
Abstract
Omics technologies, such as genomics, epigenomics, transcriptomics, proteomics, metabolomics, lipidomics, multiomics, and integrated modalities, have greatly contributed to our understanding of various diseases by enabling researchers to probe the molecular wiring of cellular systems in a high-throughput and precise manner. With the development of tissue-engineered three-dimensional (3D) in vitro disease models, such as organoids and spheroids, there is potential of integrating omics technologies with 3D disease models to elucidate the complex links between genotype and phenotype. These 3D disease models have been used to model cancer, infectious disease, toxicity, neurological disorders, and others. In this review, we provide an overview of omics technologies, highlight current and emerging studies, discuss the associated experimental design considerations, barriers and challenges of omics technologies, and provide an outlook on the future applications of omics technologies with 3D models. Overall, this review aims to provide a valuable resource for tissue engineers seeking to leverage omics technologies for diving deeper into biological discovery.
Impact statement
With the emergence of three-dimensional (3D) in vitro disease models, tissue engineers are increasingly interested to investigate these systems to address biological questions related to disease mechanism, drug target discovery, therapy resistance, and more. Omics technologies are a powerful and high-throughput approach, but their application for 3D disease models is not maximally utilized. This review illustrates the achievements and potential of using omics technologies to leverage the full potential of 3D in vitro disease models. This will improve the quality of such models, advance our understanding of disease, and contribute to therapy development.
Introduction
Over the last decade, omics technologies, namely genomics, epigenomics, transcriptomics, proteomics, metabolomics, lipidomics, multiomics, and integrated modalities, have helped unmask the molecular wiring of cellular systems in a high-throughput and precise manner. Early applications of these technologies have focused extensively on precision medicine applications by investigating patient samples for disease diagnosis and biomarker identification.1–4 These omics technologies also offer the opportunity to molecularly dissect disease mechanisms and identify novel targets. For these mechanistic applications, it is not typically possible to probe specific questions to link genotype to phenotype for distinct cell populations in the tissue, and it is challenging to perform spatiotemporally resolved omics to understand heterogeneity and microenvironmental impact. Furthermore, the use of patient samples is not always ideal as these are not widely available for all diseases. Recently, three-dimensional (3D) in vitro disease models, such as patient-derived organoids (PDOs) and spheroids, have emerged as valuable engineered constructs that can recapitulate the in vivo microenvironment, but can be more easily manipulated to decipher the relationship between genotype and phenotype. While 3D in vitro models have also been used to study tissue development, in this study, we focus solely on disease modeling. In this review, we provide an overview of existing omics technologies, discuss the current and emerging applications of omics technologies with 3D disease models, and highlight the experimental design considerations, barriers, and challenges of applying omics to 3D disease models. The goal of this review is to provide a resource for tissue engineers seeking to leverage omics technologies for biological discovery, and to provide an outlook on the future of omics and 3D disease models in unraveling the intricate molecular networks that control disease.
Overview of Omics Technologies
Several omics technologies have emerged as new tools to probe specific cellular properties, namely genomics, epigenomics, transcriptomics, proteomics, lipidomics, metabolomics, multiomics, and integrated modalities (Fig. 1). A short description of the various omics methods can be found in Box 1, whereas a brief overview of the general workflow of each technology is shown in Figure 2. A nonexhaustive list and description of omics methods can also be found in Supplementary Table S1 and is meant to function as an initial reference for those unfamiliar with the variety of available omics technologies.
