-12 (DMEM/F12; Gibco), StemFit Basic03 (Ajinomoto, Tokyo, Japan), and Essential 8 (Gibco). The additives in the commercial media package were added following the manufacturer's protocols except for GF3 in DEF-CS, which is used for adherent cell culture. The other culture protocol, supplement set, and cytokines were added as in the control condition, which is not otherwise mentioned in the Results section. The effects of various mixing ratios of Stemline II and DEF-CS were compared. After deciding the OMM mixing ratio, the ratios of supplement set addition were tested.
Treatment with antioxidants and other supplements
Erythroid cells were cultured in Stemline II, Iscove's Modified Dulbecco's Medium (IMDM) (Gibco), or DMEM/F12 (Gibco) media treated with or without the following reagents as indicated in the Results section: resveratrol (Sigma), capsaicin (Sigma), and Tempol (Sigma) as antioxidants, and apo-transferrin (Sigma), human platelet lysate (hPL) (UltraGRO advanced; Helios, Atlanta, GA), recombinant human albumin (Sigma), β-estradiol (Sigma), testosterone (Sigma), fibroblast growth factor 2 (FGF-2; PeproTech, Rocky Hill, NJ), and lysophosphatidic acid (LPA, Sigma). Hormones, transforming growth factor (TGF-beta), and LPA were analyzed by electrochemiluminescence immunoassay, enzyme-linked immunosorbent assay, and liquid chromatography-mass spectrometry (LC-MS), respectively.
Cell morphology analysis
Cells were centrifuged using Cytospin (Cellspin; Hanil Science Industrial, Incheon, Korea) at 900 rpm for 3 min onto a glass slide and stained with the Wright–Giemsa stain (Sigma). Maturation status and dysplastic cells were observed under a light microscope (Nikon Eclipse TE2000-U) in a blinded manner by two experts.
Flow cytometry
After washing with phosphate-buffered saline, cells were labeled with glycophorin A (GPA)-fluorescein isothiocyanate (FITC) (Life Technologies, Gaithersburg, MD), CD71-PE (BioLegend, San Diego, CA), CD11b-FITC (BioLegend), and CD13-PE (BD Biosciences, San Jose, CA) antibodies. Nuclei were stained with NucRed Live 647 (Invitrogen). For hemoglobin staining, cells were fixed, permeabilized, and tagged with fetal hemoglobin (Hb-F)-FITC (BD Biosciences), hemoglobin beta (HBB) (Santa Cruz Biotech, Santa Cruz, CA), GPA-PerCP cy5.5 (BioLegend), and NucRed Live 647 (Invitrogen). Analysis was performed on an Accuri C6 (BD Biosciences). Median fluorescence intensity (MFI) was calculated using BD CSampler Software.
Quantitative real-time polymerase chain reaction
Total RNA was isolated from harvested cells and measured with a NanoDrop (BioSpecNano Spectrophotometer; Shimadzu, Japan). cDNA was synthesized using the SuperScript III Reverse Transcriptase (Invitrogen, ThermoFisher Scientific, Waltham, MA). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed on an Applied Biosystems 7500 Real-Time polymerase chain reaction (Applied Biosystems, Foster City, CA) in duplicate using a SYBR Green reaction kit (Enzynomics, Daejeon, Korea). mRNA expression levels were normalized to the expression of glyceraldehyde-3-phosphate dehydrogenase. The primers are listed in Supplementary Table S2.
Oxygen-binding function assay
Hemoglobin oxygen-binding capacity was evaluated with a Hemox analyzer, Model B (TCS Scientific, New Hope, PA) according to the manufacturer's recommendations.
Coculture of 293T and OP9 cells
293T cells were cultured in DMEM (Gibco) containing 10% FBS (Gibco) and 1% penicillin–streptomycin solution (PS, Gibco). OP9 cells were cultured on 0.1% gelatin-coated (Sigma) plates and in α-modified minimum essential medium (Gibco) containing 10% FBS and 1% PS. Cell seeding densities were 3 × 105 cells/2 mL for 293T cells and 1 × 105 cells/2 mL for OP9 cells in 12 wells. After washing the media for feeder cells, 1 × 105 immature erythroblasts were seeded on the feeder cells and maintained in control conditions (Stemline II). Erythroblasts were harvested every 2–3 days with media changes. Then, reactive oxygen species (ROS) from erythroid cells were measured using H2DCFDA (Invitrogen).
Statistical analyses
Statistical analyses were performed with GraphPad prism (GraphPad, San Diego, CA). Significance was determined using the Kruskal–Wallis one-way analysis of variance, followed by Dunn's post hoc test to compare multiple groups. For two group comparisons, the Mann–Whitney test was performed. A p-value of <0.05 was considered statistically significant.
