
Abstract
The lack of appropriate experimental models often limits our ability to investigate the establishment of infections in specific tissues. To reproduce the structural and spatial organization of vaginal mucosae to study human immunodeficiency virus type-1 (HIV-1) infection, we used the self-assembly technique to bioengineer tridimensional vaginal mucosae using human cells extracted from HIV-1-negative healthy pre- and postmenopausal donors. We produced a stroma, free of exogenous material, that can be adapted to generate near-to-native vaginal tissue with the best complexity obtained with seeded epithelial cells on the organ-specific stroma. The autologous engineered tissues had mechanical properties close to native mucosa and shared similar glycogen production, which declined in reconstructed tissues of the postmenopausal donor. The in vitro-engineered tissues were also rendered immune competent by adding human monocyte-derived macrophages (MDMs) on the epithelium or in the stroma layers. The model was infected with HIV-1, and viral replication and transcytosis were observed when immunocompetent reconstructed vaginal mucosa tissue has incorporated MDMs into the stroma and infected with free HIV-1 green fluorescent protein (GFP) viral particles. These data illustrate a natural permissiveness of immunocompetent untransformed human vaginal mucosae to HIV-1 infection. This model offers a physiological tool to explore viral load, HIV-1 transmission in an environment that may contribute to the virus propagation, and new antiviral treatments in vitro.
Impact statement
This study introduces an innovative immunocompetent three-dimensional human organ-specific vaginal mucosa free of exogenous material for in vitro modeling of human immunodeficiency virus type-1 (HIV-1) infection. The proposed model is histologically close to native tissue, especially by presenting glycogen accumulation in the epithelium's superficial cells, responsive to estrogen, and able to sustain a monocyte-derived macrophage population infected or not by HIV-1 during ∼2 months.
Introduction
In 2018
Over 80% of new HIV infections occur through sexual transmission, 9 mainly involving vaginal or rectal mucosae. Many studies have characterized HIV-1 capability of crossing the mucosal barriers and establishing infection in the FRT. It was initially suggested that the lower FRT epithelium's numerous layers act as an inaccessible barrier to viruses and other microbes.10,11 In contrast, the endocervix, which constitutes the upper FRT, lacks a thick barrier and is lined only by a single layer of cuboidal epithelial cells, which may be a more susceptible site for HIV-1 penetration and transmission. 12
Nevertheless, many factors pointed out the ability of HIV-1 to pass through the vaginal epithelial barrier: first, its large surface area. Vagina surface area is 15 times larger than the upper FRT. 10 Second, there is a decrease in thickness of the epithelium during the hormonal cycle13,14 or contraceptive use. 15 Third, sex hormones influence vaginal mucosa immunity.16,17 Finally, HIV-1 or simian immunodeficiency virus infections occur in hysterectomized women and hysterectomized macaques, both lacking a cervix.12,18
The semen contains both high viral loads and the principal HIV-1 target cells (i.e., CD4+ T lymphocytes and macrophages) that facilitate the transmission of infection during sexual exposure.19,20 Using in vitro mucosal models, it was shown that HIV-1-infected cells transmit the virus across the epithelial barrier. If the epithelium's surface is intact, in that case, the virus translocates by transcytosis, 21 or can be directly taken up by local target cells such as Langerhans cells, macrophages, and intraepithelial T cells. 22
Macrophages are of particular interest for the vaginal infection route because HIV better replicate in vaginal macrophages compared with intestinal macrophages. 23 After the dissemination phase, they are a source of plasma-circulating virus. 24 Contrarily to CD4+ T cells, macrophages can store virions in their cytoplasm for an extended period without cell death, and then can also constitute a reservoir for HIV-1.25–27
In the search for an adequate model to study vaginal HIV-1 transmission, various models are used: monolayer cell cultures, three-dimensional (3D) models, or animal models. Monolayer cultures of human vaginal or cervical cell lines lack the complex 3D structure, therefore cell differentiation and organization and the variety of cell types present within the vaginal epithelium. Among animal models, nonhuman primates such as pig-tailed macaques 28 and sheeps, 29 or humanized mice30–32 are the most often described. Nevertheless, their use for studying vaginal diseases and treatment applications has scientific, ethical, and economic limitations. 33
Except for nonhuman primates, animal models are limited in simulating the human vaginal infection and inflammatory response due to the differences in genital tract physiology, pH, microflora composition, and anatomy. 34 3D models constructed from human or porcine explanted tissue35,36 or grown from primary human cells, including 37 or excluding38–42 immune cells, have the potential to more closely mimic human vaginal epithelium than do the in vivo animal models. Although these models recapitulate essential aspects of HIV-1 pathogenesis close to physiological settings, they have certain limitations. These include the unavailability of enough tissues for large-scale experiments, incorporation of exogenous membrane and biomaterials, insufficient mechanical properties for in vivo studies, lack of organ-specific stroma and the submucosal compartment, and in some cases, lack of fully differentiated epithelial layers. The use of an exogenous matrix may also influence susceptibility to HIV-1 infection and transmission.