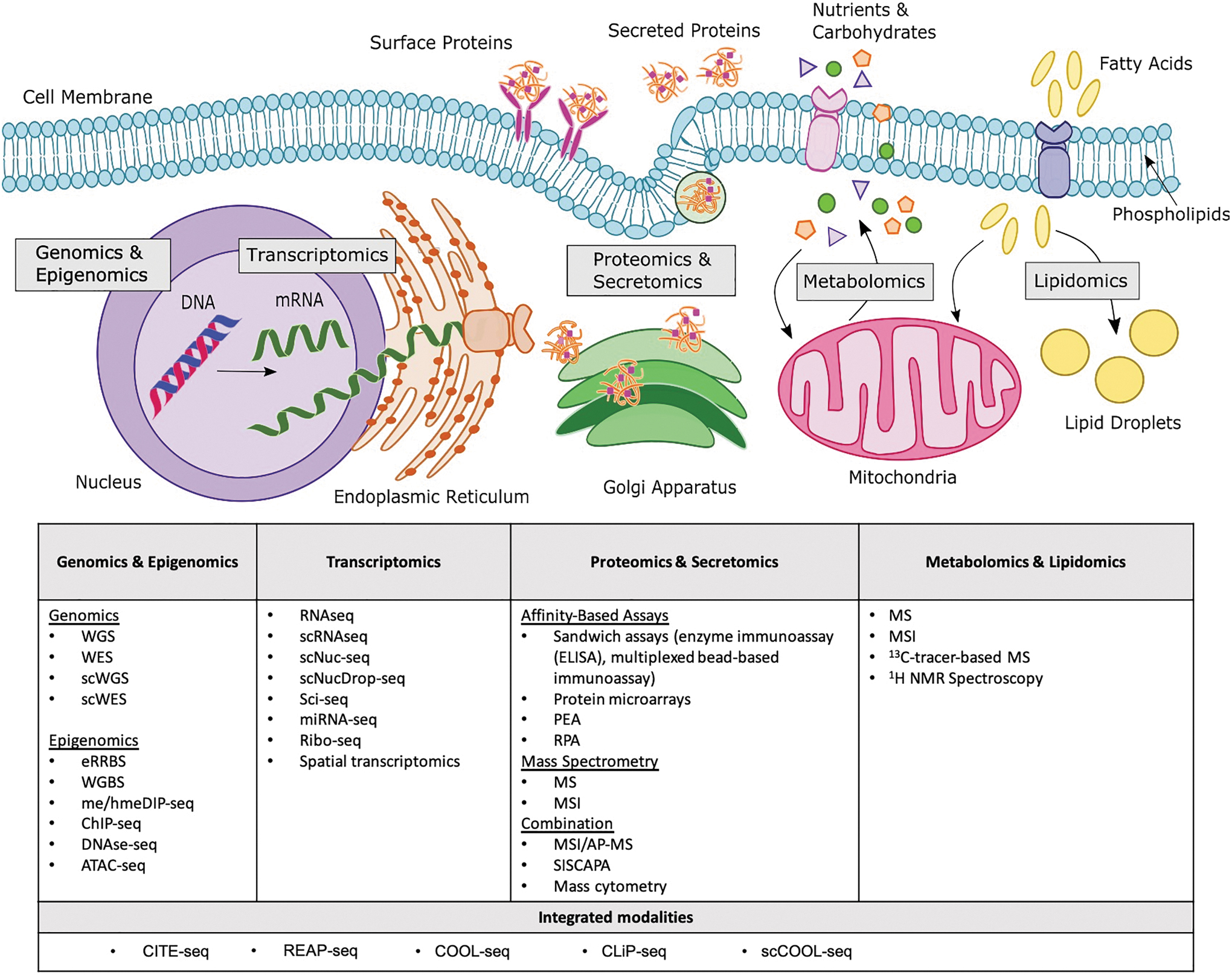
Overview of omics technologies and which elements of the cell they probe. Genomics and epigenomics analyze nucleic DNA; transcriptomics analyzes all RNA molecules (from messenger RNA in the nuclei to ribosomal RNA in the endoplasmic reticulum); proteomics and secretomics analyze proteins that are synthesized in the endoplasmic reticulum and transported to the membranes or lysosomes or secreted through the golgi apparatus and secretory vesicles; metabolomics analyzes metabolites produced and consumed by the cell and processed in the mitochondria; lipidomics analyzes lipid species that are located in membranes and organelles and play a role in metabolic processes. See Supplementary Table S1 for the definition of acronyms and a brief description of the omics technologies. Color images are available online.