Results
Screening of commercial serum-free medium for better erythroid cell maturation and proliferation
To develop a serum-free media composition, we searched for commercial serum-free media with abundant nutrients for stem cells like HSCs, induced pluripotent stem cells (iPSCs), and embryonic stem cells and chose eight serum-free media. When compared with Stemline II, which is a serum-free GMP-grade medium, only cells in DEF-CS showed similar proliferation (Fig. 1A). Erythroid cells, especially in DEF-CS, showed high proliferation until day 9 and generated more enucleated RBCs at days 17 and 21 (Fig. 1B, D). However, cells in DEF-CS showed dysplastic morphology such as multinucleation and abnormal nuclei contouring at day 21 (Fig. 1C). DEF-CS 500 Xeno-Free GMP Grade Basal Medium was also tested. However, the cells could not proliferate and thus could not be analyzed (data not shown).
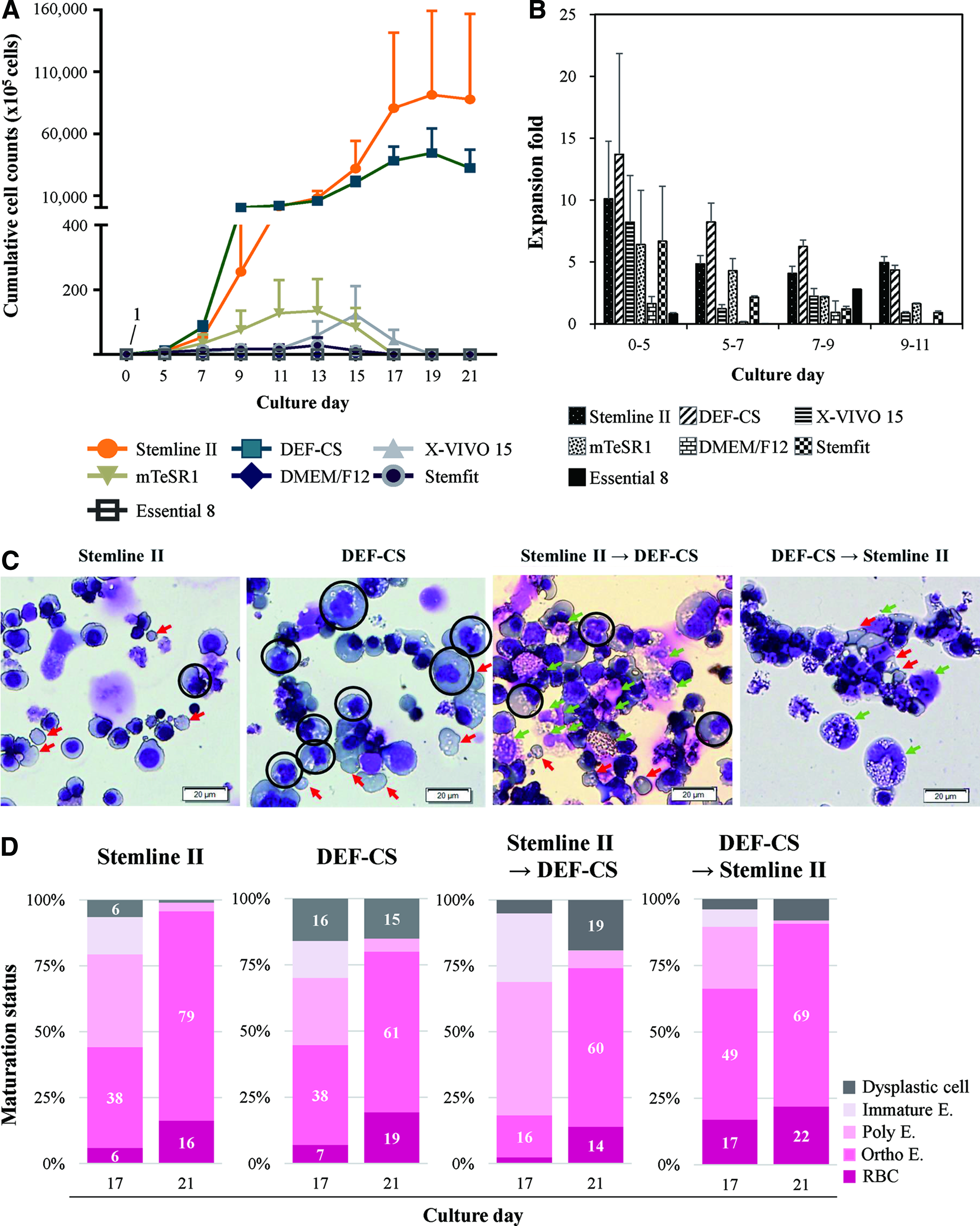
Validation of various serum-free commercial media for erythrocyte production.
