
Abstract
Current treatments for cardiovascular diseases use biological implants to replace or repair nonfunctional tissue. Besides a low immunogenicity, those matrices should induce balanced immune responses enabling proper tissue regeneration. Polymorphonuclear leukocytes (PMNs) represent the first cellular players of an immune response, initiating subsequent attraction and activation of other immune cells. However, so far, robust in vitro assays are missing that predict PMN responses to diverse implant matrices and help to understand immune response mechanism. Therefore, we aimed to establish an easy and standardized in vitro assay to assess matrix-induced PMN responses. First, PMNs were isolated from blood of healthy donors and stimulated with various cytokines, chemokines as well as lipopolysaccharide (LPS) and the peptide N-formyl-met-leu-phe (fMLP). To select appropriate activation conditions and markers, surface expression of CD11b, CD16b, CD35, CD43, CD62L, CD63, and CD66b on PMNs was determined by flow cytometry after 4 h in culture. In addition, the cytokine secretion of interleukin (IL)-6, MIP-1β, and IL-8 was measured by enzyme-linked immunosorbent assays (ELISA). Second, PMNs were cultured on “proof-of-principle” matrices: (1) aortic tissue after conventional frozen cryopreservation or ice-free cryopreservation and (2) decellularized amniotic membrane (DeAM), additionally coated with a cardiac extracellular matrix hydrogel (DeAM+E), and analyzed for PMN activation. The activation assay validated LPS as most effective and dose-dependent inducer of PMN activation in vitro. The increase of CD63 and CD66b expression and decrease of CD16b, CD43, and CD62L on PMNs, as well as enhanced IL-6 secretion, were determined as reliable PMN activation markers. Both cryopreserved matrices did not trigger any PMN activation. In contrast, DeAM+E, but not DeAM alone, induced a similar expression level for CD43, CD62L, and CD63 on PMNs as the LPS activation control, but no increase of IL-6 secretion. In this study, we developed an easy, fast, and suitable assay to measure the PMN activation by biological matrices in vitro through a characteristic activation marker and cytokine profile to reveal mechanism of their potential immunogenicity and compatibility when used as implants. The established PMN assay could be a powerful additional tool to existing in vitro tests and might be implemented in the evaluation for future clinical implants.
Impact Statement
Polymorphonuclear leukocytes (PMNs) are essential in the first infection and host-versus-graft reactions. Strategies for adequate and standardized assays to test PMN activation by diverse types of matrices such as cardiovascular implants are urgently needed. To overcome this limitation, we established a straightforward PMN activation assay and validated lipopolysaccharide (LPS) as a reliable PMN activator that induces defined changes in surface marker expression and cytokine release. Biological “proof-of-principle” matrices demonstrated the feasibility of this PMN assay. Overall, this assay provides an instrument conducting an initial immunological assessment of biological implants prior their clinical application.
Introduction
Globally, cardiovascular diseases (CVDs) such as myocardial infarction or atherosclerosis are still the leading cause of death. 1 To repair or replace nonfunctional cardiovascular structures, application of synthetic biomaterials and injection of processed cardiac extracellular matrix (ECM) are promising strategies for local treatments.2–4 In the case of damaged heart valves or vessels, often the whole structure is replaced by synthetic prostheses, xenografts or allografts.
Upon biomaterial implantation, an immunological reaction is induced, resulting in either inflammation or regeneration. 5 The usage of inert biomaterials counteracts inflammatory responses but could consequently lead to encapsulation processes. 6 Therefore, desired immune effects, which are beneficial for healing and regeneration, require defined biomaterial features. 5 A balanced immune response would be fast, specific, and self-limiting. 7 As part of the innate immune reaction, the acute inflammatory response occurs within hours after implantation. Besides the activation of the complement and coagulation cascade, it is characterized by polymorphonuclear leukocytes (PMNs), predominantly neutrophils, which are the most abundant circulating leukocytes.5,8 Attracted by chemokines, PMNs from the blood infiltrate to the implant site. 9
Activated PMNs have a high capacity to phagocytose, release neutrophil extracellular traps (NETs) through NETosis, and secrete a number of factors such as chemokines, cytokines, proteases, and reactive oxygen species to mobilize and orchestrate more immune cells, mainly monocytes, to the site of inflammation.10–15 In vitro and in vivo studies suggest that the type of PMN activation can be crucial for later modulation of the immune response by macrophages and T cells.16–18 Consequently, the initial PMN-to-implant reaction is able to shape the long-term immune response either leading to chronic tissue damage, calcification and fibrosis, or rather promoting healing and regeneration.
The majority of studies evaluating the immunogenicity of implanted material investigated the behavior of macrophages or T cells upon contact.19,20 However, the performance of PMNs and their role for immunological compatibility are often neglected. So far, only a few investigations regarding PMN response toward biomaterials were conducted.11,21
Just recently, it was proposed that PMNs and their heterogeneity are an essential feature of immune pathophysiology and require more in-depth investigations. 22 Therefore, also a deeper understanding on PMN activation upon direct biomaterial contact is needed to precisely evaluate the outcome of new implant strategies and, if applicable, attempt to alter host immune reaction by specific parameter adjustments of the biomaterial. Thus, we aimed to establish a reliable and straightforward in vitro assay to evaluate the PMN reaction upon contact with biological matrices. The assay should help to deeply understand the events of an immune response to a particular implant and anticipate its compatibility and immunogenicity.
In this comparative study, we validated lipopolysaccharide (LPS) as the most potent and reliable PMN activator in vitro. We tested the suggested alteration of PMN surface marker expression during activation and identified CD16, CD43, CD62L, CD63, and CD66b as appropriate markers.
Moreover, we assessed the applicability of the established assay procedure to biological scaffolds. Therefore, PMNs were cultured with two biological “proof-of-principle” matrices: (1) cellular human aortic tissue after conventional frozen cryopreservation (CFC) or ice-free cryopreservation (IFC) and (2) decellularized amniotic membrane (DeAM) with and without coating with human cardiac ECM hydrogel (hcECM-HG). Both tissue types have the potential to restore cardiac function or improve the endogenous regenerative capacity.