Therefore, we aimed in this study to elaborate and validate a 3D human cell culture model that mimics female genital mucosae. To reproduce as much as possible the invasion by HIV-1 of the vaginal mucosa, this model has been derived from a reconstructed human 3D organ-specific vaginal bilayer mucosa.43,44 The model should be at the minimum histologically close to native tissue, hormone responsive, offer mechanical resistance to be used in vitro, and able to sustain a macrophage population infected or not by HIV-1.
Materials and Methods
Ethics statement
All procedures involving patients were conducted according to the Helsinki Declaration and were approved by the Research Ethical Committee of CHU de Québec-Université Laval. Donors' consent for tissue harvesting was obtained for each specimen. Experimental procedures were performed in compliance with the CHU de Québec guidelines.
Cell lines
Human VK2 vaginal epithelial HPV-16 E6/E7 transformed cells (ATCC CRL-2616) were purchased and cultured according to ATCC guidelines. Human vaginal stromal and epithelial cells were isolated from vaginal biopsies of three HIV-1 negative females of different ages undergoing urological surgeries to manage a urogenital sinus anomaly (3 years old patient), a vaginal septum (32 years old), or a cystocele (64 years old). The vaginal biopsy was cut into small pieces and subjected to 0.5 mg/mL thermolysin (Sigma, St-Louis, MO) solution containing 1 mM CaCl2 (Sigma) and Hepes pH 7.4. 45 The epithelium was manually separated from the stroma. The vaginal epithelium was harvested using trypsin/ethylenediaminetetraacetic acid (EDTA) solution (0.05% trypsin; Intergen Company, Purchase, NY; 0.01% EDTA/disodium salt, J.T. Baker, Phillipsburg, NT). The vaginal stroma was digested by collagenase H (Roche Diagnostics, Laval, QC, Canada) to extract stromal cells. Human dermal fibroblast and bladder mesenchymal cells were extracted from human skin and bladder biopsies following the same method used for vaginal cells.44,46
Stromal cells were cultured with Dulbecco-Vogt modified Eagle's medium (DMEM; Invitrogen, Burlington, ON, Canada) and 10% fetal calf serum (Hyclone, Scarborough, ON, Canada). In contrast, epithelial cells were grown on a feeder layer of irradiated mice fibroblasts S3T3 in DMEM supplemented with HAM's F12 in a 3:1 ratio (DMEM HAM, Invitrogen), with 10% fetal calf serum, 0.4 μg/mL hydrocortisone (Calbiochem, La Jolla, CA), 5 μg/mL insulin (Sigma), 1 nM cholera toxin (Sigma), and 10 ng/mL epidermal growth factor (Austral, San Ramon, CA). Both media were also supplemented with antibiotics (penicillin 100 UI/mL and gentamicin 25 μg/mL) to avoid bacterial contamination. Cells were expanded with a change of media every 2–3 days and frozen until use.