Effects of media combination and formulations
To develop better media conditions for high RBC yields, we designed two strategies. First, we changed the medium from Stemline II to DEF-CS when mature erythroblasts came out or vice versa at day 13. However, these methods induced more dysplastic cells than the Stemline II control. The protocol using DEF-CS followed by Stemline II induced more RBC production (Fig. 1D). However, nonerythroid lineage cells such as macrophages were observed more than those with Stemline II (Fig. 1C). Second, we mixed two media at the indicated ratio. Stemline II 90% and DEF-CS 10% showed the highest expansion, with a 4.8-fold increase compared with that with Stemline II at day 21. In this condition, over 1.3 million-fold proliferation was observed from CB CD34+ cells. Stemline II 80% with DEF-CS 20%, Stemline II 100%, and DEF-CS 100% showed similar expansion-fold (Fig. 2A). DEF-CS 500 Xeno-Free GMP Grade Medium was also tested in a mixture with Stemline II (2:8 and 5:5) but showed a very low expansion rate, demonstrating a great difference in erythropoiesis from the counterpart DEF-CS medium (data not shown).

Optimization of media by mixing formulation. CB CD34+ cells were cultured to erythrocytes at the indicated media conditions.
As the DEF-CS medium package contains additional frozen additives (DEF-CSTM 500 additives) that might contain many cytokines and nutrients, we tested whether the other supplement set (Supplementary Table S1) added to the control condition is completely necessary when DEF-CS medium is used. Therefore, we next adjusted the best ratio of the supplement set depending on the mixed media with Stemline II 80% and 90%. The control condition was designated as supplement set 100% in Stemline II 100%. The experimental condition was adjusted from 0% to 100% supplement. As a result, cells in both Stemline II 80% and 90% proliferated more than those in Stemline II 100%. In both conditions, 50% addition of the original supplement set volume enhanced proliferation to the highest (Fig. 2B). At day 19, cells in Stemline II 80% with DEF-CS 20% and 50% supplement were expanded to a mean of 83.3-fold higher than those in Stemline II (Fig. 2C). The enucleation rate at day 19, RBC production rate was higher than other conditions when supplement was not added (Fig. 2D). Based on these results, we selected a mixture of 80% Stemline II and 20% DEF-CS with a 50% supplement set and named this mixture as OMM.
OMM enhances erythroid cell proliferation and maturation