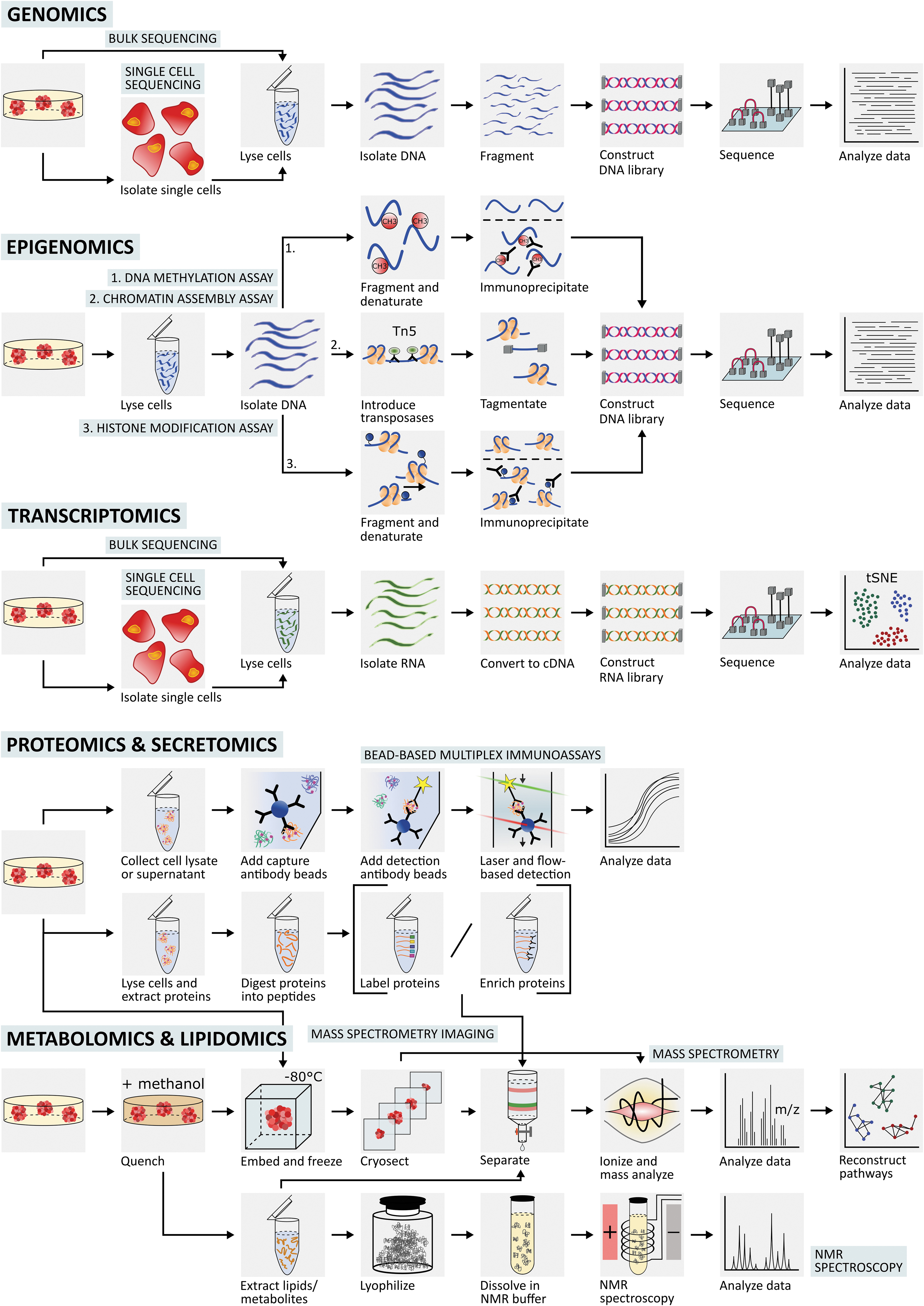
Brief and general overview of the workflow and sample preparation of each omics technology from the 3D in vitro disease model to the data analysis. For omics analysis of EV molecular cargo, such as miRNA, lncRNA, lipids and proteins, an extra ultracentrifugation step (not shown in the figure) is performed to isolate the secreted EVs in the cell culture media before any specific extraction method to isolate cargo material for the appropriate omics strategy. 3D, three-dimensional; EV, extracellular vesicle; lncRNA, long-noncoding RNA; miRNA, microRNA; NMR, nuclear magnetic resonance. Color images are available online.
Current Landscape of Omics Technologies and 3D In Vitro Disease Models
Having provided an overview of some of the key omics technologies available, in this section we will highlight the types of studies to date that have combined omics technologies with 3D in vitro disease models. Due to the constraints of this review, we highlight only a few key examples in each case and Table 1 is not exhaustive.
Brief Descriptions of Different Omics Technologies, Namely Genomics, Epigenomics, Transcriptomics, Proteomics, Metabolomics, and Lipidomics
Omics Technologies Applied to Three-Dimensional In Vitro Disease Models
Overview of studies that have used omics technologies to address questions related to 3D in vitro disease models. The omics technologies in this table are described in as much detail as the specific study provided. The definition of the acronyms of the omics technologies and a brief description can be found in Supplementary Table S1.
Multiomics study.
Study applied proteomics and secretomics.
Study applied secretomics.
