
Abstract
Differentiation of stem cells as a cell-based therapy for repairing, replacing, or restoring damaged tissues such as bone, cartilage, and tendon is becoming increasingly attractive within the field of musculoskeletal tissue engineering. Toward this end, there are numerous published and well-defined protocols to differentiate stem cells toward cartilage and bone tissues, but the protocols toward tendon tissue are still emerging and thus less developed. Recent studies focused on the induction of tendon-specific markers in cultured stem cells using different growth factors (GFs), including bone morphogenetic proteins (BMPs) and transforming growth factor (TGF) isoforms. However, the inclusion of serum in relatively high concentration across these studies is less favorable, since the components within serum may interfere with the induction of the markers. Alternatively, in vitro studies with low concentration or absence of serum would be ideal. In this study, we assessed the induction effect of BMP-12 and TGF-β1 on tendon-specific markers in adipose-derived stem cells (ADSCs), in serum-free conditions. Specifically, we investigated the temporal and dosing effects of both GFs on several markers. Our results demonstrate that BMP-12 induces late expression of the transcription factors Scleraxis (SCX) and Mohawk (MKX), whereas TGF-β1 induced their earlier expression. Moreover, BMP-12 induced Decorin (DCN), but was inhibited by TGF-β1. Other markers such as collagen Iα1 (COL1A1) likewise showed this pattern. Importantly, the protein analysis generally supported the gene expression data. Interestingly, differences were observed in the cellular localization of SCX between BMP-12 and TGF-β1 stimulations. Furthermore, the addition of ascorbic acid with either BMP-12 or TGF-β1 resulted in increased deposition of collagen I. Our results enhance the existing protocols for the differentiation of ADSCs toward the tenogenic lineage in serum-free conditions and contribute to the understanding and the development of tenogenic induction protocols.
Impact Statement
Herein, we describe the tenogenic effect of bone morphogenetic protein-12 and transforming growth factor-β1 in cultured adipose-derived stem cells (ADSCs) in serum-free conditions. This culture system provides an insight into serum-free culture conditions in stem cell differentiation protocols. A positive response of the ADSCs to the tenogenic induction was observed. In particular, the different growth factors used in this study displayed notable differences both on the gene and on the protein expression of the tendon-specific markers. The results underline the positive outcome of the serum removal in tenogenic differentiation protocols, contributing to the development of future cell-based therapies for tendon regeneration and repair.
Introduction
Tendon injuries are a common clinical problem affecting millions of patients worldwide. In less severe cases, conservative treatments target clinical symptoms such as pain or inflammation. However, in severe cases such as tendon rupture, surgical interventions are needed, and this may result in prolonged rehabilitation processes. In either case, the affected tissue often fails to fully recover or regain the native tendon functions, leading to pain and inferior tissue quality. 1 Moreover, the prevalence of these injuries is likely to set to increase, particularly in the sports community or in the elderly population. 2 Therefore, alternative treatments focusing on tissue regeneration, in particular, cell-based therapies, are becoming attractive revenues for next-generation therapeutics.
Toward this end, the application of autologous adult mesenchymal stem cells for tissue regeneration is becoming increasingly popular within the musculoskeletal field. 3 This is because of the ability of stem cells to differentiate to other phenotypes of the same embryologic origin in vivo, or under the influence of physical and chemical cues in vitro.4,5 Over the past decade, numerous reports have been published on the application of stem cells in musculoskeletal research, with a particular focus on methods for describing stem cell differentiation and their in vitro characterization, with specific attention to bone and cartilage tissues. However, the methods for the differentiation of stem cells toward tendon tissue are less understood and still emerging.
Recently, several studies have focused on the induction of stem cells toward tenogenic lineage, demonstrating promising results on this front. For example, reports have shown the upregulation of common tendon-specific markers such as scleraxis (SCX) in in vitro cultured bone marrow-derived mesenchymal stem cells (BMSCs) with a range of growth factors (GFs), including bone morphogenetic proteins (BMPs) or transforming growth factor (TGF) isoforms, thus indicating the tenogenic potential of BMSCs.6,7 However, for the BMSCs to become a clinically viable source of cells, a high volume of them is required. This could pose some concerns, since obtaining BMSCs requires an invasive surgical procedure 8 and very often, the amount of cells collected in each sample is relatively low in numbers, although these can be further expanded in in vitro conditions. 9
To address these concerns, researchers have shifted the focus onto the use of adipose-derived stem cells (ADSCs) as an alternative to BMSCs. 10 ADSCs can be obtained through lipoaspirates 11 in high volumes12–15 . Preliminary studies have shown the induction of tenogenic-specific markers in cultured ADSCs stimulated with BMPs and TGF isoforms, demonstrating their differentiation potential toward a tenogenic lineage. For example, tenogenic markers, including SCX and Mohawk (MKX) were shown to be upregulated in ADSCs induced with BMP-12 and TGF-β1, and the deposition of collagen type I and several proteoglycans has been reported.6,14–17 Notably, the use of serum in the published studies is a common feature, raising concerns around the interference of the mixture of proteins and GFs in unknown quantities within serum. Moreover, the use of serum represents a limitation for the clinical applicability of the cell-based therapies to humans. Therefore, and despite the promising results reported so far, the emergence of a well-defined method for differentiation of ADSCs toward tendon tissue is still lacking. 6
Herein, in serum-free conditions, we have investigated the differentiation potential of ADSCs stimulated with BMP-12 and TGF-β1 tenogenic GFs. In particular, we expanded the study characterizing the effect of dosing and time intervals on a variety of tendon-specific markers. Parameters such as the induction of tendon markers and the deposition of a tenogenic extracellular matrix (ECM) by the cultured ADSCs were investigated and subsequently characterized by RT-qPCR, Immunostaining, and Western Blotting techniques, expanding our understanding on the protocols for the differentiation of ADSCs in serum-free conditions toward a tenogenic lineage.