Human peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Hypaque gradient from a healthy donor's blood samples. These cells were seeded in flasks supplemented with 10% human serum type AB (Valley Biomedical, Winchester, VA) to allow monocyte adherence for 2 h, and then washed to remove unattached PBMCs. Cultures were complemented with 25 ng/mL macrophage-colony stimulating factor (M-CSF; Genscript, Piscataway Township, NJ) for 3–4 days to allow differentiation into monocyte-derived macrophages (MDMs). Cells were then kept in culture for three additional days in a complete culture medium (without M-CSF) to obtain nonpolarized MDMs. After 6 days of differentiation, MDMs were treated with Accutase (Invitrogen, ThermoFisher Scientific) for 1 h and detached with a soft cell scraper (Sarstedt, Nümbrecht, Germany). Cells were plated for 24 h in culture medium before virus infection in ultralow attachment plates (Corning).
3D tissue generation
The self-assembly technique exploited fibroblasts' inherent characteristics to produce an extracellular matrix (ECM) supplemented with 50 μg/mL ascorbic acid. This vitamin C derivate promotes collagen protein synthesis and deposition of sulfated glycosaminoglycans.47–49 Once fibroblasts are supplemented with ascorbic acid for 21–35 days, they form matrix sheets where stromal cells are embedded. After seeding epithelial cells on one of the sheets, the layers are peeled from the culture dish, superimposed, and maintained at an air/liquid interface for an additional 3 weeks for further cell–matrix reorganization, layer fusion, and epithelial cell apical differentiation. Estrogen was added to optimize the culture, and ensure enough hormonal levels to support the proliferation and maturation of the vaginal compartments and an adequate glycogen supply. Ten nanometer estrogen (β-estradiol E2758 Sigma-Aldrich) was used during stroma culture, epithelium maturation only, or throughout the culture of stroma and epithelium cells.
Tissue characterization, histology, and microscopy
The mechanical properties of engineered tissues were measured by uniaxial tensile testing using an ElectroPulse E1000 mechanical tester (Instron, Norwood, MA). Instrument-specific biopsies were dissected out of the tissue with a punch, and both extremities of the sample were stretched at a constant rate of 0.2 mm/s until the tissue ruptured. Tissue thickness, well-differentiated epithelium, and enhanced glycogen reservoirs were evaluated by histology and immunofluorescence (IF) analyses from either formalin-fixed paraffin-embedded blocks or fresh tissue frozen in optimal cutting temperature (Sakura, Finetek, Torrance, CA). Masson's trichrome (MT) staining was used for general tissue architecture, cellular organization, morphology assessment, and stroma versus epithelium measurements. Richard-Allan Scientific™ periodic acid-Schiff (PAS) (ThermoFisher Scientific; 87007) staining was used to evaluate the polysaccharides such as glycogen, glycoproteins, glycolipids, and mucin content in the epithelium. Polysaccharides stained bright magenta with this technique. To confirm that the depots were attributed to glycogen and no other glycoproteins, glycolipids, and mucins, we used α-amylase that acts on glycogen by digesting it out, followed by PAS staining. 50 IF staining to visualize the epithelium or the stroma was performed using antibodies that recognized the epithelial cells, cytokeratin 13 (K13, clone K812 IgG1 mouse 1/20). Secondary antibodies coupled with a fluorochrome Donkey antimouse Ig (clone A21203 Alexa 594 1/100); IgG Alexa 488 (Life Technologies 1/500); or isotype control IgG1 (Dako X093 1/100) were used. Hoechst 33258 5 mg/mL (Sigma-Aldrich) was added to counterstain the nuclei. Samples were observed with a Zeiss Axio Imager M2 microscope equipped with an AxioCam HR Rev3 camera (Oberkochen, Germany). Images were processed with the AxioVision 40 V4.8.2.0software (Carl Zeiss), and scale bars were added with ImageJ software (NIH, Bethesda, MD).