CFC is commonly used to preserve cardiovascular allografts, however, the durability is limited due to the induction of inflammatory processes, leading to calcification and finally deterioration and damage of tissue structures. 23 Alternatively, IFC has been demonstrated to preserve cardiovascular matrix structures,24,25 by reduction of human immune responses in vitro26–28 and immune cell infiltration in vivo.29,30 However, the initial reaction of PMNs triggered by this new cryopreservation strategy is not characterized yet.
To treat lesions after myocardial infarction, amniotic membrane isolated from human placenta can be used as a regenerative cell-free epicardial patch. Recently, it was shown in vivo that DeAM possesses a higher potential to restore cardiac function upon infarction than the cell-containing membrane, even after forced induction of epithelial-to-mesenchymal transition. 31 Furthermore, hcECM-HG was developed and revealed preserved cytoprotective effects in vitro.32,33 The coating of the DeAM with hcECM-HG (DeAM+E) represents a very promising strategy to support cardiac regeneration from the activated epicardium.34,35 Moreover, it was demonstrated that proinflammatory cytokine secretion by cardiac fibroblasts cultured on DeAM was absent 31 and that the immune response by monocytes cultured on DeAM+E was attenuated. 34
However, only little is known about the initial immune cell activation. Thus, evaluation of PMN activation upon contact with DeAM with or without hcECM-HG coating would provide deeper insights into the immunogenicity of that biological matrix.
The test system we present in this study addresses the first cellular immune reaction to a given biological matrix in vitro. Moreover, we demonstrated the suitability and easy performance of the assay for two types of therapeutically relevant biological matrices for CVDs.
Materials and Methods
PMN isolation and purification
PMNs were isolated from healthy volunteers with written informed consent approved by the ethics committee of the Charité–Universitätsmedizin Berlin (EA2/139/10) using Polymorphprep density gradient (Alere Technologies AS, Oslo, Norway) according to manufacturer's protocol. Polymorphprep was covered with the same volume of blood and centrifuged at 480 g for 35 min at room temperature (RT) without brake. Plasma and upper band containing peripheral blood mononuclear cells were removed and lower band containing PMNs was collected and washed first with ddH2O half diluted phosphate-buffered saline without calcium and magnesium (PBS; Life Technologies/Thermo Fisher Scientific, Waltham, MA) and second with PBS (400 g, 10 min, RT). Finally, cells were resuspended in complete RPMI medium (very low endotoxin Roswell Park Memorial Institute 1640 culture medium (VLE-RPMI; Biochrom/Merck, Darmstadt, Germany) supplemented with 10% human serum from male AB plasma (Sigma-Aldrich, St. Louis, MO), 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM
Preparation of biological matrices
Human aortic tissue was obtained from patients undergoing replacement of the ascending aorta at German Heart Center Berlin (Germany), and human myocardial samples were obtained from end-stage cardiomyopathy patients who underwent a cardiac transplantation according to the ethics committee of Charité–Universitätsmedizin Berlin (EA4/028/12). All patients gave written informed consent. Human full-term placentas were anonymously obtained after cesarean delivery of healthy and uncomplicated pregnancies from women who had given written informed consent for use of the placenta for research purposes.
As described previously, aortic tissue was washed, treated with antibiotics, and frozen either according to the protocol of CFC or IFC 26 and patches composed of DeAM coated with hcECM-HG (DeAM+E) were prepared. 34
PMN standard culture conditions and in vitro activation
In a 48-well plate (Cellstar; Greiner Bio-One, Kremsmünster, Austria), 5 × 105 human PMNs (5 different donors: 2 female, 3 male, age: 20–40 years) were seeded in 400 μL complete RPMI medium and cultured in a humidified atmosphere with 5% CO2 at 37°C. For stimulation, medium was supplemented with 10 ng/mL interferon γ (INF-γ; Preprotech, Hamburg, Germany), 100 ng/mL interleukin 15 (IL-15; Preprotech), 500 ng/mL lipopolysaccharide (LPS; Sigma), 10 ng/mL tumor necrosis factor α (TNF-α; Miltenyi Biotec, Bergisch Gladbach, Germany), 40 ng/mL N-formyl-met-leu-phe (fMLP; Merck), or 1 ng/mL granulocyte-macrophage colony-stimulating factor (GM-CSF; Miltenyi Biotec), respectively. As control, 5 × 105 cells were cultured without simulation. After 4 h of culture with or without stimulation (t4), PMNs were collected and analyzed for surface marker expression or cytokine secretion. Surface marker expression of all tested markers at t4 was normalized to marker expression on PMNs at t0. Additional experiments were performed using various LPS concentrations ranging from 31.3 to 200 ng/mL LPS (three different donors: one female, two male, age: 20–40 years).
PMN culture on matrices with biological matrices as “proof-of-principle” implants
Biological “proof-of-principle” matrices (CFC and IFC aortic tissue or DeAM and DeAM+E) were placed to the bottom of the 48-well plate and fixed with a ring cut from a silicone tube (Ismatec/Cole-Parmer, Wertheim, Germany). For PMN culture on matrices, 2 × 105 PMNs (two different male donors, age: 20–30 years) were seeded directly onto biological matrices in 400 μL complete RPMI medium and cultured on the scaffolds for 4 h (t4). PMNs were collected and analyzed. Again, surface marker expression of all tested markers at t4 was normalized to marker expression on PMNs at t0.
Cytokine measurement
Culture supernatants were collected and analyzed by ELISA for presence of cytokines using human IL-6 and IL-8 ELISA MAX™ Deluxe Kit (BioLegend, San Diego, CA) and human CCL4 (MIP-1β) ELISA Kit (eBioscience/Thermo Fisher Scientific). Secretion of TNF-α, IL-10, IL-15, and PMN-Elastase was determined by using Human TNF-α ELISA MAX Deluxe, Human IL-10 ELISA MAX Deluxe, Human IL-15 ELISA MAX Deluxe (all BioLegend), and Invitrogen™ eBioscience™ Human PMN-Elastase Platinum ELISA Kit (Thermo Fisher Scientific). Absorbance was measured at 450 nm on a plate reader (SpectraMax) with the software SoftMax® Pro V5 (both Molecular Devices, Sunnyvale, CA).