Materials and Methods
Cell culture
Human ADSCs from one donor (Catalogue number PT-5006, Lot number 0000439846) were supplied by the manufacturer (Lonza), under ethical conditions after the donor consent. Cells were used in passage 3 for the experiments and grown as monolayers in ADSC basal media supplemented with 10% FBS, 1% of L-glutamine, and 0.1% of gentamicin-amphotericin as specified by the manufacturer (Catalogue number PT-4505; Lonza). During expansion, ADSCs were grown in Nunc EasYFlasks of 175 and 75 cm2 culture area (Thermo Fisher Scientific) until 80% of cell confluence was reached, refreshing 70% of the media every 4 days, to supply fresh nutrients to the cells, but preserving secreted GFs necessary for cell communication. For the cell seeding, after washing the cells in PBS, cells were detached with trypsin/EDTA solution (Lonza) for about 3 min at 37°C and subsequently neutralized with trypsin neutralization solution (Lonza). Cells were centrifuged before counting to remove residual trypsin. For the experiments, three independent cell culture replicas were used.
Study design and ADSC stimulation
Cells were left to attach for 24 h in basal media containing FBS (10%). Monolayers were then washed with PBS and starved overnight (0% FBS). BMP-12 or TGF-β1 was added (10 or 50 ng/mL) in the presence or absence of ascorbic acid (AA) (50 μg/mL) for determined time intervals. Unstimulated ADSCs grown in starved media were used as a control in all the experiments. All supplements were refreshed every 3 days. All the experiments were performed at the same hour to avoid potential circadian variation in the cell response. Schematic representation of the study design is shown in Supplementary Figure S1. Cultures were monitored and photographs were taken using a phase contrast microscope (EVOS XL Core; Thermo Fisher Scientific) with a 10 × magnification and processed in ImageJ software.
MTS assay
Cells were cultured in 96-well cell culture plates (Nunc™ Surface; Thermo Fisher Scientific) in specific densities (4000 cells/0.3 cm2) and treated with 10 or 50 ng/mL of BMP-12 or TGF-β1 in serum-free media for 1, 5, 7, and 14 days. Ten percent of MTS (Promega) was added and incubated at 37°C in the dark for 3 h. Absorbance was measured at 490 nm with a microplate reader (CLARIOSTAR; BMG Labtech). Data are presented as optical density (OD) values.
Live and dead viability assay
Cells were cultured in 24-well cell culture plates (Nunc Surface; Thermo Fisher Scientific) in specific densities (28,000 cells/1.9 cm2) and treated with 10 or 50 ng/mL of BMP-12 or TGF-β1 in serum-free media for 1, 5, 7, and 14 days. Monolayers were washed with PBS and freshly prepared Calcein AM-Ethidium homodimer-1 (Thermo Fisher) solution was added to the monolayers following manufacturer's guidelines. Imaging was performed using a Zeiss Axiovert 200M microscope using AxioVision software. Pictures were taken with a 20 × magnification Plan-Neofluar (0.5 NA) objective and a Zeiss AxioCam HRm CCD camera. Fluorescence was excited either at 494–517 nm (calcein, green) or 528–617 nm (ethidium homodimer-1, red). Merged images were created in Image J software. Live and dead cells were counted in every picture, using minimum three random locations per sample.
Real-time quantitative PCR
Cells were cultured in six-well cell culture plates (Nunc surface, Thermo Fisher Scientific) in specific densities (150,000 cells/9.6 cm2) and treated with 10 or 50 ng/mL of BMP-12 or TGF-β1 in serum-free media for 1, 5, 7, and 14 days. Monolayers were lysed with TRI Reagent solution (Thermo Fisher). Total RNA was separated into the aqueous phase with 1-bromo-3-chloropropane (Sigma), precipitated with isopropanol, and washed with ethanol at 4°C. DNA contaminations were removed using DNA-free™ DNA removal kit (Thermo Fisher) following manufacturer's guidelines. RNA purity was assessed by NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific). First-strand complementary DNA (cDNA) was synthesized using high-capacity cDNA Reverse transcription kit (Thermo Fisher) in a PTC-100 Thermal Cycler (Biorad) (10 min at 25°C, 120 min at 37°C, and 5 min at 85°C). Ten nanograms of cDNA was used in RT-qPCR reactions in a 20 μL volume, with a SYBR Green PCR Master Mix (Sigma) in a Rotor-Gene Q 2plex Platform (Qiagen) (initial denaturation 94°C for 2 min, 40 cycles of denaturation at 94°C, and annealing and extension at 60°C). Data were analyzed using the threshold cycle (Ct)—relative quantification method (2−ΔCt), between the target gene(s) and the housekeeping gene GAPDH, and expressed as a relative expression. Primers used for RT-qPCR experiments were obtained from published literature; their specificity was confirmed using NCBI Primer Blast and purchased from Thermo Fisher Scientific (Supplementary Table S1).
Immunocytochemical staining
Cells were cultured on glass coverslips in 24-well cell culture plates (Nunc Surface; Thermo Fisher Scientific) in specific densities (28,000 cells/1.9 cm2) and treated with 10 or 50 ng/mL of BMP-12 or TGF-β1 with or without AA in serum-free media for 1 and 7 days. Monolayers were fixed in 4% paraformaldehyde for 10 min and washed with PBS. For C1A1 and SCX intracellular localization, cell membranes were permeabilized for 3 min with 0.5% NP-40 (Thermo Fisher) and blocked with 3% BSA in PBS for 1 h at room temperature. For non-permeabilized samples, cell membranes were directly blocked. Coverslips were then incubated overnight at 4°C in a humidified chamber with anti-SCXA (1:100, Abcam; ab58655, Rabbit Polyclonal) or anti-C1A1 (1:100, Abcam; ab34710, Rabbit Polyclonal). Monolayers were then incubated with 3% BSA in PBS for 10 min and subsequently incubated with Goat anti-Rabbit IgG (H+L) (1:400; Invitrogen, A-11034) for 1 h in the dark. Coverslips were then washed with PBS and mounted with mounting media containing DAPI (Vector Laboratories; H-1500). Imaging was performed using a Zeiss Axioplan 2ie microscope using AxioVision software. Pictures were taken with a 20 × magnification Plan Apochromat (0.6 NA) objective and a Zeiss AxioCam HRm CCD camera. Fluorescence was excited either at 300–400 nm (DAPI) or 500–560 nm (AlexaFluor 488). Merged images of DAPI (blue) and collagen (green) or SCX (red) were created and fluorescence was quantified in Image J software. Data are expressed as mean of the fluorescent intensity, using minimum three random locations per sample.