Incorporation of MDMs into the engineered vaginal tissues
MDMs were either incorporated into the stroma between two sheets at 5 × 103 cells per layer before the final stacking or on top of the differentiated epithelium at 2 × 105 cells per tissue. Antibodies mouse antihuman CD163 clone 2G12 IgG1 (1/100) was used to visualize the MDMs by IF. The MDMs were either mock-infected or infected with HIV-1.
p24 antibody capture assay
The measurement of virus-encoded p24 protein was determined by an in-house enzymatic assay. 51 In brief, flat-bottomed 96-well plates (Immunol 2; Dynatech, Ltd.) were initially coated with 183 H12–5C, a monoclonal anti-p24 antibody. After the wells were washed and blocked with 1% bovine serum albumin (Sigma), viral lysates were added to the wells at various dilutions, along with samples of known p24 concentration (recombinant purified p25gag/SF2, supplied by Chiron Corporation), to establish a standard curve. After a 60-min incubation at 37°C, the plates were washed, and a second biotinylated anti-p24 monoclonal antibody (i.e., clone 31-90-25) was then added. After a 1-h incubation at 37°C, the plates were washed. A streptavidin–peroxidase conjugate (Streptavidin-HRP-40; Research Diagnostics, Inc., Flanders, NJ) was added; this was followed by the addition of the TMB-S substrate (Research Diagnostics, Inc.). After 30 min at room temperature, the reaction was terminated by adding 1 M H3PO4, and the absorbance was measured at 450 nm. Unknown p24 values were calculated based on regression analysis of p24 standards over a linear range of 31.25 to 2000 pg/mL. To eliminate free p24, before capture assay, each supernatant was ultrafiltered in Centricon® Plus-20 Biomax-100 filter devices (Millipore Corporation).
Viral stock preparation
Viruses were produced by calcium-phosphate transfection of HEK293T cells with the NL4.3 eGFP-IRES-Nef Bal env molecular construct, a replication-competent R5-tropic virus construct into which the reporter gene enhanced GFP (eGFP) was inserted in front of an IRES sequence and the Nef gene. 52 After 2 days, the cell-free supernatant was filtered and ultracentrifuged at 100,000 g for 1 h in an Optima L-90K ultracentrifuge (Beckman Coulter). Viral pellets were resuspended in endotoxin-free Dulbecco's phosphate-buffered saline (PBS). Infected cells could be monitored by GFP detection. The viral preparation p24 content was measured with a homemade enzyme-linked immunosorbent assay (ELISA) specific for the HIV-1 capsid antigen p24. 51 Mucosal tissue equivalents were infected with cell-free viruses (50 ng of p24) in 25 μL PBS for 4 h. Experiments with infected MDMs were performed using 2 × 105 MDMs exposed to 90 ng of p24 for 5 days, harvested, washed, and then seeded apically on the tissue. All mock or infected tissues were fixed at 24–72 h postinfection in 2% paraformaldehyde for 24 h.
Luciferase assays
This assay was used to evaluate the replicability of the virions produced in the 3D vaginal mucosa model. One hundred microliters of cell culture supernatant was transferred into 2 × 105 TZM-bl indicator cell culture for 48 h. This cell line is a genetically modified HeLa-derived cell line expressing large amounts of cell surface CD4, CCR5, and C-X-C chemokine receptor type 4 (CXCR4). 53 These cells are available through the NIH AIDS Reagent Program, Division of AIDS NIAID, NIH (catalogue no. 8129), and carry separate integrated copies of the luciferase and β-galactosidase genes under the control of the HIV-1 promoter, which makes them highly susceptible to infection with different HIV-1 variants (both R5 and X4 tropic). Luciferase activity was measured according to the manufacturer's protocol using the luciferase assay system (Promega, Madison, WI) in a microplate luminometer (MLX; Dynex Technologies, Chantilly, VA). Luciferase activity is expressed in relative light unit (RLU).