2D, two-dimensional; 3D, three-dimensional; ATAC-seq, Assay for Transposase-Accessible Chromatin with NGS; ChIP-seq, chromatin immunoprecipitation with massively parallel DNA sequencing; E, epigenomics; ELISA, enzyme-linked immunosorbent assay; eRRBS, enhanced reduced representation bisulfite sequencing; ESC, embryonic stem cell; G, genomics; iPSC, induced pluripotent stem cell; L, lipidomics; M, metabolomics; MDOTS, murine-derived organotypic tumor spheroid; meDIP-seq, methylated DNA immunoprecipitation in combination with next-generation sequencing; miRNA, microRNA; MS, mass spectrometry; MSI, mass spectrometry imaging; NGS, next-generation sequencing; NMR, nuclear magnetic resonance; P, proteomics; PD, patient-derived; PDO, patient-derived organoid; PDOTS, patient-derived organotypic tumor spheroid; PDS, patient-derived spheroid; PDX, patient-derived xenograft; RNA-seq, RNA-sequencing; scRNA-seq, single-cell RNA sequencing; T, transcriptomics; WES, whole exome sequencing; WGS, whole genome sequencing; CyToF, mass cytometry by time of flight; DIGE, difference gel electrophoresis; miRNA-seq, microRNA sequencing; RPA, reverse phase protein microarray; sc-WGS, single-cell whole genome sequencing; SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis.
Genomics technologies for 3D in vitro disease
Genomics technologies, such as next-generation sequencing (NGS), have been applied to the complete characterization of the genotype of 3D disease models to classify disease subtypes. For example, model characterization by means of genomics, specifically NGS, has contributed to the creation of “living biobanks” of PDOs and patient-derived xenografts.28,29 These biobanks have greatly improved the availability of such models, and have enabled many groups to investigate disease phenomena associated with specific patient subtypes in these 3D in vitro systems.
Building on model characterization, another key application of genomics in the context of 3D disease models has been model validation by comparing the disease model to patient tissue data, to determine to what extent the model resembles the in vivo disease genotype. For example, whole exome sequencing (WES) and whole genome sequencing (WGS) were used to investigate how the mutational landscape of 3D tumor models recapitulates that of patient tumor samples.28–32 WES and WGS were further used to demonstrate that organoids capture the intratumoral heterogeneity, an important feature of in vivo tumors.28,33,34
With validated models now emerging, a combination of genomics and tissue models has now begun to contribute to the identification of gene signatures that are predictive of response to therapy for personalized medicine applications. 35 For example, studies using WGS 36 and WES 31 have facilitated the understanding of how drug sensitivity is related to genomic mutation profile in PDOs. Interestingly, these correlations were not detectable in monolayer cultures, 36 highlighting the relevance of 3D disease models. Furthermore, another emerging application for genomics studies of 3D disease models is the investigation of disease mechanisms and progression. For example, WGS has facilitated the probing of clonal evolution in PDOs to discover cancer drivers, 37 while WES was used to map the accumulation pattern of mutations in patient-derived adult stem cell organoids to investigate tumorogenesis. 38 Additionally, whole-genome perturbations in clonal organoids, executed by CRISPR/Cas9 and screened with NGS, have enabled the precise identification of mutations that confer therapy resistance.39,40 As more validated models emerge, such approaches will be useful to stratify patients for the purpose of developing precision medicine applications.
Epigenomics technologies for 3D in vitro disease models
Epigenomics technologies are valuable tools for probing 3D disease models due to the discovery of distinct epigenetic signatures that associate with specific disease phenotypes. In the context of 3D disease models, epigenomics has therefore most frequently been used to address disease mechanisms associated with specific phenotypes and therapy resistance. For example, the coupling of chromatin immunoprecipitation with massively parallel DNA sequencing (ChIP-seq) identified the DNA-binding sites of transcription factors critical to prostate41,42 and colon 43 oncogenesis in organoid models. Additionally, the role of chromatin accessibility in the chemoresistance colon cancer organoids was investigated using the Assay for Transposase-Accessible Chromatin with NGS (ATAC-seq). 44 Similarly, epigenomics has facilitated the investigation of the alterations of epigenetic signatures upon stimulation with environmental factors that impact phenotypes. For example, ChIP-seq demonstrated the effect of stromal cues on histone acetylation in a 3D pancreatic cancer model. 45 Furthermore, methylated DNA immunoprecipitation in combination with next-generation sequencing (meDIP-seq) was employed to investigate the DNA methylation profile of cardiac and hepatic spheroids in response to stimulation with a toxic agent. 46 In addition, as with genomic analysis, one of the earliest applications of epigenomics has been to validate novel 3D disease models by comparing to patient samples. For example, ChIP-seq has been employed to investigate if organoids recapitulate epigenetic characteristics of patient tissue. 43 Additionally, enhanced reduced representation bisulfite sequencing (eRRBS) has demonstrated correlation of prostate cancer organoid DNA methylation to their corresponding patient tissue samples. 47
Increasingly emerging are multiepigenomics approaches that provide more insight into the various types of epigenetic signatures and their association to disease. For example, the combination of ChIP-seq and ATAC-seq has allowed the analysis of DNA/protein-binding sites as well as open chromatin sites of 3D tumor models, correlating their broad epigenetic profile to metastatic transition. 48 Such integrative approaches to epigenomics have the potential to elucidate the full regulatory mechanism that determines gene expression and eventually cell phenotype.