Erythroid cells derived from CD34+ cells were cultured for 21 days in OMM and the control. The mean cell number in OMM was 30-fold higher than that in the control at day 17 (Fig. 3A). Proliferation was maintained until day 15 in the control with decreased viability (lower than 50%). In contrast, the proliferation in OMM was continued until day 17 with higher viability over 50% until day 21 (Fig. 3B).
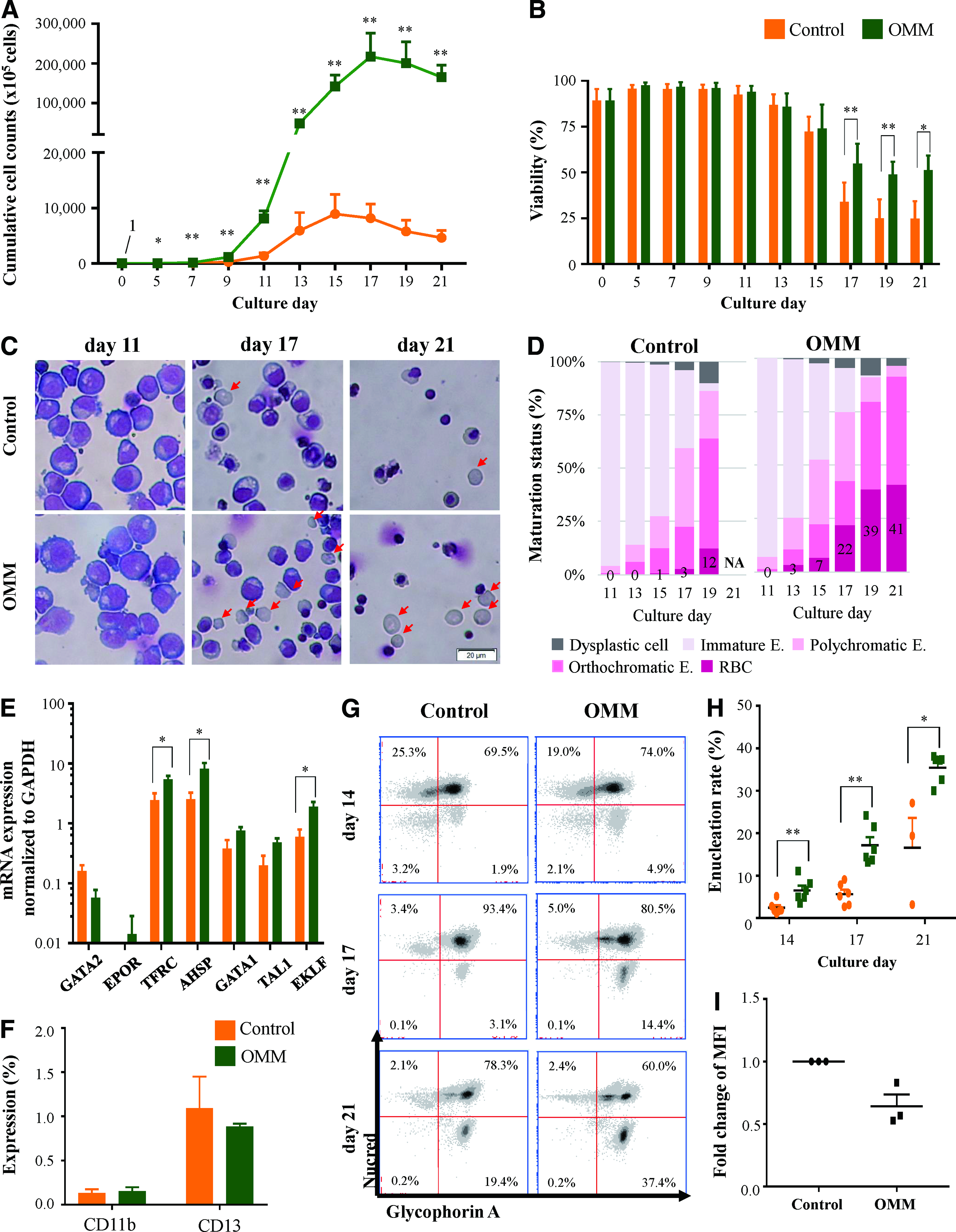
Cumulative cell counts
The cell morphology was good in both the control and OMM during the immature stage until day 11, but dead cells were observed more in the control than in OMM from day 17. The surviving cells on the final culture day showed intact cell membrane in both conditions (Fig. 3C). In contrast to 12% RBCs in the control at day 19, the enucleated RBC rate in OMM was increased until a mean of 41% at day 21. When cultured in a bioreactor using OMM, the enucleation rate was as high as 95% (data not shown). Dysplastic cells were reduced in OMM compared with those in the control (Fig. 3D). As erythroblasts from some CB cases in Stemline II were almost dead, we could not measure their maturation status at day 21. As the proliferation and viability results among controls were different from Figure 1D in that, a different lot of Stemline II was used, Stemline II seems to have large lot variations.
The transcripts of EPOR (erythropoietin receptor), TFRC (transferrin receptor), AHSP (alpha hemoglobin stabilizing protein), GATA1 (GATA-binding protein 1), and TAL1 (TAL bHLH transcription factor 1, erythroid differentiation factor), which increase along with erythroid cell maturation as analyzed by qRT-PCR, were increased in OMM compared with those in the control at day 17. GATA2 (GATA-binding protein 2) that decreases along with erythropoiesis was lower in OMM than in the control (Fig. 3E). These results support that OMM facilitates erythropoiesis at the mRNA level.
Cell surface markers were evaluated by flow cytometry. Erythroid purity was about 99% when measured with monocyte (CD11b) and myeloid (CD13) markers (Fig. 3F). Expression of GPA that is an erythroid-specific marker was similar in the control and OMM (at day 21, control, 97.7%; OMM, 97.4%). The enucleation rate marked by GPA+ and the DNA marker Nucred− were higher in OMM than in the control, confirming the results obtained by the Wright–Giemsa staining (Fig. 3G, H). CD71, which is expressed until the reticulocyte stage and disappears in mature RBCs, 13 showed a 0.6-fold decrease in MFI among GPA+Nucred− cells in OMM than in the control, showing further maturation in OMM (Fig. 3I).