Western blotting
Cells were cultured in 60 × 15 mm dishes (Nunc Surface, Thermo Fisher Scientific) in specific densities (300,000 cells/21.5 cm2) and treated with 10 ng/mL of BMP-12 or TGF-β1 in serum-free media for 1 and 7 days. Media were removed from monolayers, washed three times with chilled PBS, and lysed for 5 min at 4°C with RIPA lysis buffer (Thermo Fisher), freshly supplemented with phenylmethanesulfonyl fluoride (Sigma Aldrich) and protease inhibitor cocktail (Sigma Aldrich). Cell lysates were collected and centrifuged for 30 min at 4°C. Supernatants were quantified for protein concentration using Pierce BCA protein assay (Thermo Fisher) following manufacturer's instructions. For collagen 1A1 and SCX detection, 5 μg of cell lysates was separated in 8% (120V, 70 min) or 12% (200V, 50 min) SDS-PAGE gels, respectively, under reducing conditions. Gels were wet transferred onto a 0.45 μM pore size PVDF membrane (GE Health care) for 18 h at 15V for collagen 1A1 or for 2 h at 90V for SCX at 4°C. Membranes were blocked with milk protein (5%) in TBS-T buffer for 1 h and then incubated with anti-C1A1 (1:1000, Abcam; ab34710, Rabbit Polyclonal), anti-SCXA (1:250, Abcam; ab185940, Rabbit Polyclonal), or anti-GAPDH (1:1000, R&D Systems; AF5718, Goat Polyclonal) overnight at 4°C. For the secondary antibody incubation, membranes were incubated with anti-goat for GAPDH (1:1000; R&D systems) or anti-rabbit for collagen 1A1 (Collagen type I) and SCX (both dilution 1:1000) (Abcam), conjugated with horseradish peroxidase. Membranes were analyzed using ECL plus detection system (Amersham Biosciences) using ImageQuant LAS 4000 platform (GE Health care).
Statistical analysis
Two-way ANOVA statistical analysis was used with two main variables, dose of GFs and time of stimulation. In RT-qPCR data, Tukey's multiple comparison tests were applied for displaying significant differences of the doses in between the time points and Dunnet's multiple comparison tests were applied for displaying significant differences of the doses compared to unstimulated cells. In MTS, OD values are expressed as mean ± SD (n = 3 independent cell culture replicates). In Live and Dead assay, counted live and dead cells per picture are expressed as mean ± SD (n = 3 independent cell culture replicates). In RT-qPCR, fold change compared to control data is expressed as mean ± SD (n = 3 independent cell culture replicates). In immunocytochemical (ICC), for permeabilized ADSCs, data are expressed as mean of fluorescence intensity (FI) ± SD (n = 3 independent cell culture replicates) and normalized to cell number. For nonpermeabilized cells, FI without cell number normalization is reported as mean of FI ± SD (n = 3 independent cell culture replicates).
Results
Effect of serum-free conditions and tenogenic media in the viability of ADSCs
Due to the serum-free conditions, morphology and viability of the cultured ADSCs were monitored. In serum-free conditions, ADSCs presented a star-shaped morphology with a less elongated cytoplasm, compared to standard culture conditions containing serum (Fig. 1A). Tenogenic media containing BMP-12 did not reverse the observed morphology (Supplementary Fig. S2), but the addition of TGF-β1 generated a more elongated cytoplasm (Supplementary Fig. S3). Interestingly, no significant differences were found in the metabolic activity (Fig. 1B), although it was found to be slightly higher and constant with TGF-β1. To further confirm the viability of the cultures, we then stained and quantified the live and dead cells (Fig. 1C–E). BMP-12 stimulation did not change the ADSC morphology during the time-course experiment (Fig. 1C), and both the number of live and dead cells remained stable with no significant variations (Fig. 1E). In contrast, we detected enlarged cells in the presence of TGF-β1, which became visible 5 days after the induction (Fig. 1D). However, no significant increase in the number of live cells was detected (Fig. 1E).

Viability of ADSCs in serum-free media and stimulated with BMP-12 or TGF-β1. Cell morphology and general status of cultured ADSCs were monitored by bright field.
BMP-12 tenogenic media moderately induce the expression of tenogenic markers in ADSCs
To detect changes in the gene expression, RT-qPCR was used in ADSCs cultured in serum-free conditions with BMP-12 refreshed every 3 days (Fig. 2). The analysis revealed a late and mild effect of BMP-12 on the expression of almost every tendon marker. Collagen 1α1 (COL1A1), collagen 1α2 (COL1A2), collagen 3α1 (COL3A1), tenascin C (TNC), decorin (DCN), MKX, thrombospondin-4 (THB-4), and cartilage oligomeric matrix protein (COMP) mRNA levels were upregulated at the end of the stimulation period (14 days), being barely detectable beforehand. Among those, DCN, MKX, THB-4, and especially, COMP were significantly induced. Interestingly, the latter displayed the highest mRNA levels by BMP-12 addition, in particular with the highest dose administrated (50 ng/mL). On the other hand, variable SCX expression was observed over the time course, being considerably upregulated 7 and 14 days after induction.