Results
The vaginal mucosa model reconstructed with organ-specific stromal and epithelial cells presented the closest histological features to native tissue
The vaginal mucosa model was first reconstructed using transformed cells VK2, commonly used in HIV-1 infection studies. A MT was performed on a slide of reconstructed tissue. Compared with native tissue, models reconstructed using VK2 cells, whatever the origin of stromal cells (dermal or vaginal fibroblast), presented an unorganized stacking of epithelial cells. Since VK2 did not allow obtaining an adequately differentiated epithelium, primary organ-specific epithelial cells (EvaX32) were tested on the different stroma. Reconstructed dermis and bladder lamina propria failed to produce a convincing epithelium. Only the combination of vaginal fibroblasts to reconstruct the stroma and untransformed vaginal epithelial cells allowed the production of an adequately differentiated vaginal mucosa showing basal cells, intermediate cells, and empty cells on the luminal side (Fig. 1).
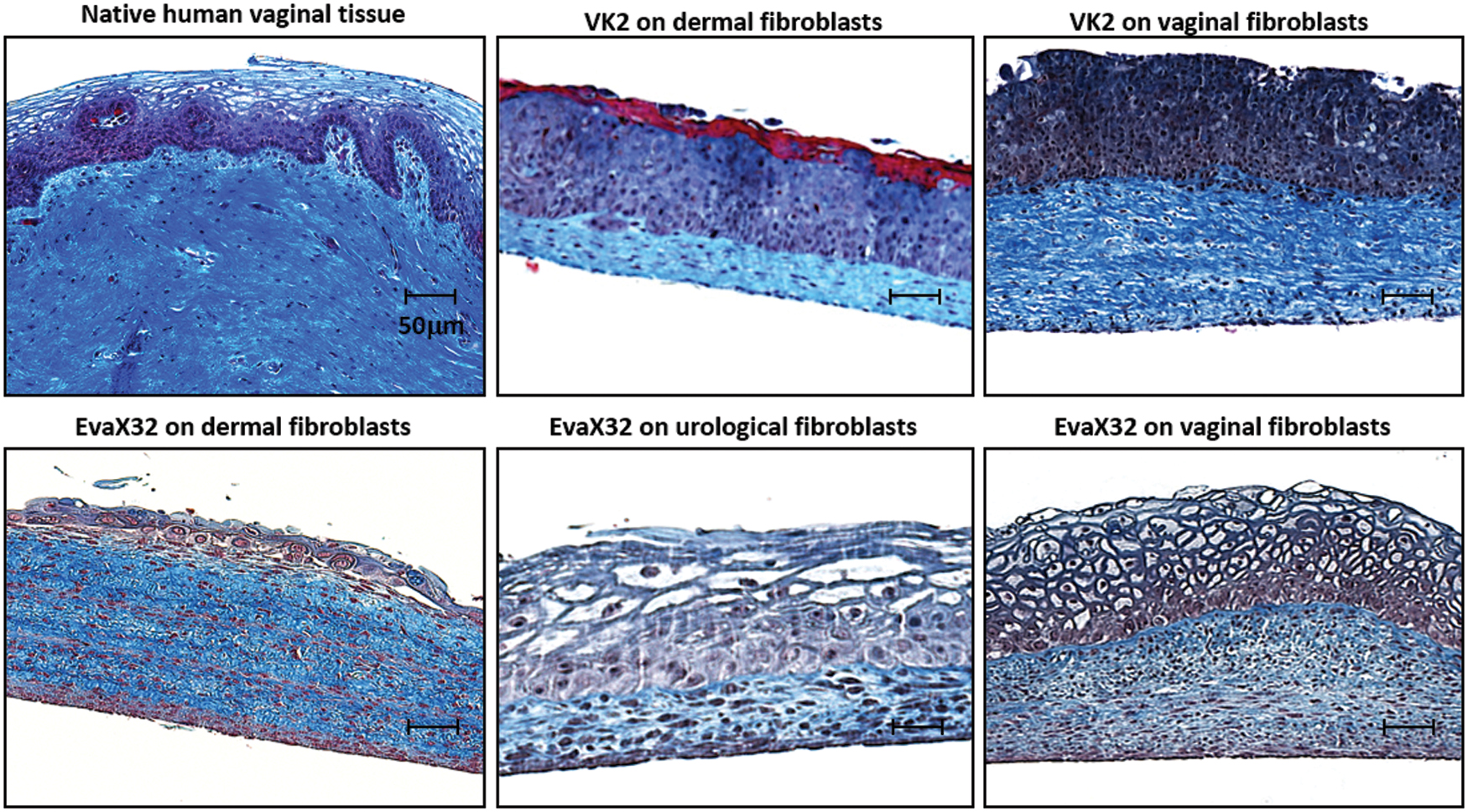
Self-assembly reconstruction of near-to-native vaginal mucosa tissue using organ-specific human untransformed fibroblast and epithelial cells. Native human vaginal cross-section analyzed by MT staining ( × 100) is compared with in vitro reconstructed tissue using epithelial HPV E6/E7 transformed vaginal epithelial VK2 cells seeded on human dermal fibroblast layers or human vaginal fibroblast layers (top). Healthy vaginal epithelial