Erythroid cells cultured in OMM show higher level of adult hemoglobin with enhanced oxygen-carrying ability than that in control
The subtypes of hemoglobin in each medium condition were evaluated by flow cytometry. Contrary to the previous report that erythroid cells derived from CB CD34+ cells mainly showed Hb-F and adult hemoglobin (Hb-β) double positivity,14,15 cells in OMM mainly expressed Hb-β up to 98.8% and 96.4% at days 17 and 21, respectively (Fig. 4A). In OMM, cells with only Hb-F positivity were very few and the mean proportion of Hb-β+Hb-F− was increased from days 17 to 21 (Fig. 4B).
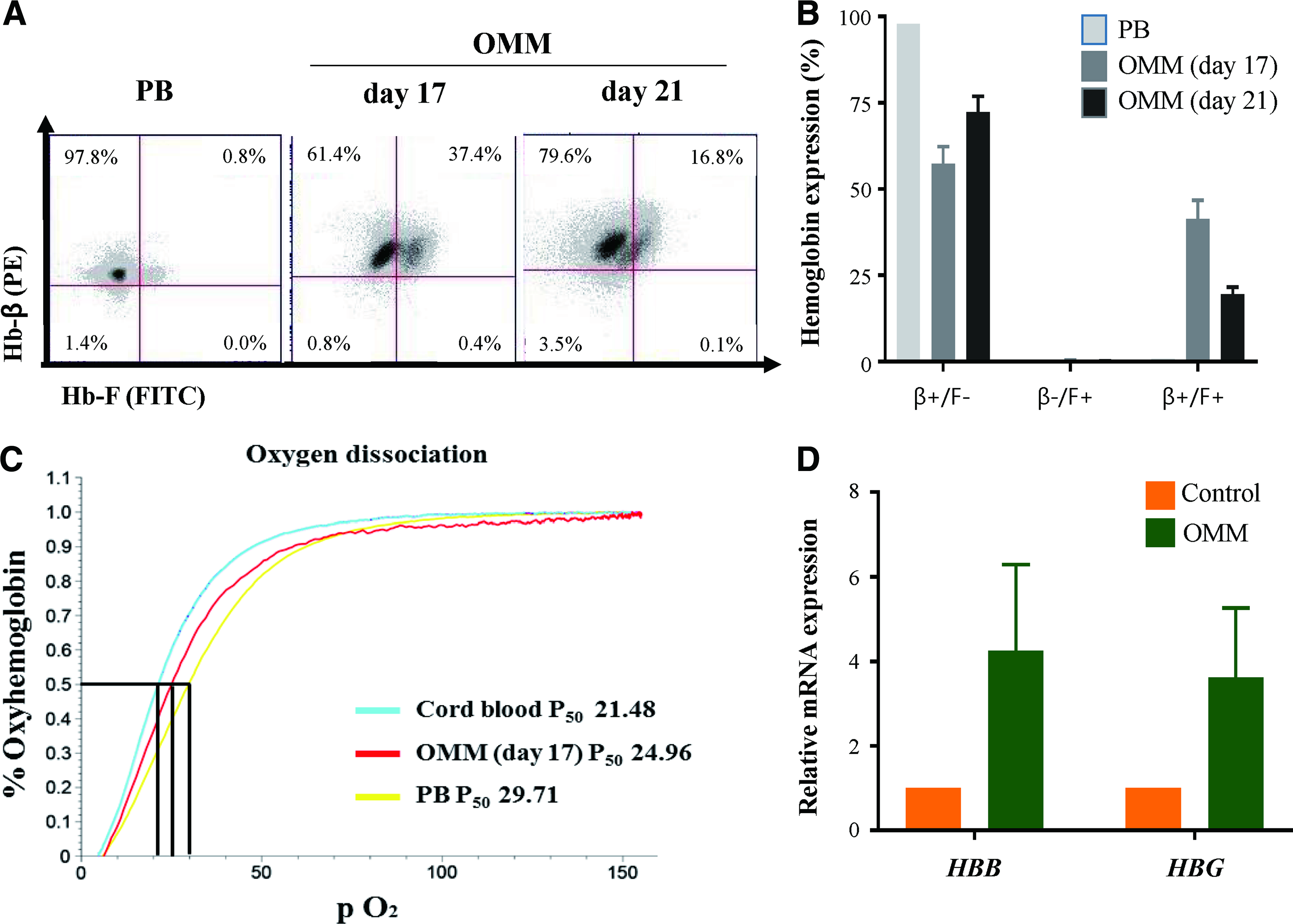
Hemoglobin evaluation from cells in OMM.