Flow cytometry
PMNs were harvested by washing the scaffolds and cell culture dishes with PBS several times. Cell containing PBS was collected and the staining procedure was performed as described earlier. 34 In brief, cells were washed (PBS, 1% fetal calf serum [Biochrom], 0.5% sodium azide [Sigma]) and 50 μL antibody mixture was added and incubated for 30 min at 4°C. The following human-specific antibodies were used: CD11b-AF488 (1:100; BD Bioscience, San Jose, CA), CD63-PE (1:20; BioLegend), CD66b-PerCPCy5.5 (1:200; BioLegend), CD43-PE/Cy7 (1:1000; BioLegend), CD35-APC (1:400; eBioscience/Thermo Fisher Scientific), CD62L-APCCy7 (1:20; BioLegend), CD16b-VioBlue (1:200; Miltenyi Biotec), and LIVE/DEAD® Fixable Aqua Dead Cell Stain Kit, for 405 nm excitation (Thermo Fisher Scientific). Samples were fixed (1% paraformaldehyde), measured at BD FACS Canto II (BD Biosciences), and analyzed by BD FlowJo software V10 (TreeStar, Inc., Ashland, OR).
Scanning electron microscopy
After 4 h of PMN culture on matrices, the medium was carefully removed and scaffolds were washed once gently with PBS. Subsequently, scaffolds were prepared as previously described26,34 and imaged at the JCM 6000 benchtop scanning electron microscopy (SEM) (JEOL, Freising, Germany) in high vacuum mode at 10 kV.
Statistics
Results are depicted as the mean ± standard error of the mean. Statistical differences between two groups were analyzed using the nonparametric Mann–Whitney test. A p-value of p < 0.05 was considered significant with *p < 0.05, **p < 0.01, and ***p < 0.001. Statistical analysis was performed using GraphPad Prism software V5 (GraphPad Software, San Diego, CA).
Results
Development of the assay design
A defined experimental procedure was developed to analyze the reaction and activation profile of PMNs after contact with biological matrices. Human PMNs from the blood of healthy donors were isolated by density gradient centrifugation. For comparing and determining the impact of cell culture conditions, surface marker expression on PMNs was analyzed by flow cytometry (FACS) immediately after isolation (t0; Fig. 1A). Subsequently, different stimulation strategies with cytokines, chemokines, as well as LPS and fMLP were compared with suitable activation markers. Therefore, PMNs were cultured in the presence of the activation stimuli for 4 h, and subsequently surface marker expression and released cytokines at 4 h time point (t4) were analyzed by FACS and ELISA, respectively (Fig. 1B). The most effective proinflammatory stimulation strategy was selected to define an appropriate activation control for later application in the assays with both types of biomaterials (cryopreserved and decellularized). Then, activation behavior of PMNs after a direct PMN culture on matrices for 4 h (t4; Fig. 1C) was studied.
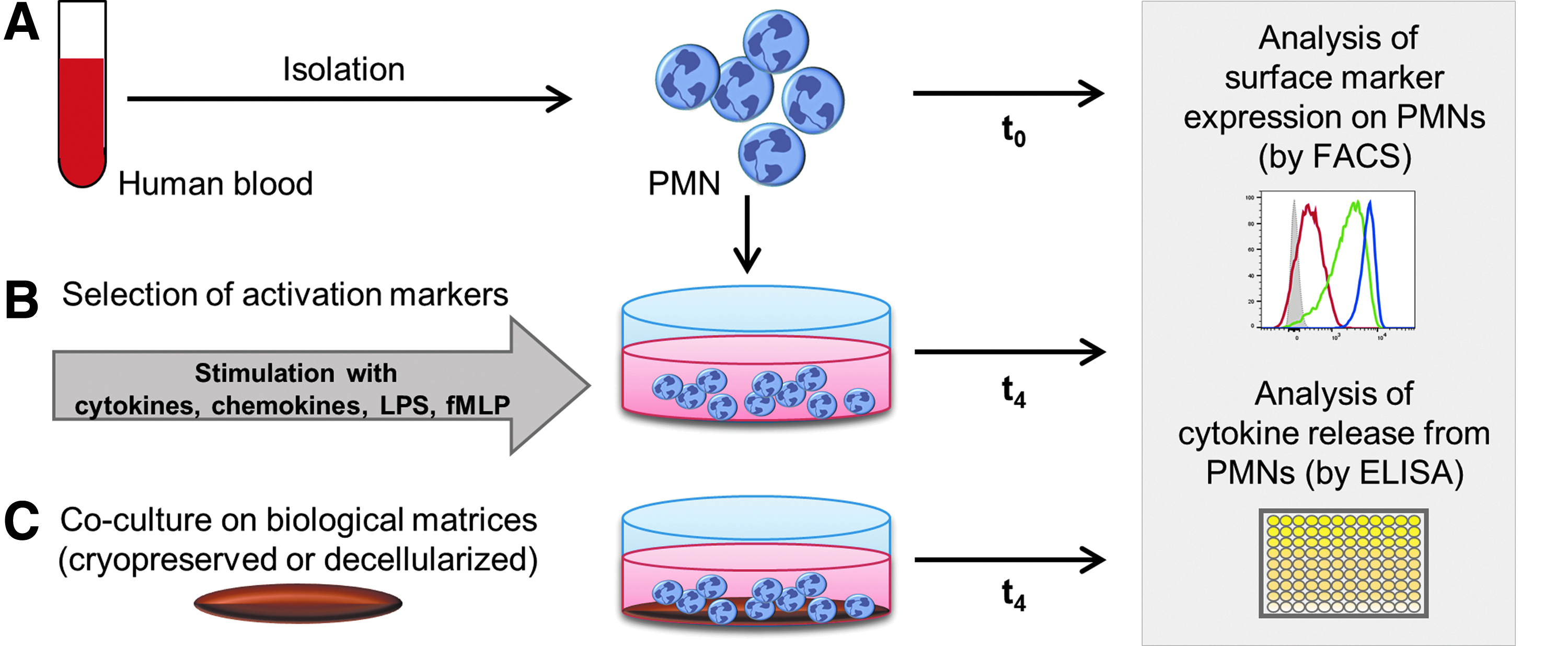
Experimental design of PMN activation assay and interaction with biological matrices.