Transcriptomics technologies for 3D in vitro disease models
Transcriptomics is perhaps the most widely used omics technology with 3D in vitro disease models and has been applied to answer a variety of questions. As with genomic analysis, transcriptomics studies have mainly been driven by bulk RNA sequencing (RNA-seq) for the purpose of validating models by comparing them to patient tissue, as shown in Table 1, further highlighting the importance of this initial validation step to confirm the relevance of 3D disease models for biological discovery. Often, these studies show the ability of 3D models such as PDOs to recapitulate the patient transcriptional landscape at various disease stages, 49 as well as match patient response to therapies.28,29,47,50–55 Several model validation studies on PDOs have also aided the assessment of PDO transplantability in vivo 56 and the generation of living biobanks or centralized PDO distribution centers.28,29,52,54 These characterization efforts have also enabled the profiling of cell population heterogeneity of 3D disease models, in combination with validation and mechanism-discovery studies. Such profiling has led to the discovery of novel disease features in otherwise inaccessible tissues like the brain, using induced pluripotent stem cell-derived organoids,57–59 and to the identification of spatial 34 and temporal transcriptional changes,60–62 using PDOs. These applications would be challenging to decipher in patient samples.
Beyond model validation, one of the most powerful applications that combining transcriptomics and 3D disease models is enabling are studies driven by biological questions that are difficult to address with clinical samples. Such studies are focused on understanding the mechanisms of disease development or progression,43,63–66 and drug response.37,67 For example, the groups of Valeri 68 and Tuveson 35 treated PDOs from different patients with various chemotherapeutics and performed pharmacotranscriptomics to stratify and explain transcriptional variations to therapy response. Similarly, Romero-Calvo et al. showed that this varied therapy response was due to patient-specific clonal diversity. 51 These works especially highlight the value of 3D disease models and transcriptomics for precision medicine.
An emerging trend in transcriptomics and 3D disease models is the use of single-cell approaches. Single-cell techniques have been widely adopted in the investigation of patient samples and in vivo systems and can also address heterogeneity of 3D disease models. For example, heterogeneity associated with different cell types or heterogeneity within one cell type, for example, cancer cells, stem cells, and immune cells. However, this approach has not yet been used extensively in the context of 3D disease models, perhaps because there is a lack of existing multicellular 3D disease models requiring single-cell resolution. One of the few studies integrating single-cell techniques includes Neal et al.'s single-cell RNA sequencing (scRNA-seq) study on multicellular PDOs with autologous immune infiltrates, which showed that mechanisms of failed immunotherapy were due to expansion of exhausted T cells in particular. 32 This work emphasizes the potential of integrating single-cell approaches with physiologically relevant multicellular 3D disease models for exploring specific hypothesis-driven questions, such as deciphering cell-specific contributions to therapy resistance.
Lastly, another emerging trend is transcriptomic profiling of extracellular vesicle cargo, such as microRNA (miRNA) and long noncoding RNA (lncRNA), in 3D disease models due to their roles in disease progression,69–71 diagnostics, 72 and therapeutics. 73 For example, lncRNA has been shown to impact tumorigenicity in breast cancer organoids, 71 whereas miRNA has demonstrated potential as a novel therapeutic target in ovarian cancer organoids. 73 These transcriptomic studies performed in 3D disease models are especially difficult to do in vivo and have been mainly limited to microarray analysis (for miRNA) and bulk RNAseq (for lncRNA).