The oxygen equilibrium curves of cultured erythroid cells in OMM showed comparable oxygen-binding affinity to peripheral blood RBCs with lower affinity than CB RBCs that is consistent data with higher composition of adult hemoglobin in OMM cells (Fig. 4C). mRNA expression levels of HBB and hemoglobin gamma by qRT-PCR showed 4.25- and 3.63-fold increase in cells in OMM when normalized to the controls (Fig. 4D).
Identification of active media constituents in OMM was not successful
To identify the effective factors in OMM compared with those in the control, 2D-gel electrophoresis was performed using concentrated Stemline II and DEF-CS. Then, the top 13 different spots that existed only in the DEF-CS gel were selected (%Vol >0.2). The spots were analyzed by LC-MS. Most spots were related to albumin. The other top 2 spots were vitamin D-binding protein precursor (Bos Taurus) and mitochondrial coenzyme A transporter SLC2FA42 (mouse). The other spots could not be identified without matched known sequences.
To evaluate the active ingredients of DEF-CS, we selected nine cytokines that were related with erythropoiesis and measured their concentrations using a multiplex cytokine assay. FGF-1, FGF-2, testosterone, estradiol, IGF-1, LPA, and TGF-beta were contained more in DEF-CS than in Stemline II (Table 1). To confirm that these factors were the reason for the superior results in OMM, FGF-2, testosterone, estradiol, and LPA were added to Stemline II at the concentration in OMM that contains DEF-CS. However, the supplements did not reproduce the results of OMM in terms of cell expansion and enucleation (Fig. 5A, B).
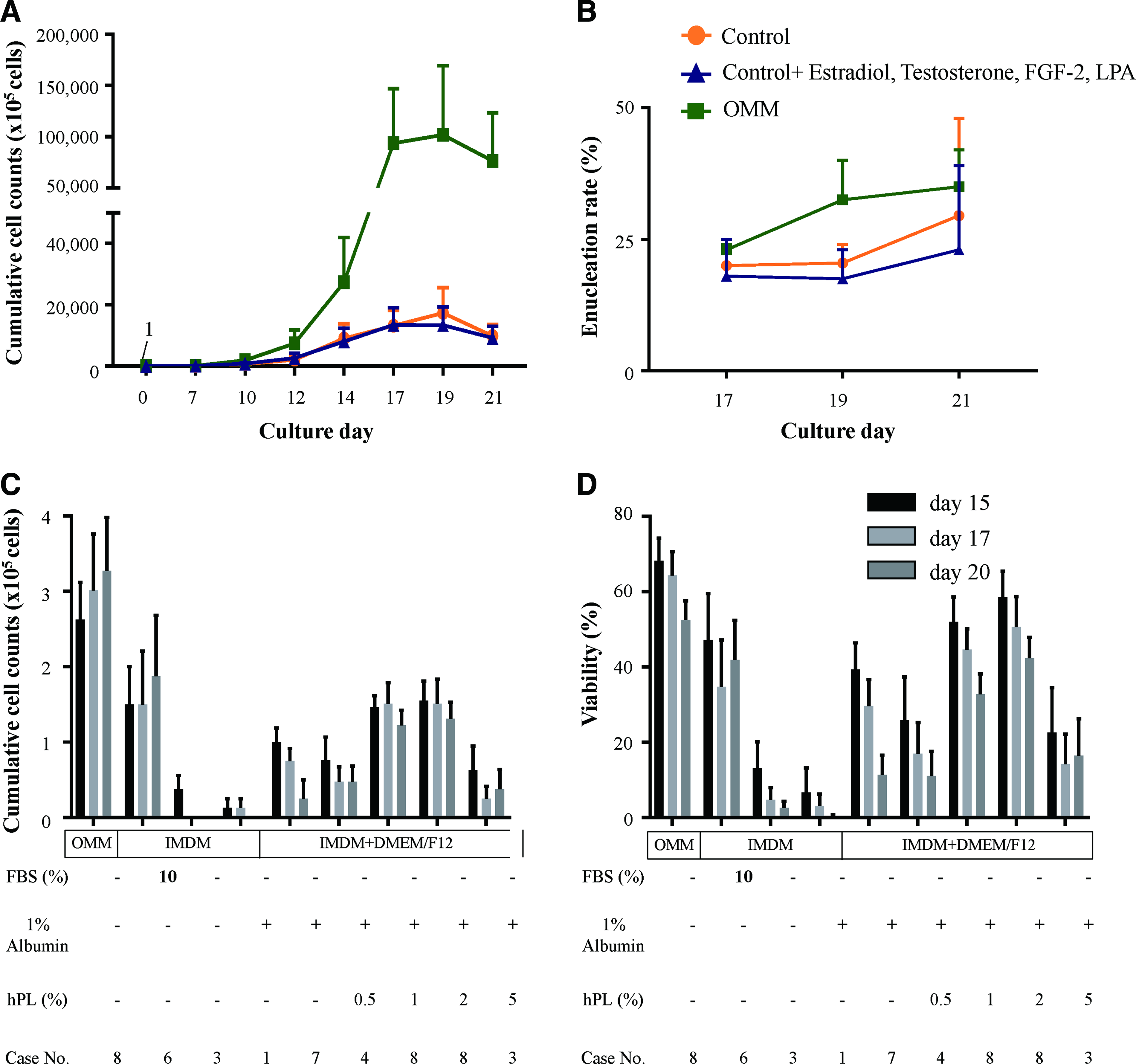
Effects of serum-replacing reagents on erythrocyte production. Control medium (Stemline II) supplemented with estradiol, testosterone, FGF-2, and LPA was compared with OMM and control for cumulative cell counts
Content Analysis of Cytokines, Hormones, and Lysophosphatidic Acid
In pg/mL: FGF-1, fibroblast growth factor 1; FGF-2, fibroblast growth factor 2; BMP-2, bone morphogenetic protein 2; BMP-4, bone morphogenetic protein 4; GM-CSF, granulocyte–macrophage colony-stimulating factor; IL-6, interleukin-6; LIF, leukemia inhibitory factor; M-CSF, macrophage colony-stimulating factor; VEGF, vascular endothelial growth factor; IGF-1, insulin-like growth factor; LPA, lysophosphatidic acid; TGF-beta, transforming growth factor beta.
hPL cannot support erythropoiesis instead of OMM
To advance toward the development of xeno-free medium, we searched for serum replacement reagents. As hPL can be used instead of FBS in mesenchymal stem cell culture,16–18 we evaluated the effects of hPL on erythroid cells in media. Cells were maintained in OMM until day 13 and then changed to mixed media with IMDM and DMEM/F12 containing human recombinant albumin and 0–5% hPL. Cells seeded to 1 × 105 cells/mL at day 13. However, cells could not be maintained with viability less than 50%. Cells in IMDM with 10% FBS showed higher cell counts than other conditions but could not reach that in OMM (Fig. 5C, D). Therefore, the OMM condition could not be replaced by just adding serum or hPL.