The PMN phenotype is solely affected by standard cell culture conditions
To determine PMN surface marker expression, cells were analyzed by flow cytometry using a defined gating strategy (Fig. 2A). The size and granularity of the isolated PMN population were not homogeneous as seen in the flow cytometric scatter plots (forward scatter area [FSC-A] vs. sideward scatter area [SSC-A]). In general, PMNs were characterized by high granularity and size, usually separated into two subsets in the scatter (Fig. 2A, I). Already without adding any further stimuli, the phenotype of PMNs changed after cell culture compared with the phenotype directly after isolation from blood (t0) (Fig. 2B). The surface molecules CD11b, CD35, and CD66b were not affected by culture conditions, whereby CD66b showed in general a weak expression level. The expression of CD16b was only slightly reduced after 4 h in cell culture. Remarkably, CD43, CD62L, and CD63 showed a clear downregulation of their expression solely by culturing under standard culture conditions at t4 compared with t0.

Surface marker expression on PMNs after isolation (t0) and after in vitro culture without stimulation (t4).
LPS activation induces CD63/CD66bhigh and CD16b/CD43/CD62Llow expression on PMNs in vitro
After defining the PMN marker signatures in standard culture conditions, the impact of several activation molecules known from published protocols was examined. The aim was to choose the most effective activation strategy in vitro. Morphological appearance of the PMNs by light microscopy did not reveal striking differences after application of the selected stimuli (data not shown). Considering that PMN marker expression is already altered in culture without stimulation (Fig. 2B), the mean fluorescence intensities (MFIs) of all tested markers at t4 were normalized to the MFIs of the marker on PMNs at t0. In our setting, only some stimuli were able to modulate the expression of the analyzed surface markers. For CD11b and CD35, none of the applied trigger molecules was able to affect its expression level (Fig. 3A, C). CD63 was slightly, but not significantly, higher expressed after stimulation with LPS and TNF-α (Fig. 3F). CD66b expression was elevated by trend after stimulation with TNF-α, fMLP, and GM-CSF and significantly increased after LPS stimulation (Fig. 3G). CD16b and CD43 were equally downregulated by LPS and TNF-α stimulation (Fig. 3B, D) and for CD62L also fMLP induced a clear decrease in surface marker expression (Fig. 3E). Also important, for the markers CD63, CD66b, CD16b, and CD43, a clear dose-dependent effect by LPS was detected (Supplementary Fig. S1). Triggering by IFN-γ, IL-15, and GM-CSF had no effect. In summary, upregulation of CD63 and CD66b combined with a downregulation of CD16b, CD43, and CD62L clearly indicates PMN activation and were selected for the continued analyses. In contrast, the markers CD11b and CD35 were not further included.

LPS represents the most effective inducer of PMN activation during in vitro culture (t4). PMNs were cultured for 4 h and stimulated with INF-γ, IL-15, LPS, TNF-α, fMLP, GM-CSF, or without a stimulus (Ctrl.). After harvesting, PMNs were stained with fluorochrome-labeled human-specific antibodies and analyzed by flow cytometry. The surface marker expression of CD11b
Measuring secreted cytokines after different PMN stimulations, a significantly elevated IL-6 secretion was detected only after activation with LPS. Even though the release of MIP-1β and IL-8 was by trend enhanced after LPS and TNF-α stimulation, changes did not achieve significance (Supplementary Fig. S2). Also IL-10, IL-15, TNF-α, and PMN-Elastase were measured, but IL-15 and TNF-α were not detectable. PMN-Elastase and IL-10 showed high donor variability and no clear trend was noticed (data not shown).
Structure of biological matrices determines PMN attachment and morphology
To assess the activation status of PMNs as one of the important aspects of immunological performance of biological matrices, PMNs were cultured directly on the surface of two types of matrices as “proof-of-principle” materials: (1) differently cryopreserved (CFC or IFC) aortic tissue, which still contains cells, and (2) DeAM with or without hcECM-HG coating (DeAM+E or DeAM, respectively). First, cultured PMNs were visualized on both scaffold types by SEM (Fig. 4). Depending on the structure of the matrices resulting from the preparation method, we found modified adhesion characteristics of PMNs. PMNs cultured on aortic tissue (Fig. 4A, B) appeared to be more round shaped than cultured on DeAM (Fig. 4C, D). The IFC aortic tissue (Fig. 4A) provided a smooth surface and adherent PMNs were spread out uniformly. However, CFC technique resulted in a rougher tissue surface and induced clustered accumulation of PMNs (Fig. 4B). PMNs cultured onto both types of the DeAM showed differences in morphology. PMNs on DeAM+E demonstrated a more heterogeneous morphology than cells on DeAM without coating. Although the hcECM-HG coating provided an additional nanoscaffold onto the membrane, PMN adhesion to DeAM+E was not altered (Fig. 4C) compared to DeAM without hcECM-HG coating (Fig. 4D).