Characterization of tenogenic induction in ADSCs stimulated with BMP-12. ADSCs were cultured in serum-free media for 1, 5, 7, and 14 days and stimulated with 10 or 50 ng/mL of BMP-12. Unstimulated ADSCs were used as a control. mRNA levels were analyzed by q-RT PCR. The dashed line represents baseline equal to 1, calculated with DCt values from unstimulated ADSCs at each time point. Data are expressed as mean of fold change ± SD (n = 3 independent cell culture replicates), compared to control (unstimulated ADSCs). Statistical analysis two-way ANOVA with Tukey's multiple comparison test was applied for displaying significant differences of the doses in between the time points and Dunnet's multiple comparison test was applied for displaying significant differences of the doses compared to unstimulated cells. **p < 0.002, ***p < 0.0002, ****p < 0.0001. COL1A1, collagen 1α1; COL1A2, collagen 1α2: COL3A1, collagen 3α1; SCX, scleraxis; MKX, Mohawk; THB-4, thrombospondin–4; DCN, decorin; TNC, tenascin C; COMP, cartilage oligomeric matrix protein; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
TGF-β1 tenogenic media highly induce the expression of tenogenic markers in ADSCs
Contrary to the observed induction by BMP-12 (Fig. 2), TGF-β1 showed an earlier upregulation in all the tenogenic markers, except for DCN (Fig. 3). Five days after the induction, tendon markers such as COL1A1 and MKX were significantly upregulated and observed to decrease thereafter. However, the expression was maintained for over 14 days of induction in the case of COL1A1 and barely detected in the case of MKX. The mRNA levels of other markers such as THB-4, TNC, and COL3A1 were upregulated and maintained since the beginning of the induction, increasing thereafter. Moreover, TGF-β1 showed significant upregulation and maintenance of the markers SCX and COMP throughout all the time-course experiments. In the case of SCX, high mRNA levels were detected from the first time interval. In the case of COMP, a large increase in expression was observed. Interestingly, none of the tested conditions induced the expression of DCN, which was continuously downregulated.
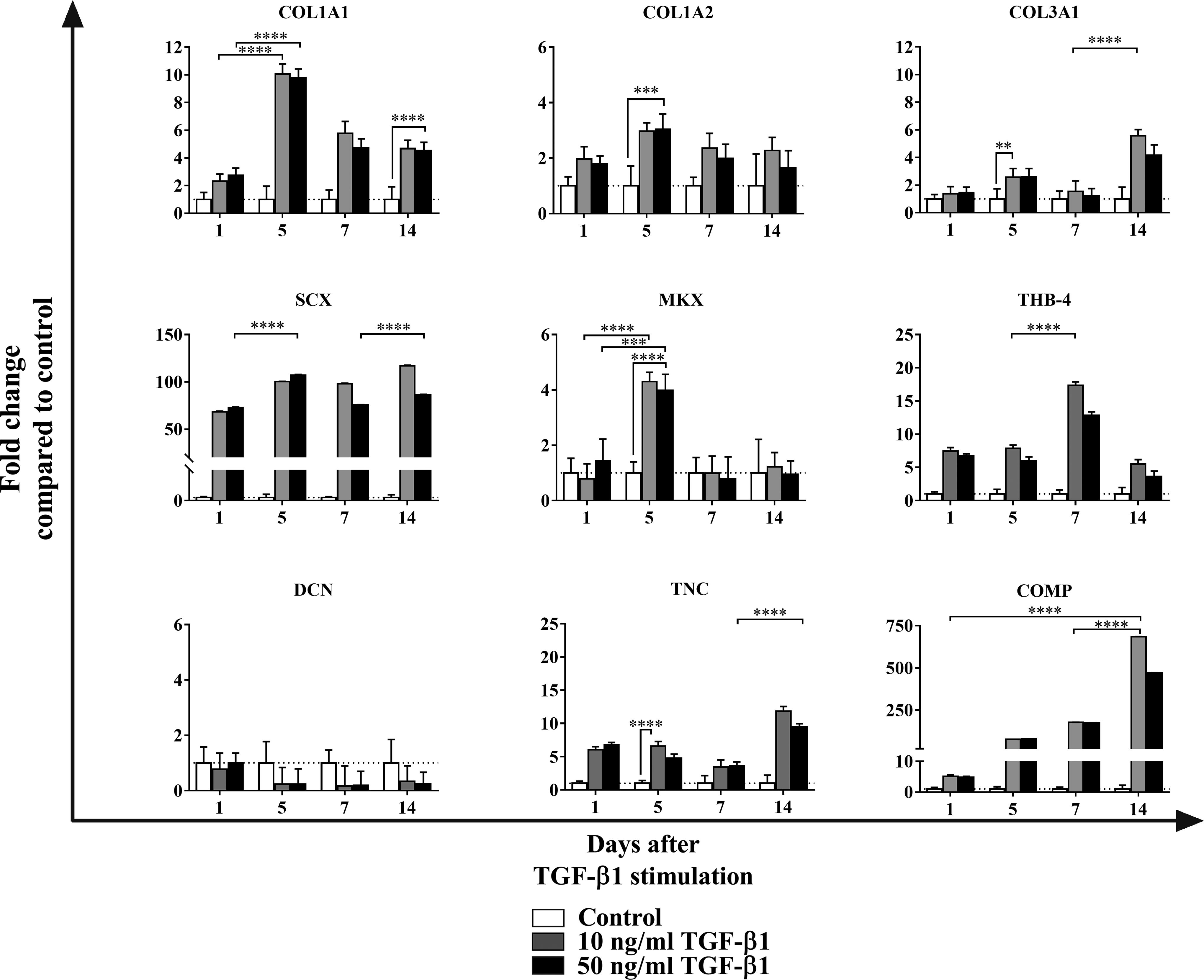
Characterization of tenogenic induction in ADSCs stimulated with TGF-β1. ADSCs were cultured in serum-free media for 1, 5, 7, and 14 days and stimulated with 10 or 50 ng/mL of TGF-β1. Unstimulated ADSCs were used as a control. mRNA levels were analyzed by qRT-PCR. The dashed line represents baseline equal to 1, calculated with DCt values from unstimulated ADSCs at each time point. Data are expressed as mean of fold change ± SD (n = 3 independent cell culture replicates), compared to control (unstimulated ADSCs). Statistical analysis two-way ANOVA with Tukey's multiple comparison test was applied for displaying significant differences of the doses in between the time points and Dunnet's multiple comparison test was applied for displaying significant differences of the doses compared to unstimulated cells. **p < 0.002, ***p < 0.0002, ****p < 0.0001. COL1A1, collagen 1α1; COL1A2, collagen 1α2; COL3A1, collagen 3α1; SCX, scleraxis; MKX, Mohawk; THB-4, thrombospondin-4; DCN, decorin; TNC, tenascin C; COMP, cartilage oligomeric matrix protein; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Media containing BMP-12 and TGF-β1 mildly induce the expression of cartilaginous markers in ADSCs, while having little or no effect on the expression of the osteogenic marker