Proteomics technologies for 3D in vitro disease models
Compared with transcriptomics, the use of proteomics to investigate 3D disease models is less extensive. This can be partially attributed to the still growing and evolving field of proteomics technologies that, despite having made significant advancements, has not established standard and widely available protocols for use with 3D disease models. Nevertheless, understanding disease development and progression using proteomic profiles has provided insight in areas that cannot be assessed using genomics and transcriptomics in 3D disease models.66,74–76 Meanwhile, proteomic profiling has been more frequently used with established models, such as the two-dimensional (2D) in vitro models, in vivo murine models, and patient tissue and serum samples, and has contributed to major findings that have had clear clinical applications.77–81
In the context of 3D disease models, proteomic profiling has mainly been applied to address questions that compare 2D and 3D culture systems. Several studies have demonstrated a clear distinction between the protein profiles of 2D and 3D cultures, with the latter retaining more similarities to in vivo disease states.82–85 Similarly, the phosphoproteome, proteins that contain a post-translational phosphate group modification that guides many cellular processes such as DNA replication, apoptosis, and invasion, has also demonstrated distinct differences between 2D and 3D cultures.86,87 Moreover, proteomics has elucidated differences in drug mechanism and chemotherapeutic resistance between 2D and 3D culture model studies such as drug penetration, dose-dependent drug efficiency, and drug clearance of colorectal cancer 88 and Alzheimer's disease models. 89
A few studies have used proteomics as the foundation to compare 3D disease model data with in vivo patient data. These studies used PDOs and spheroids to demonstrate the preservation of in vivo protein expression patterns.90,91 Similarly, stratifying patient populations using 3D disease models and proteomics is not common, likely due to the lack of available proteomic patient databases. Moreover, the inherent dynamic characteristic of proteins in disease adds complexity. A single recent study is, to our knowledge, the only example of proteomic profiling of human organoids using mass spectrometry that was correlated with clinical diagnosis and importantly, addressed the interpatient heterogeneity. 92
Secretome analysis of 3D disease models has relied on immunoassay-based technologies. These studies have largely focused on the human immune system and its defining network of secreted proteins, including cytokines, chemokines, and growth factors. For example, membrane-based immunoassay arrays were used to demonstrate that the innate immune and cytokine responses in a 3D disease model recapitulated those observed in Hepatitis B virus-infected patients. 93 Additionally, bead-based multiplexed immunoassays were used to characterize the levels of cytokines and growth factors in the supernatant of multicellular spheroid tricultures, 94 and the cytokine profile of a novel 3D culture system to model immune checkpoint blockade. 67
There is huge untapped potential in enhancing disease discoveries and knowledge through the combination of 3D disease models and proteomics. There are powerful proteomics technologies, as listed in Figure 1 that have seldom or never been leveraged for 3D disease models, such as single-cell proteomics using mass cytometry 95 and spatial proteomics technologies. 96 The proteome plays a significant role in disease development and progression, as it is the main functional machinery of cells and the source of communication between cells, thereby linking heterogeneous disease cell populations. As such, there are key questions that would benefit from proteomics analysis of 3D disease models that have not yet been addressed. The application of proteomics to analyze 3D disease models is therefore an important evolving area for the future.
Metabolomics technologies for 3D in vitro disease models
Metabolomics provides a more comprehensive characterization of cellular phenotype than the omics technologies discussed above. One of the most thoroughly explored applications of metabolomics in the context of 3D disease models is the demonstration of improved relevance of 3D disease models compared with traditional 2D monolayers. For example, mass spectrometry and nuclear magnetic resonance-based metabolomics have revealed that 3D tumor models demonstrate a significant difference in metabolic profile,97–100 as well as an increase in therapy resistance101–103 in comparison to 2D monolayers. Additionally, metabolomics of 3D disease models revealed new disease pathways that were not recapitulated in traditional monolayers. 104 These studies have successfully demonstrated the use of metabolomics to highlight the importance of physiologically relevant 3D disease models for drug screening and drug target discovery.
Metabolomics has also been applied as a tool to validate if the 3D disease model appropriately recapitulates the in vivo phenotype. 105 For example, a novel 3D human/microbiome microfluidics system (HuMiX) was validated relative to datasets from clinical studies. 105 Additionally, analogous to the field of precision medicine, metabolomics has assisted in determining if a 3D model maintains disease subtype, 106 as well as donor-specific metabolic characteristics. 99
Another interesting use of metabolomics analysis in the context of 3D disease models is the investigation of disease mechanisms as well as physiological interactions. For example, metabolomics studies investigated the metabolic reprogramming of cancer cells and correlated this to their proliferative 107 or metastatic 100 phenotype in a spheroid model. Other groups have found significant differences in metabolic processes between cancer cells and their (isogenic) cancer stem cell counterpart in spheroids.97,108 Metabolomics has also been applied to the investigation of physiological systems, such as that of the symbiotic human/microbiome axis. For example, Jansen et al. investigated the sensing and signaling of gut/microbiome metabolites in a 3D kidney tissue model. 109 As an extension of metabolomic profiling, an isotope-resolved branch of metabolomics that traces an isotope-labeled metabolite (fluxomics) has assisted in the investigation of specific metabolomic pathways and has identified intracellular metabolic fluxes in 3D disease models. 110 For example, several groups have used fluxomics to investigate metabolic reprogramming 111 or drug response 102 of cancer spheroids.