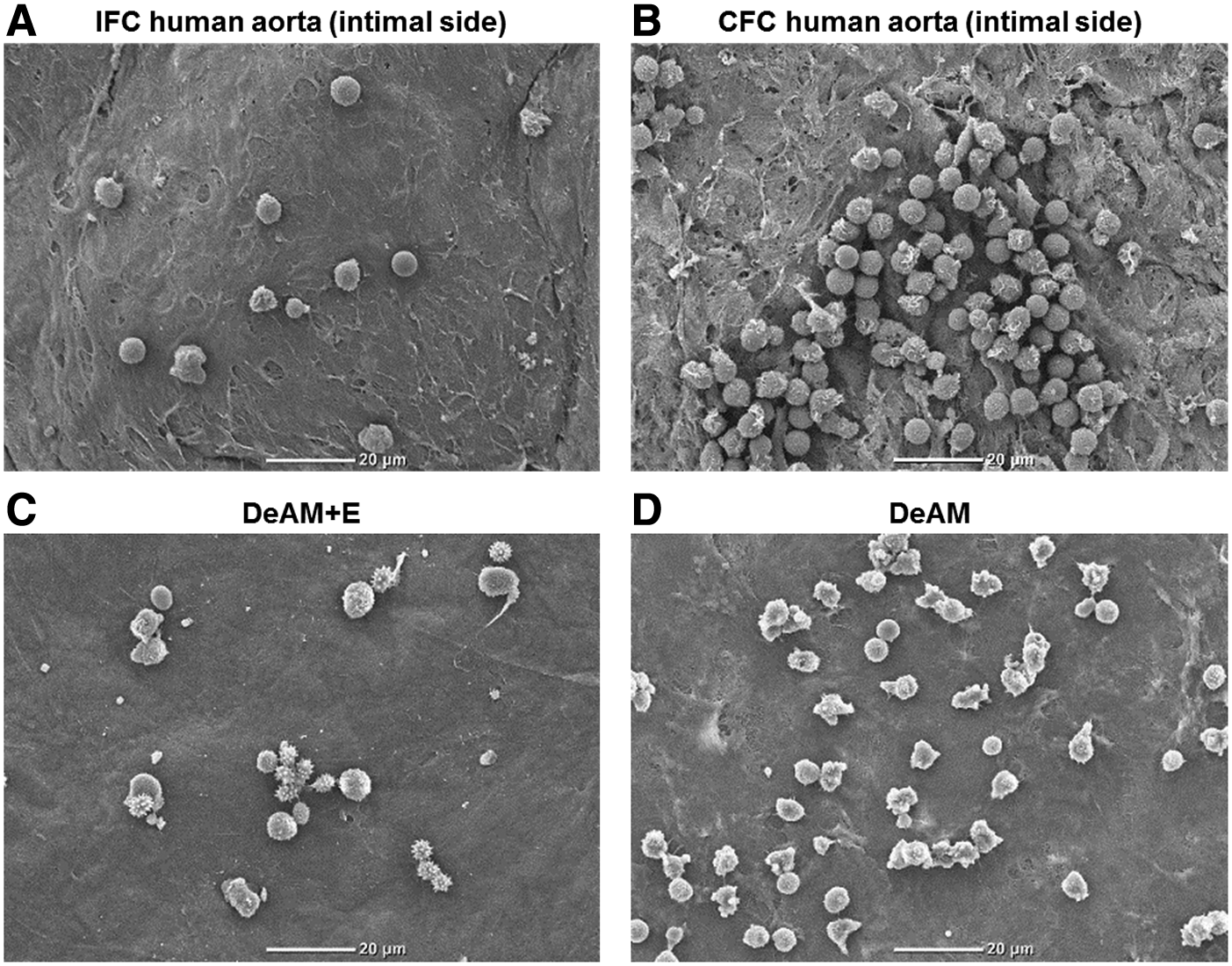
Morphology and adherence of PMNs cultured on cryopreserved aortic tissue and DeAM. Morphological analysis of PMNs cultured for 4 h on both investigated matrix types was performed by scanning electron microscopy. Representative images are shown for human aortic tissue after IFC
The PMN assay is able to assess and discriminate varying immunological properties of biological matrices
PMNs were cultured for 4 h onto aortic tissue cryopreserved by the IFC or CFC technique (Fig. 5) and on DeAM+E or DeAM (Fig. 6) and analyzed by flow cytometry. For the purpose of analysis, data from PMNs stimulated with LPS acquired during the assay establishment were added to the activation control group, and again, all data were normalized to the marker expression of unstimulated PMNs at t0. Furthermore, to exclude false positive results for the detection of PMN activation, materials were initially tested for endotoxin contamination. By Limulus Amoebocyte Lysate tests, CFC and IFC human aortic tissue and DeAM+E and DeAM were shown to be endotoxin free (data not shown).
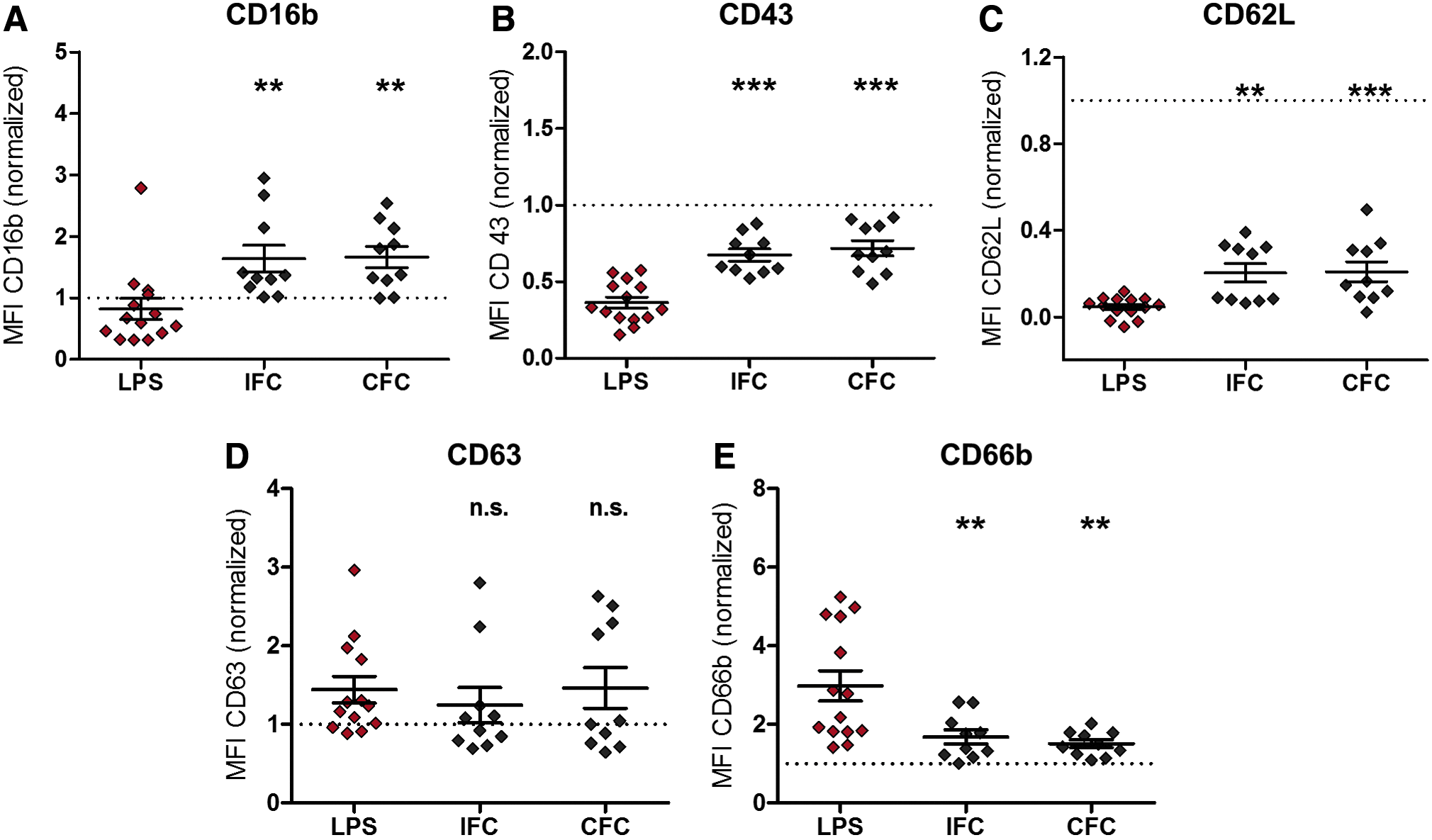
Both types of cryopreserved human aortic tissues do not induce PMN activation. After 4 h coculture on IFC and CFC aortic tissue, PMNs were harvested, stained with fluorochrome-labeled human-specific antibodies against CD16b