Antioxidant supplementation does not provide additional advantages for erythrocyte yields
Cell culture media are usually deficient in antioxidants that are essential for normoxic in vitro culture. Even though vitamin C and other antioxidants like tocopherol were already added to our control condition, we searched for additional antioxidants that could improve the vacuoles seen in the cytoplasm and the low cell viability. We searched previous publications for antioxidants related to erythropoiesis19–21 and referred their adequate concentration ranges. Resveratrol, capsaicin, and Tempol were added to erythroid cells in the control Stemline II medium from day 11 when the cells still showed good morphology and viability. Fresh media and antioxidants were refreshed every other day. After 6 days of addition, no significant effect was observed in terms of cell viability regardless of the antioxidant concentration (Supplementary Fig. S1A). Treatment with both resveratrol and Tempol did not increase cell viability compared with that in the control (Supplementary Fig. S1B).
While erythroid cells are matured, they accumulate iron (Fe) that is supplied from transferrin. As Byrnes et al. observed erythroid expansion through regulation of holo- or apo-transferrin concentration, 22 we changed the transferrin concentration to holo-transferrin 1 mg/mL and apo-transferrin 0.2 mg/mL at day 13. Despite the increased viability by about 10% compared with that in the control, cell morphology and enucleation were not affected (data not shown). Therefore, the beneficial effects of additional antioxidants or iron supplements were doubtful.
Coculturing with stromal cells was not effective for erythrocyte yields
To study the effects of erythroblast coculture with feeder cells on viability and cell yield, erythroblasts cultured in Stemline II until day 7 were transferred to 293T and OP9 feeder cell layers at day 8 with media change. As some erythroblasts were attached to feeder cells, we counted only the detached erythroid cells (Supplementary Fig. S2A). After 5 days of coculture, the GPA portion showed 73.4%, 61.9%, and 73.3% in the control, 293T cocultured, and OP9 cocultured cells, respectively (Supplementary Fig. S2B). After 6 days of coculture, ROS decreased to 0.26-fold when cultured with OP9 cells than in the control (supplementary Fig. S2C). Cocultured erythroblasts proliferated more compared with those without feeder cells until day 12; however, their proliferation was decreased after that time point (Supplementary Fig. S2D). While 293T cells were detached from the plates after coculture probably due to the serum-free media condition, OP9 cells remained attached to the plates and maintained healthy erythroblasts similar to the control (Supplementary Fig. S2E). However, the proliferation capacity in coculture conditions was much less than that in the control.