A limitation of the described studies is that bulk metabolite collection does not allow for spatially resolved metabolomics and does not capture the heterogeneity within the 3D disease model. To address this, our group recently established a 3D culture system that can be rapidly disassembled for the efficient collection of the unstable metabolites to investigate metabolic profiles within graded microenvironments. Using our TRACER system, we employed mass spectrometry to correlate cell consumption-driven gradients of hypoxia to metabolic signatures. 112 To our knowledge, TRACER is the only model that offers rapid sample collection for the purpose of performing spatially resolved metabolomics. Other approaches to spatially resolve metabolomics in 3D disease models include mass spectrometry imaging (MSI), which has been used to spatially map metabolic signatures to investigate response to chemotherapy 113 and to indicate regions of hypoxia and oxidative stress 114 in multicellular spheroids. The collection of spatial information, either through designing inventive 3D disease model architecture or by applying MSI technology, will further exploit the use of metabolomics in 3D disease models.
Lipidomics technologies for 3D in vitro disease models
Lipidomics studies are used to investigate lipid species that are involved in metabolomic processes, associated to signaling pathways or integral to nuclear and cellular membrane structures. Although not extensively utilized in the context of 3D disease models, lipidomics has demonstrated potential to address various questions relevant to 3D systems. Similar to proteomics, lipidomics has been used as a strategy to compare 3D disease models to traditional monolayers as well as to patient samples. For example, mass spectrometry was employed to demonstrate significant differences between 3D disease models and 2D monolayers, 115 while it has identified similar lipid profiles in 3D disease models compared with those observed in diseased organ in vivo.116,117 Several lipidomics studies in the context of relevant 3D disease models have also addressed questions regarding disease progression, drug response, and therapy resistance. For example, lipidomics has been used to investigate the disease mechanism and progression of conditions directly related to a dysfunctional lipid homeostasis such as hepatic steatosis 117 or acne vulgaris. 116 Lipidomics has also shown to be a valuable tool to investigate the effect drug treatment on lipid metabolism, 118 and the effect of altered lipid metabolism on therapy resistance in physiologically relevant 3D tumor models. 119
Emerging in the field of lipidomics are spatially resolved approaches that provide information on cellular lipid function. For example, lipidomics with mass spectrometry in combination with confocal Raman microscopy has spatially mapped lipid species within the 3D disease model and correlated these lipid profiles to cell malignancy. 115 To further improve sensitivity and resolution, another study combined confocal Raman microscopy with matrix-assisted laser desorption/ionization mass spectrometry (MALDI-MS) imaging in which a full mass spectrum is generated from each MALDI laser position to resolve metabolite and lipid species. 120 These developments in spatially resolved lipidomics in 3D disease models are of great value to improve our understanding of the broad range of lipid functionalities in the context of disease.
Multiomics technologies and integrated modalities for 3D in vitro disease models
Multiomics involves extracting a variety of information by performing multiple omics technologies in a single study, and has been shown to provide complementary information in validating, characterizing and discovering new biology with 3D disease models.121,122 In the most developed tissue models, such as cancer organoids, transcriptomics and genomics have been performed together to validate the genetic mutations and transcriptional landscape of the 3D disease model with the patient tissue.28,32,35,52,54,55,123 Other combinations shown in Table 1 include transcriptomics and epigenomics to link epigenetic regulators and disease progression,43,47 transcriptomics and proteomics66,74 to understand disease mechanisms, and even a combination of all omics types to perform a toxicity screen. 46 In contrast to multiomics, integrated modalities simultaneously perform two or more omics technologies, such as transcriptomics and proteomics (Cellular Indexing of Transcriptomes and Epitopes by sequencing [CITE-seq]) 27 or transcriptomics and epigenomics (single-cell Chromatin Overall Omic-scale Landscape sequencing [scCOOL-seq]). 124 However, integrated modalities have not been used yet to investigate 3D in vitro disease models despite its high potential for improving our understanding of disease.