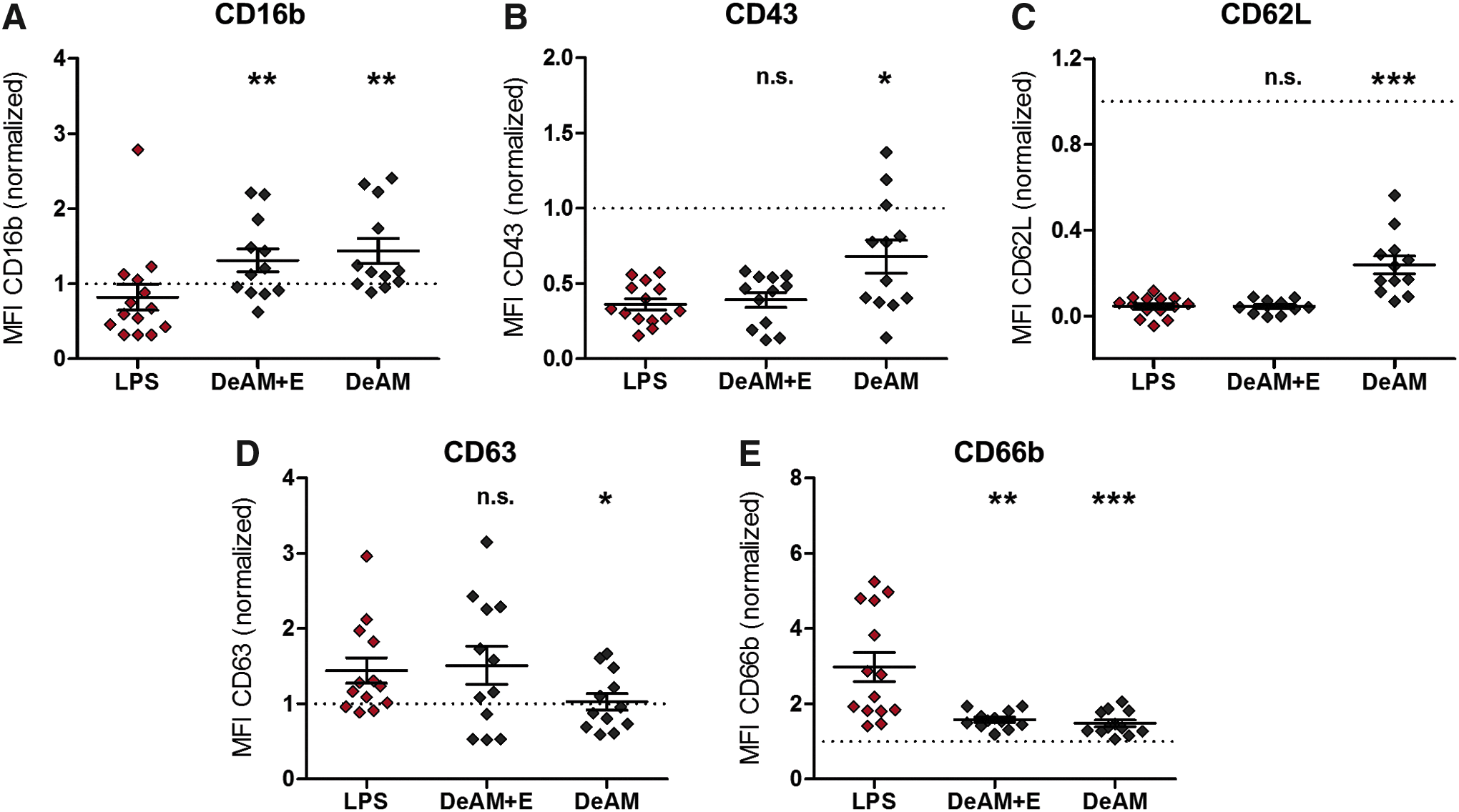
DeAM with hcECM-HG coating (DeAM+E) triggers changes in PMN phenotype but DeAM without coating does not induce PMN activation. After 4 h coculture on DeAM and DeAM+E, PMNs were harvested, stained with fluorochrome-labeled human-specific antibodies against CD16b
After contact with both cryopreserved aortic tissues, PMNs significantly upregulated expression of CD16b, CD43, and CD62L (Fig. 5A–C) and downregulated CD66b compared with PMNs stimulated with LPS (Fig. 5E). Only for CD63 expression, no difference compared with the LPS control was detected (Fig. 5D). Overall, no difference in surface marker expression on PMNs cultured on either CFC or IFC tissue was observed. As reported earlier, cryopreserved aortic tissues itself still secret high amounts of cytokines such as IL-6 and IL-8. 26 Compared with that, the cytokine amount secreted solely by PMNs is very low. Therefore, the “additional cytokine secretion” of PMNs would be in the variability range of the tissue punches. Although all aortic tissue punches had the same size, the released cytokine levels show variations and prevent to define an exact background level. Since a discrimination between the cytokines released from PMNs and tissue was not feasible, using the cytokine release as activations markers was not applicable for CFC and IFC aortic tissue.