The cartilage marker aggrecan (ACAN) showed to be significantly upregulated by BMP-12 at a later time of induction (Fig. 4A), whereas TGF-β1 had almost no effect in the upregulation of this marker, only weakly expressed 14 days after the induction (Fig. 4B). In addition, sex-determining region Y-box 9 (SOX9) was selected as another marker indicating a cartilaginous differentiation. In contrast to what was detected with ACAN, we observed that SOX9 was significantly upregulated by TGF-β1 since the first time point of induction. However, SOX9 expression significantly decreased during the time-course experiment, showing mRNA levels closer to basal at the end of the stimulation time (Fig. 4B). On the other hand, BMP-12 did not induce SOX9 expression (Fig. 4A). The expression of the osteogenic marker osteocalcin (BGLAP) was also analyzed. Neither of the tested tenogenic media, containing BMP-12 or TGF-β1 (Fig. 4A, B, respectively), increased the expression of BGLAP compared to the control.
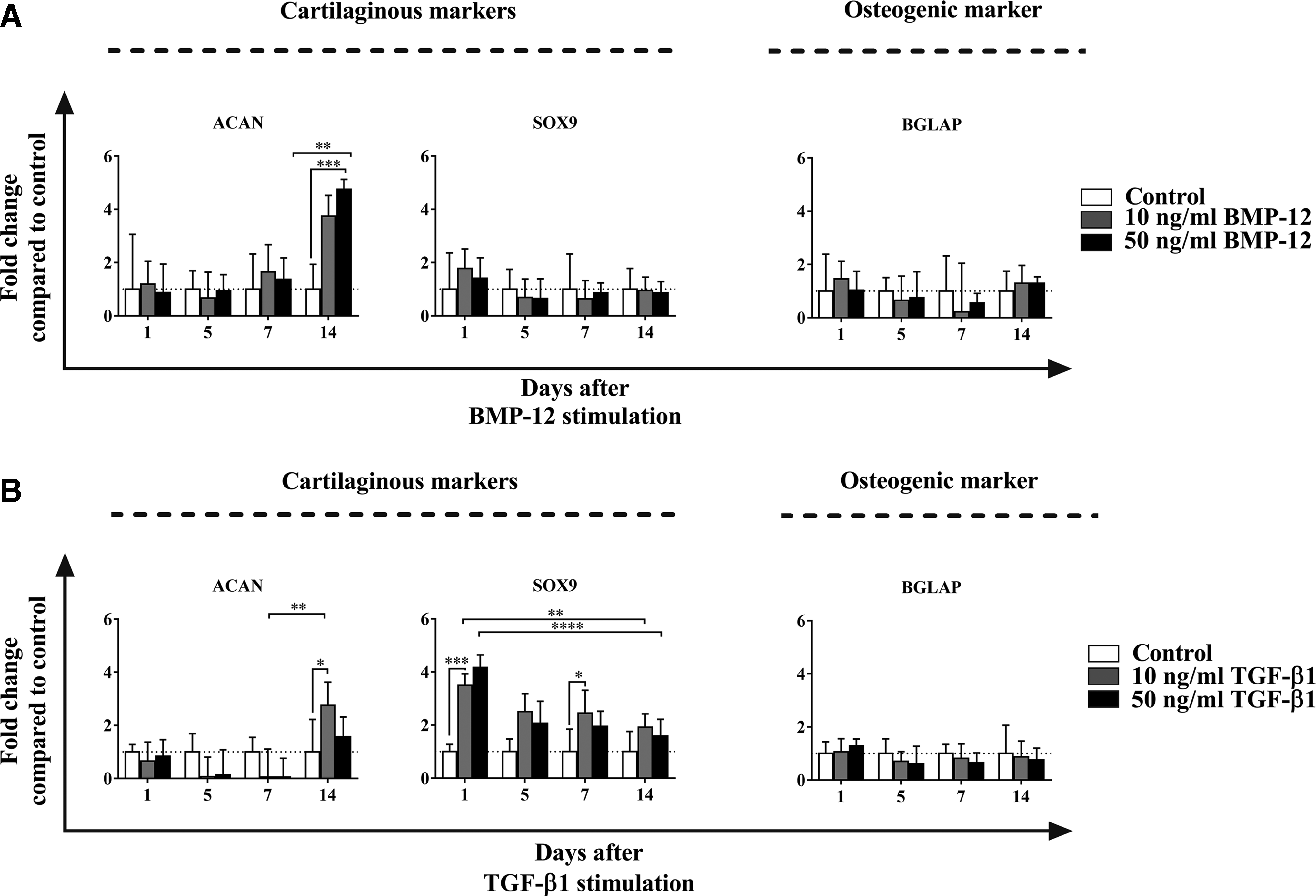
Characterization of cartilaginous and osteogenic induction in ADSCs stimulated with BMP-12 or TGF-β1. ADSCs were cultured in serum-free media for 1, 5, 7, and 14 days and stimulated with
Effect of tenogenic media and AA on collagen type I protein expression in permeabilized ADSCs
One and 7 days after the tenogenic induction with BMP-12 or TGF-β1, localization of collagen type I (COL1) was assessed and quantified by immunocytochemical techniques (ICC) (Fig. 5A, B) in permeabilized ADSCs and protein levels were confirmed by Western Blotting (Fig. 5C). Moreover, AA was added to the tenogenic media and the effect on COL1 fibril formation was observed. Overall, COL1 was detected in the presence of all the basal and tenogenic media (Fig. 5A, B) and was visible in the form of a cytoplasmic distribution. However, the addition of AA allowed for the formation of collagen fibrils, secreted into the extracellular space between cells. We observed that the few fibrils generated by the combination of BMP-12 and AA (Fig. 5A) were mostly detectable 7 days after the tenogenic induction. In contrast, the mixture of TGF-β1 and AA (Fig. 5B) strongly induced the accumulation of the fibrils from the first day of induction, maintained over the time course. These results were confirmed by quantification of the fluorescent intensity (Fig. 5B). Furthermore, analysis by Western blotting (Fig. 5C) showed COL1 protein levels were detected in basal and tenogenic media, but strongly upregulated by TGF-β1, especially at early time points. Seven days after the stimulation, we detected a decrease in the intensity of the signal both in basal- and BMP-12-containing media. At the same time, TGF-β1 stimulation increased COL1 protein levels compared to BMP-12, displaying the most intense signal from all the time point (Fig. 5C). Full-length version of the gels is available in supplementary section (Supplementary Fig. S4).