Discussion
In the present study, we developed serum-free media for increased erythrocyte yield by combining the commercial culture media, Stemline II and DEF-CS, and supplements at a specific ratio. The medium giving the best results was named as OMM and was found to provide markedly robust and efficient results in terms of erythroid cell proliferation, enucleation, and final cell yield with decreased myelodysplasia and increased cell viability, thus overcoming the limitation of previously reported erythroid maturation media. The mean enucleation rate was increased by more than twofold (Fig. 3H) and the mean erythroid cell counts were 35-fold higher (up to 71-fold) with 73-fold more erythrocytes in OMM at day 21 than in the previously reported best serum-free media (control) (Fig. 3A).
Although there are many media for HSC or iPSC production, they are not enough to reproducibly produce large amounts of erythrocytes in serum-free conditions. Attempts to increase the enucleation rate in vitro have been made over decades. Several studies have shown high enucleation, but they used serum,23,24 plasma, 25 or serum substitutes 26 at the final maturation stage. In the process to develop chemically defined GMP grade media for erythrocyte production, we first sought to optimize the media conditions showing highest erythrocyte maturation and yields. Interestingly, the DEF-CS media designed for iPSC culture not only showed good proliferation but also revealed erythroid dysplasia. Instead of dose-dependent effects of the serial ratio of mixing the media, a specific mixing ratio showed very good proliferation as well as the lowest myelodysplasia. Gao et al. revealed that TGF-beta inhibition stimulated RBC production by enhancing the expansion of burst forming unit-erythroid. 27 Therefore, we inhibited TGF-beta by blocker in OMM and confirmed that expansion was increased but enucleation rate was decreased (data not shown). In our study, 20% DEF-CS showed the highest expansion and enucleation; the effect was not dose-dependent. Therefore, we suggest that a certain amount of TGF-beta would be needed to balance proliferation and enucleation. In addition, analysis of the OMM and development of chemically defined GMP-grade media using basal media such as IMDM with previously known reagents including cytokines, antioxidants, LPA, and hPL were not effective to show erythrocyte yields and viability comparable to OMM. Despite the expensive medium for iPSCs, our OMM media including 20% of DEF-CS is cheaper than the control media and provides much higher erythrocyte yields.
Lee et al. established defined serum-free media but that medium revealed very low enucleation. 28 Oliver et al. used small molecules to differentiate human pluripotent stem cells to RBCs but observed a low enucleation rate (under 10%). 29 Lim et al. used the design of experiment-optimized cytokine formulation and found that the optimized set could achieve an average of 26,460-fold total cell expansion. 30
Trakarnsanga et al. 9 showed that OP9 coculture or OP9 conditioned medium could delay the differentiation of erythroblasts, thus resulting in increased expansion of erythroid cells. In our study, OP9 cells maintained healthy erythroblasts similar to the control (Supplementary Fig. S2E). However, after day 12, cocultured erythroblasts did not proliferate anymore (Supplementary Fig. S2D). We suggest that OP9 decreased ROS and maintained erythroblasts better but could not affect the expansion of erythroblasts in serum-free condition.
Even though there are many advantages of using OMM medium, the total RBC yield may fluctuate due to lot variation in Stemline II and variation in the CB samples. This suggests that development of chemically defined media along with constant donor cell lines is very critical.
Therefore, we sought to elucidate some important key factors from DEF-CS. Notwithstanding diverse analysis for proteins, cytokines, and vitamins, we could not substitute OMM by formulations using previously known reagents (Fig. 5A, B). Furthermore, hPL, known as a substitute for FBS in the MSC culture, could not maintain erythroblasts well (Fig. 5C, D). Even though our cell proliferation and cell yield results are excellent, GMP grade media should be developed for clinical use in transfusion. However, our simple and cost-efficient culture methods could provide a robust protocol for laboratory research and use in diagnostics as well as a basis for further evaluation of chemically defined GMP-grade media development.
In conclusion, we developed an erythroid cell culture medium that showed very high cell proliferation with good final erythrocyte yields. The OMM is simple to prepare by mixing two commercial media and can be widely applicable to erythropoiesis research in vitro as well as in vitro production of RBCs for various purposes.
Footnotes
Acknowledgments
The authors thank Dr. Jaehyung Yoo for providing samples. This research was supported by the Bio & Medical Technology Development Program of the National Research Foundation funded by the Korean Government, MSIP (NRF-2015M3A9C6029073).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.