After culturing PMNs on cell-free amniotic membranes, their surface marker expression was altered. Similar to PMNs cultured on aortic tissue, the expression of CD16b was significantly increased and CD66b was significantly decreased upon contact with both amniotic membrane types (DeAM+E and DeAM) compared with the LPS control (Fig. 6A, E). In contrast, the expression of CD43, CD62L, and CD63 on PMNs varied after culture on both tested amniotic membrane types (Fig. 6B–D). In this study, the expression level on PMNs cultured on DeAM+E was similar to the expression level of the LPS control group. However, PMNs cultured on DeAM without hcECM-HG coating expressed CD43, CD62L significantly higher and CD63 significantly lower compared with the LPS control.
As these results suggest a PMN activation by DeAM+E, the induced secretion of IL-6, MIP-1β, and IL-8 was measured (Fig. 7). No difference in the cytokine levels between PMNs cultured either on DeAM+E or DeAM was observed, and furthermore, the measured cytokine concentration was always significantly lower than the LPS control.
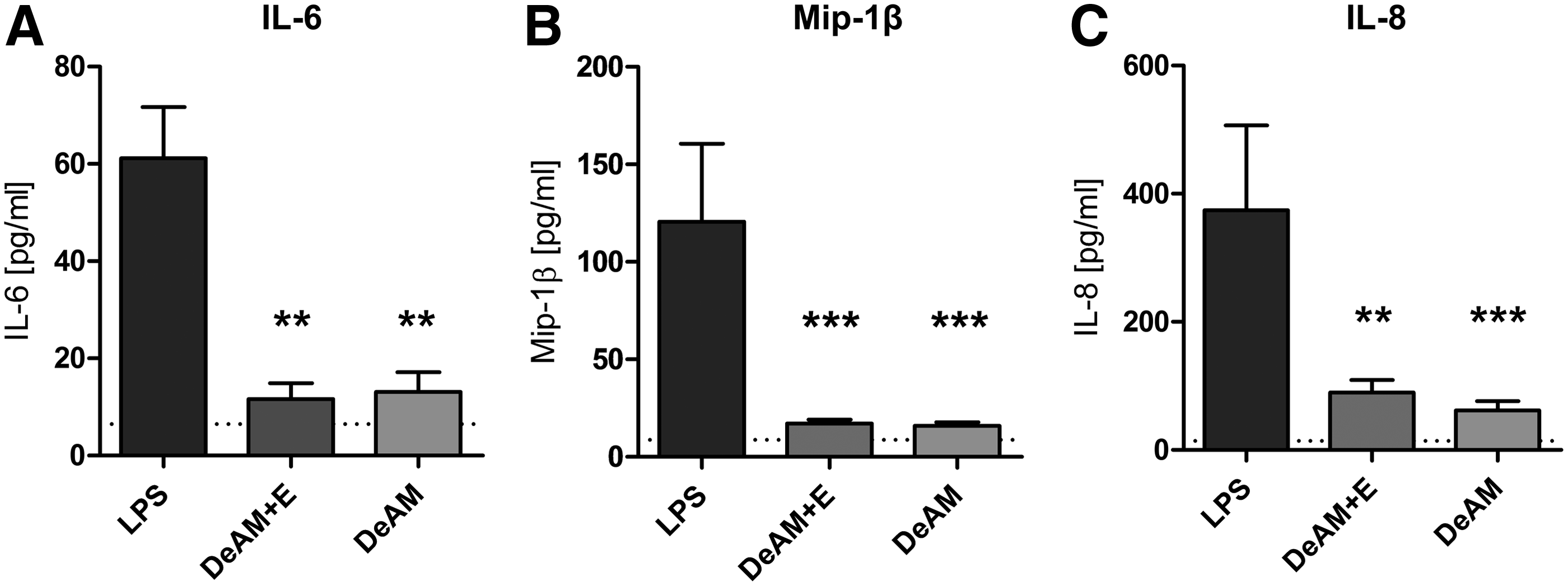
Cytokine release of PMNs after coculture on DeAM+E and DeAM. PMNs were cultured for 4 h on DeAM+E or DeAM. Cytokine/chemokine concentration in the coculture supernatant was measured for IL-6
Discussion
In general, CVDs cause an irretrievable loss of functional tissue, and therapeutic interventions include tissue replacements or bypasses. Furthermore, experimental approaches use intramyocardial injections3,36 or epicardial patches.34,37 However, the different types of implants would cause immune reactions, 6 with PMNs representing the first interacting immune cell subset. 38 So far, existing immunocompatibility assays in vitro and in vivo payed more attention to other immune cell subsets such as monocytes, macrophages, or T cells.28,39–43 Thus, there is a need to establish a simple, fast, and reliable PMN-based in vitro assay to investigate also PMN behavior to further assess the immunogenicity and compatibility of cardiovascular implants.
To evaluate PMN activation in vitro, different strategies using INF-γ, IL-15, TNF-α, GM-CSF, LPS, or the PMN activator peptide fMLP have been applied.17,44–48 However, experimental results were not comparable, thus not allowing a standardized investigation of PMN–material interactions. Yet, only in clinical studies it was shown that PMN surface molecule expression and the cytokine secretion profile were altered, for example, in neuronal diseases, gingivitis or chlamydia infection.48–52 A suitable assay for monitoring the PMN-to-implant reaction in vitro can close the gap between clinical and in vitro observations.
Interestingly, although we validated LPS as an effective PMN activator, we could not confirm published changes of CD35 expression upon LPS activation. CD35 is a mediator for immune adherence or phagocytosis 53 and was found to be upregulated on PMNs after chlamydia infection. 51 Moreover, we could not verify CD11b as a reliable activation marker in vitro, although CD11b was shown to be downregulated on PMNs in clinical scenarios. 52 Possibly this might be explained by a much longer observation time in man compared with the PMN lifespan in vitro.