Experimental Design Considerations and Challenges of Omics Technologies and 3D In Vitro Models
Factors to consider when designing omics experiments to obtain meaningful data
There are additional factors and challenges that need to be considered when using omics assays with 3D disease models, on top of those typically accounted for in 2D in vitro systems. This will be broadly discussed here across omics disciplines. The overarching challenge unique to 3D disease models is the isolation of cellular components required for the respective omics technology and corresponding biological question, to obtain a representative dataset. Across 3D disease models mentioned in Table 1, the first step is to remove cells from the 3D disease model, through dissociation and digestion protocols. Spheroids and organoids are regularly harvested, digested, and RNA or DNA molecules are extracted using kit reagents. Other common reagents include TRIzol, TrypLE (typically for Matrigel), and a range of enzymes such as trypsin, proteases, and collagenases.32,51 Selecting the optimal dissociation and digestion protocol relies on balancing a high cell yield and cell viability. For further in-depth discussion on such protocols, readers are referred to Worthington Biochemical Corporation's Cell Isolation Optimizing System manual. 140 For more complex and “novel” 3D systems, specific steps must be performed. This may involve mechanical dissociation by means of tearing the nonbiological component of a 3D cellulosic sponge system to release cells, 126 detaching cells from membranes of a microfluidic system using a plastic scrapper, 105 and pipetting through a modified pipet tip. 45
Components in 3D tissue models, such as nonbiological scaffolds, extracellular matrix (ECM), cell-specific properties, and the rarity of cell populations can also complicate obtaining a complete starting sample. For example, cell adhesion to substrates used to construct the tissue, such as paper scaffolds and cellular sponges, will complicate the digestion protocol. Certain phenomena such as fibrosis, 75 migration, and invasion 76 have been shown to require an ECM to sufficiently mimic in vivo states. As such, the ECM may present an additional physical barrier to properly access cells. Moreover, the cells used in the 3D disease model can be derived from robust cell lines as well as from primary patient samples, which presents another parameter to consider. Although the availability of primary patient samples has been greatly improved by the implementation of living biobanks, these populations are not extensive and often do not include patient-matched samples of cell types essential to disease progression, such as supporting cells in the tumor microenvironment. 141 In these cases, cells that have not previously been expanded will likely be limited in supply, thereby decreasing initial sample quantity. In addition, for analyses of individual cell types within multiculture systems, an extra step to sort cells through fluorescence-activated cell sorting is required. However, this is conducive to losing fragile and sparse samples, and potentially results in the enrichment of more robust cells. The take-home message is that the optimal digestion and dissociation protocol making use of specific enzymes, digestion times, and shaking speeds, will differ across systems, and often need to be systemically designed and optimized for each specific model.
Barriers and challenges associated with omics
Tissue engineers experience several barriers that come with integrating omics technologies with 3D in vitro models. First, limited access to equipment and facilities can be an obstacle. Not all institutions are equipped with omics-related facilities and seeking these out through interinstitution or international contacts can be logistically difficult. Second, the omics experiment, analysis, and facility fees can be costly, especially for novel techniques that have not been widely adopted yet. For this reason, it is important for the researcher to fully understand the purpose of using the omics approach and the significance of the question that is being addressed. Thirdly, the difficulty to obtain bioinformatics expertise for analyzing the large amounts of raw data provides a barrier to entry. Training is usually required to obtain useful information from complicated omics datasets and the selection of appropriate software and tools is not standardized. 121 To get started with analysis of omics datasets, we recommend looking into popular tools that comprehensively describe approaches for data analysis and have been extensively reviewed by others (Supplementary Table S2). However, as omics technologies gain popularity and, arguably, necessity in discovering new biology, there will be a strong demand for institutions to centralize omics facilities and for leaders in the field to standardize omics analysis protocols. With this support, the associated barriers may dissolve and render omics technologies significantly more accessible for tissue engineers.
In addition to these logistical barriers to implement omics technologies, teams developing 3D disease models must prioritize functionally validating the biological relevance of their models. This provides confidence that hypotheses or mechanisms derived from omics analyses are relevant to the in vivo phenomenon being modeled. Such model validation is nontrivial as this ideally requires the assay to provide a functional readout of the 3D disease model that can be directly mapped to an in vivo tissue functional property, such as drug resistance or toxicity. Datasets that enable this functional validation are available for only a few disease models such as cardiac toxicity, 142 cancer,35,68 and cystic fibrosis. 143 Omics technologies have provided mechanisms to assess the similarity of the disease model to patient samples. However, without a functional assay, the importance of any observed gene differences, and whether they render the model predictive or not, is challenging to assess. Partnering with clinicians to link relevant clinical datasets and readouts to validate model predictivity in disease models is therefore, an important goal for the future.
Conclusion: Perspective on Omics Technologies for Future Applications in 3D In Vitro Models
The combination of tissue models and omics technologies is emerging as a powerful tool to benchmark models to clinical samples, identify new targets and understand disease mechanisms. As adoption of complex, spatial, and multiomics approaches is still limited and the field is early in development, there are a number of technical and logistical barriers that make integrating these technologies difficult. Currently, there are also challenges associated with functional validation of the model before assessing with omics technologies. However, the potential of combining 3D in vitro disease models with omics to understand disease more deeply is extremely promising and will provide the opportunity to better link genotype to phenotype in multicellular complex systems.
Footnotes
Acknowledgments
The authors would like to acknowledge Michail Paraskevopoulos for contributions to the literature search, and Ziting (Judy) Xia and Margaret Ho for contributions to resources included in this article.
Disclosure Statement
The authors have no conflicts of interests to declare.
Funding Information
This work was funded by a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery grant to A.P.M. (Fund #RGPIN-2019-06486) and an NSERC CREATE Fellowship to NW.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.