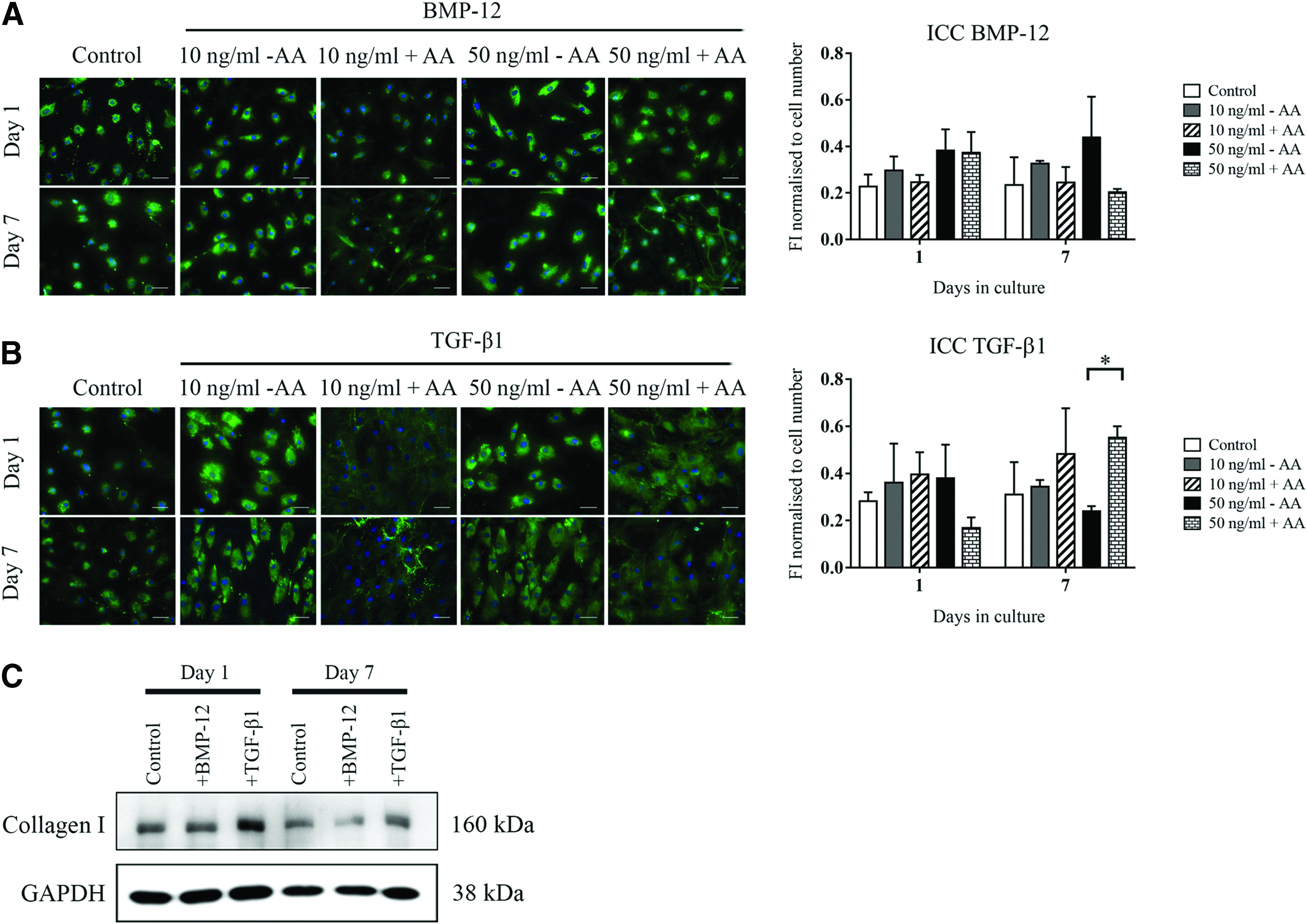
Collagen type I protein immunolocalization in permeabilized ADSCs stimulated with BMP-12 or TGF-β1. Collagen 1 protein was detected by immunocytochemical techniques in permeabilized ADSCs stimulated with
Effect of tenogenic media and AA on collagen type I deposition in nonpermeabilized ADSCs
Following the results seen in Figure 5, we then next assessed the collagen I localization in nonpermeabilized ADSCs. We observed both basal and BMP-12-containing media were not able to induce large quantities of deposited collagen, and mostly without a defined arrangement (Fig. 6A). Few fibrils were detected in the presence of AA, although only in a significant manner at early time points, as confirmed by the quantification analysis. The mixture of TGF-β1 and AA also generated collagen fibril deposition without a defined orientation (Fig. 6B), however, in larger quantities, significantly increased on day 7, compared to the previous time point.