cells extracted from a 32-year-old donor (EVaX32) were seeded on human dermal fibroblasts, human bladder fibroblasts, or vaginal fibroblasts from the same 32-year-old donor (bottom), to illustrate the crosstalk between organ-specific fibroblasts and epithelial cells. N > 5; scale bar: 50 μm. MT, Masson's trichrome. Color images are available online.
The maturation of the vaginal epithelium with an adequate glycogen supply is ensured by organ-specific crosstalk between the stroma and the epithelium
To verify if the empty cells on the luminal side observed using a MT staining were fully differentiated glycogen-storing cells, a PAS staining was performed. Half of the slides were treated with amylase, which digests glycogen to ensure the specificity of staining. α-Amylase treatment removed the magenta granules, therefore, likely to be glycogen and no other polysaccharides. As it was observed in native tissues, the PAS staining confirmed the maturation of the vaginal epithelium. Enhanced glycogen reservoirs were observed on the luminal side of the reconstructed tissue only when vaginal fibroblasts were used to reconstruct the stroma, allowing organ-specific crosstalk between the stroma and the epithelium (Fig. 2).
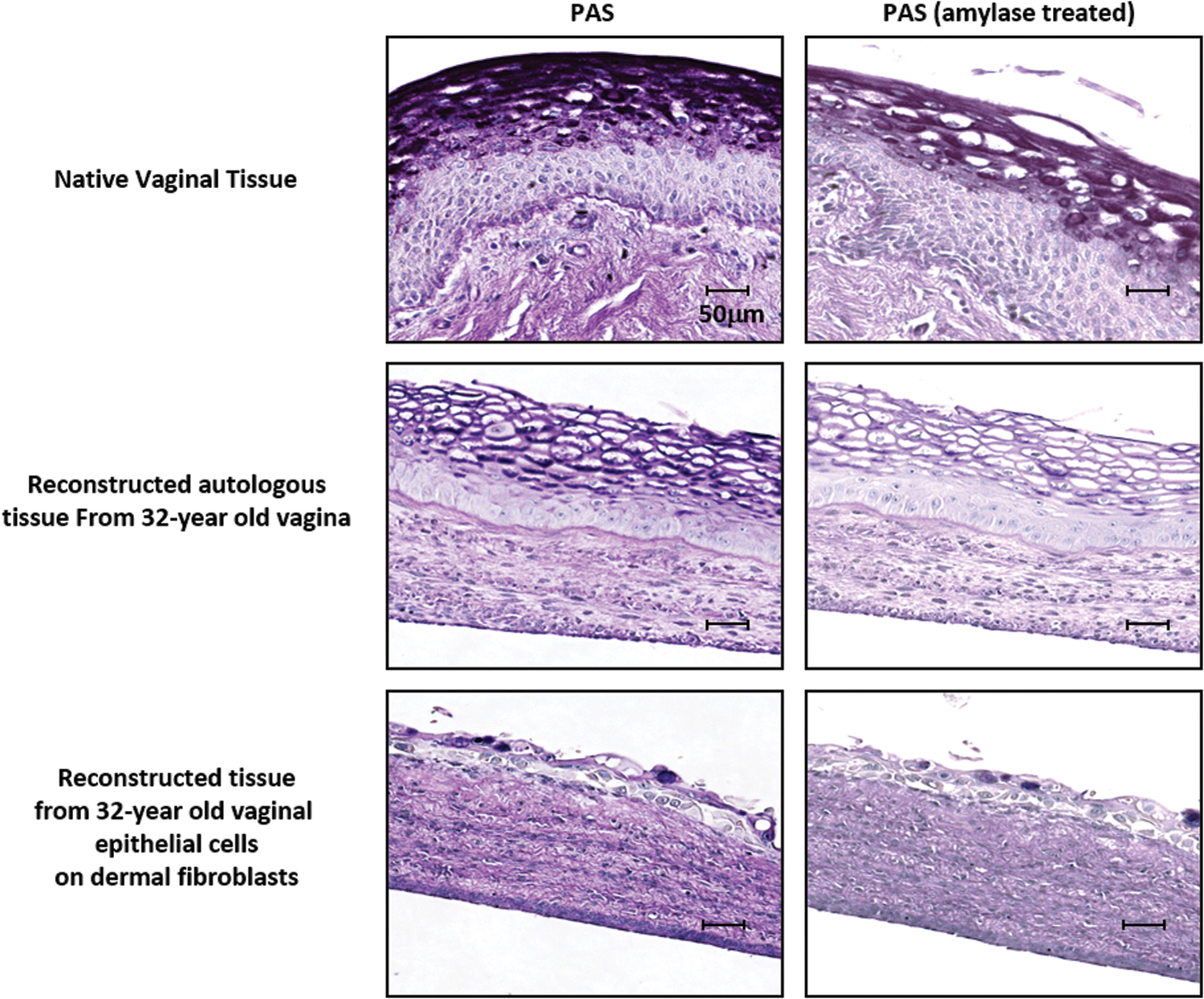
The proliferation and maturation of the vaginal epithelium and an adequate glycogen supply are ensured by organ-specific crosstalk between the stroma and the epithelium. Vaginal reconstructed tissues using dermal stroma cells or vaginal stroma