Besides PMN stimulation with TNF-α and fMLP, mainly LPS induced a significant upregulation of CD66b and CD63 and a reduction of CD16b, CD43, and CD62L. These LPS-triggered effects were dose dependent. Similar results for CD62L, CD63, and CD66b were observed earlier in vivo in a humanized mouse model. 54 This supports our suggestion of employing a CD63/CD66bhigh and CD16b/CD43/CD62Llow PMN phenotype as a reliable hallmark for PMN activation in vitro.
However, it should be noted that PMNs without additional stimulus underwent some basal activation after 4 h culture in vitro mainly reflected by the downregulation of CD43 and CD62L, as the mechanical isolation and cultivation process already triggers an activation. 55 Therefore, we additionally measured the phenotype of freshly isolated PMNs (t0) to consider that PMNs potentially can undergo a sensitive change in their phenotype. In contrast to other experimental attempts,11,17 it must be emphasized that all our data measured after 4 h were normalized to the values at time point t0. In our view, this normalization step is essential for accurate PMN analysis.
After determining suitable PMN activation markers, in a next step the suitability of the assay for screening of clinically relevant implant materials should be analyzed. Because clearly categorized low- or high-immunogenic materials are hardly to define, always the LPS activation control and a negative control without any stimulus should be performed in parallel with the matrices. A material with low PMN activation potential would lead to a PMN phenotype comparable with the negative control and should differ significantly from the phenotype of LPS-activated PMNs. In “proof-of-principle” experiments for assessing the immunogenicity of cardiovascular allografts, we first used aortic tissue preserved according to IFC or CFC.24,26 Furthermore, as a model for potential treatment of myocardial infarction, DeAM with or without hcECM-HG coating was analyzed by our newly established assay. 34
PMNs adhered on both biological materials but showed various morphologies. That different matrices selectively modulate adhesion and viability of PMNs was already shown by Cohen et al. for nonbiological materials. 11 Cryopreservation or decellularization of biological matrices has an impact on the structure and composition of the material, which might be the reason for immune cell activation.23,56,57 Upon contact with CFC and IFC aortic tissue, PMNs revealed a similar surface marker expression pattern. However, with exception of CD63, all markers were significantly different from the LPS activation control, which demonstrates that neither type of cryopreserved matrices induces PMN activation. In line with this, former studies of our group showed that both types of cryopreserved aortic tissue were not able to induce a polarization toward the proinflammatory M1-macrophage phenotype. 26
Interestingly, DeAM+E but not DeAM induced an activated phenotype of PMNs regarding expression of CD43, CD62L, and CD63. The hcECM-HG for coating was produced by decellularization of human cardiac tissue followed by pepsin-based homogenization, 33 and collagen, which is an ECM component, might act as an extracellular damage-associated molecular pattern molecule and chemoattractant for PMNs.58,59 Furthermore, ECM degradation could lead to peptide fragments, which can be recognized by PMNs and induce activation, as recently detected for DeAM+E. 60
However, not all surface markers on PMNs upon DeAM+E contact indicated an activation. Measuring cytokine release from the PMNs cultured on the membranes, we could show that neither IL-6, MIP-1β, or IL-8 was elevated in PMNs cultured on the amniotic membranes. Taken into account that activation of PMNs is characterized by differential surface marker regulation and cytokine secretion, it can be concluded that DeAM+E does activate PMNs to some extent, but rather incompletely. Functionally it might imply that PMNs will get the signal to migrate into the DeAM+E in vivo (shedding of CD43 and CD62L) but would not secret high amounts of proinflammatory cytokines. A reduced IL-6 secretion would then also lead to a reduced recruitment of monocytes10,61 and, therefore, attenuate the subsequent immune response. Own previous results support this assumption, wherein monocytes cultured on DeAM and DeAM+E showed reduced proinflammatory cytokine secretion. 34
This demonstrates that both, surface marker expression and—if applicable—cytokine secretion of PMNs, should be measured within the PMN assay to anticipate how the response of other immune cells like monocytes is influenced. In our case, only cytokine measurement for PMNs cultured on DeAMs was feasible, since no cytokines are secreted by the patch material. 31 In contrast, in case of the cell containing tissue such as CFC and IFC aorta, cytokine detection was not applicable, because especially from CFC aortic tissue itself, very high levels of cytokines (like IL-6 and IL-8) are released. 26 That will overlap the measurement of cytokines secreted by PMNs. Future studies should additionally include different LPS concentrations as control groups to support a better assessment of the degree of the immune reaction.
In conclusion, we developed an easy and reliable assay to evaluate PMN responses to biological matrices in vitro, to assess a possible immunogenicity or compatibility of the material of interest. For the assay procedure, LPS stimulation appeared to be the most potent activation control. Furthermore, we could show the importance of analyzing both, surface marker expression at t0 and t4. Also, if possible, measuring released proinflammatory cytokines at t4 is beneficial to characterize the PMN response mechanism more precisely. Overall, the presented PMN assay provides an additional tool to answer whether PMNs play an important role in initiating the immune response to an implanted material and help to understand the overall picture of immune response mechanism to that particular biomaterial. However, for a more comprehensive understanding and prediction of the immune response toward matrices in vivo, it would be essential to investigate further innate and adaptive immune cells, since especially monocytes and T cells might have a stronger impact on the final outcome of the immune reaction. In addition, including the evaluation of NETosis, which is another important feature of PMNs during cell–scaffold interaction, would complement the gained insights. 21 Moreover, the impact and type of reaction of the immune cell subsets are probably very unique for each material. Therefore, the introduced PMN assay would represent an essential element in clinical implant evaluation procedures.
Footnotes
Acknowledgments
This study was supported by funding of the Berlin-Brandenburg School for Regenerative Therapies (BSRT, Graduate School 203 of the DFG Excellence Initiative) and partially by the German Research Foundation (Deutsche Forschungsgemeinschaft, grant DFG SE 657/12–1 [M.S.]). The authors thank Janita Aline Maring for proof reading the article and acknowledge the assistance of the BIH Cytometry Core-Facility.
Authors' Contributions
M.Sch. and M.B. carried out study design, performance of experiments, data acquisition, analysis and interpretation, and writing the article. M.S. and C.S. interpreted the data and substantively revised the article.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.