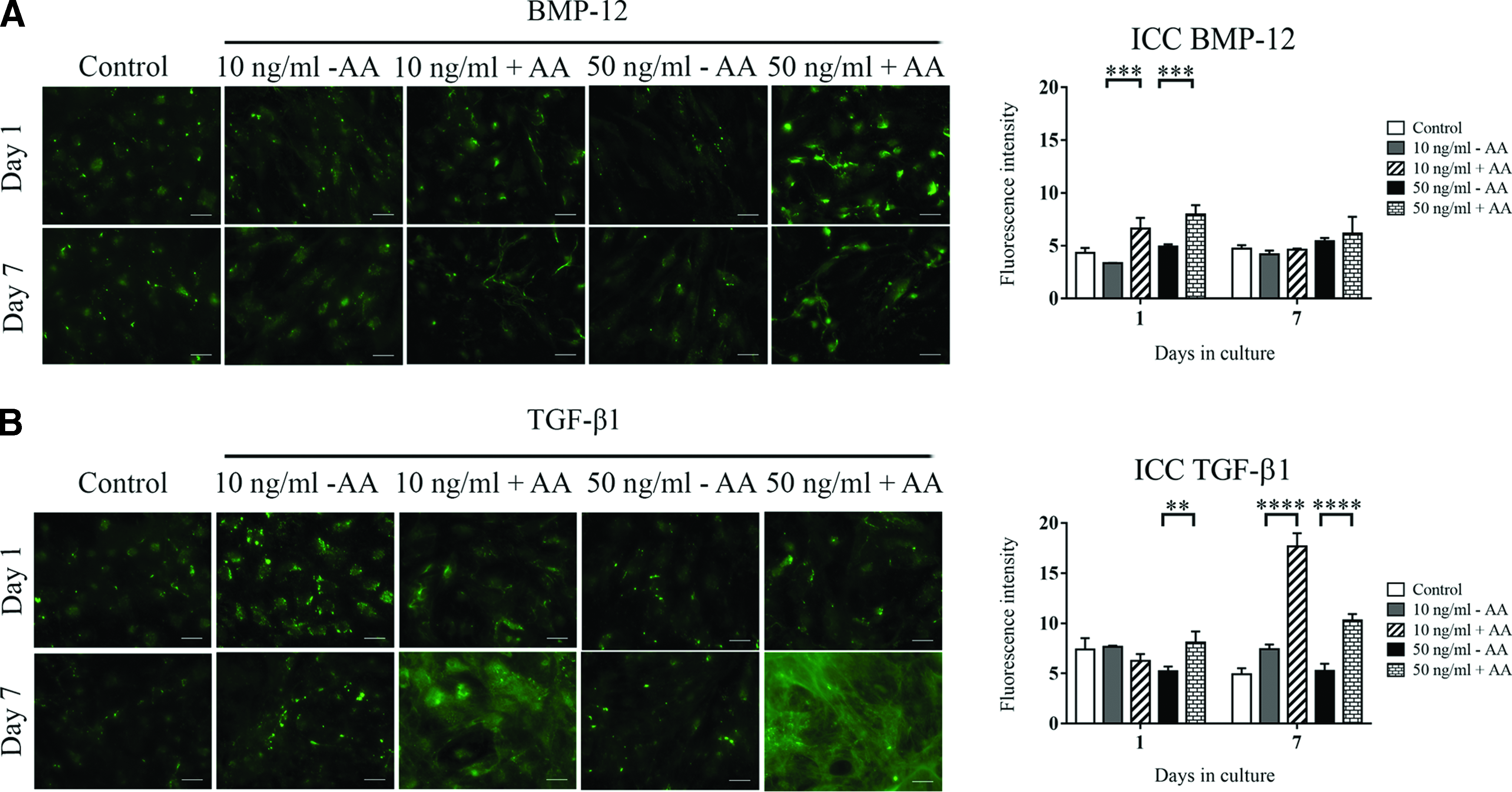
Collagen type I protein immunolocalization in nonpermeabilized ADSCs stimulated with BMP-12 or TGF-β1. ADSCs were cultured in serum-free media for 1 and 7 days and stimulated with
Evaluation of SCX expression as an effect of the administered tenogenic media in ADSCs
The presence of SCX in cultured ADSCs was assessed next by ICC and protein levels confirmed by Western Blotting (Fig. 7). Overall, we detected differences in the distribution of SCX depending on the tenogenic media used. BMP-12-containing media displayed a positive SCX signal in early induction time points, with levels closer to basal. SCX displayed a subcellular localization, with strong staining in the cell nuclei. However, the longer induction time resulted in a complete loss of the SCX signal, barely above the background, as confirmed by the quantification of the FI (Fig. 7A). On the other hand, SCX was detected both in the cell nuclei and in the cytoplasm of ADSCs induced with TGF-β1, regardless of the dose or time point (Fig. 7B). In addition, the intensity of the signal was higher than both basal and BMP-12-containing media (Fig. 7A), remaining constant in late induction times (Fig. 7B). Consistent with the ICC results, protein levels analyzed by Western blotting showed a decrease in SCX protein 7 days after the induction, only being maintained and upregulated in tenogenic media containing TGF-β1 (Fig. 7C). Full-length version of the gels is available in Supplementary Figure S5.

Scleraxis protein expression in ADSCs stimulated with BMP-12 or TGF-β1. Scleraxis protein was detected by immunocytochemical techniques in ADSCs stimulated with
Discussion
In this study, we report a comprehensive analysis of tenogenic differentiation induced by BMP-12 and TGF-β1 in cultured ADSCs, varying doses as well as different time intervals. Most importantly, the study was conducted in serum-free conditions to eradicate the endogenous effect of the serum.
The culture conditions did not cause ADSCs cytotoxicity, although a morphology change was observed. Instead of the well-described fibroblast morphology, 10 ADSCs displayed a smooth-edged configuration. This shift was not reversible upon addition of BMP-12, but effectively reversed by TGF-β1, in agreement with previous studies performed with low serum concentrations. 18
Overall, the results revealed a delayed effect of BMP-12 in the induction of the tenogenic markers. In contrast, the addition of TGF-β1 demonstrated a predominant role not only in the early activation of the markers but also in the maintenance of their expression across the time intervals, both at gene and protein level. The tenogenic media containing TGF-β1 rapidly activated SCX and MKX, two transcription factors that have key mediator roles in tendon development.6,19 Although MKX was found to be significantly upregulated 5 days after the induction, SCX expression was strongly upregulated throughout all the time intervals of TGF-β1 induction. On the contrary, it scarcely achieved significance in tenogenic media containing BMP-12. Moreover, the contained nuclear distribution of SCX upon addition of BMP-12 was observed both in the nuclei and in the cytoplasm of the ADSCs after addition of TGF-β1, indicating a possible effect of TGF-β1 in the accumulation of SCX. However, the morphology of the cultured ADSCs in tenogenic media containing TGF-β1 was notably enlarged compared to the rest of the conditions; therefore, the accumulation of SCX in the cytoplasm could be related to the changes in cell morphology and should be addressed further. Nonetheless, these results suggest that TGF-β1 is more effective at activating and preserving SCX expression, in agreement with other studies.19–21
The results further revealed a potential inhibitory effect of TGF-β1 in DCN expression, 22 an ECM component with a role in collagen fibril formation.23,24 In contrast, BMP-12 significantly upregulated DCN 2 weeks after the first induction. These results are in agreement with previous studies. 6
AA is a well-known co-factor for the correct assembly of the collagen triple helix. 25 Therefore, the investigation of the role of AA as a supplement in stem cell differentiation media is highly desirable. Initially, we carried out a preliminary study assessing the effect of AA in RT q-PCR, ICC, and Western blotting experiments. However, the addition of AA did not display significant differences at mRNA or protein level, compared to cultures without AA (data not shown). Although the positive effect of AA at mRNA and protein level has been described previously, 26 we then considered that the effect of AA in the collagen deposition is apparent in the ICC experiments, and thus, the inclusion of AA is exclusively in ICC in this study. Collagen fibers in the extracellular space are apparent by immunolocalization upon addition of AA. Future work, including AA in all experimental designs, especially Western blotting, should be considered and investigated. Overall, upon stimulation with TGF-β1 and BMP-12, a cytoplasmic distribution of Collagen was observed, suggesting a procollagen synthesis before the extracellular secretion, 27 independent of the GF. However, the addition of AA clearly showed the extracellular and intracellular accumulation of fibrils, especially in combination with TGF-β1, confirming the positive role of this co-factor in collagen biosynthesis and deposition,26,28 and the beneficial combination of AA and TGF-β1 in collagen synthesis, as described previously. 26 Further contributing to collagen fibrillogenesis, COMP is a glycoprotein that is believed to provide integrity to the tendon ECM. 7 Interestingly, all tenogenic media displayed an exponential increase of COMP expression. These observations could suggest a potential and gradual effect of these GFs on the accumulation of COMP to the ECM.
The obtained results underline the positive response of the ADSCs to the GF addition in the absence of serum. The selected tendon-specific markers were overall upregulated and both BMP-12 and TGF-β1 revealed to have mild induction in cartilage and no effect on bone markers. Nonetheless, the chosen bone marker (osteocalcin) is considered a marker of a late osteogenic development, and further analysis with other earlier osteogenic markers such as runt-related transcription factor 2 (RUNX2) should be carried further in these conditions, although other examples can be found in the literature.14,15
Moreover, the experimental design of this study agrees with previously reported studies, since the continuous administration of GFs is a commonly used approach.14,15,29 However, recent research explores the stepwise addition of GFs, 6,17 priming the cultured cells with one or several GFs before the stimulation. The differences in the effect of the mentioned approaches on the cultured cells address the necessity of consolidation of tenogenic protocols. In addition, the use of no serum or low serum concentrations in stem cell characterization studies should be explored further. Moreover, other experimental setups during crucial steps for stem cell differentiation protocols such as the initial stem cell expansion could be addressed further. In this study, the ADSCs were expanded in 10% FBS, and recent literature describes the use of low serum maintenance medium as a beneficial alternative 30 for the later applicability of the stem cells.
Therefore, the significance of this study resides in the utilization of multiple doses and time intervals of two important tenogenic GFs in serum-free conditions. The overall positive response of the ADSCs, the absence of cell cytotoxicity, and the differences found in the tenogenic profile reinforce the potential exclusion of serum in tenogenic differentiation protocols, although deeper studies should be conducted. In addition, the positive assets of ADSCs venture them as an important stem cell population for regenerative medicine and, in particular, for tendon regeneration.
We have demonstrated a strong upregulation and maintenance of the tenogenic commitment upon addition of TGF-β1, not only inducing early key tendon markers such as SCX but also promoting the deposition of a tendon-like ECM, especially in combination with AA. Despite extensive studies probing the effect of BMP-12 in different cell types15,29,31 we found it ineffective to actively induce tendon commitment in ADSCs. Even so, we address the importance of longer time intervals to further test the effect of BMP-12.
Ultimately, this study emphasizes the relevance of cell culture microenvironments for the development of more accurate and representative tenogenic differentiation tools for pluripotent stem cells, to develop efficient future cell-based therapies for tendon repair.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.