cells from a 32-year-old donor were untreated or treated with α-amylase that acts on glycogen by digesting it out, followed by PAS staining. Polysaccharides such as glycogen, glycoproteins, glycolipids, and mucin content in the epithelium were stained bright magenta using the PAS technique, which becomes very light post-α-amylase. N = 3 in triplicate for each experiment. Scale bar: 50 μm. PAS, periodic acid-Schiff. Color images are available online.
Reconstructed 3D vaginal mucosa tissues share similar native tissue architecture and glycogen production, depending on the donor's age
In the previous experiment, only cells from a vaginal biopsy of a 32-year-old woman were used. To verify the impact of the donor's age on the tissue's architecture and its correlation to what is observed in vivo, cells from a 3-year-old girl, 32-year-old premenopausal woman, and 64-year-old postmenopausal woman were used. MT and PAS staining were performed on the reconstructed tissues. As expected, tissues produced using the cells from 3- and 64-year-old donors presented a thinner epitehlium with lower glycogen-content than those produced using cells from premenopausal women (Fig. 3). Once again, if dermal fibroblasts were used, the epithelium failed to differentiate (Fig. 3, last lane). Nevertheless, the use of labia cells gave an intermediate result more mature than the one lying on a dermis stroma but less mature than the completely organ-specific ones. Also, the presence of keratinization was observed in this case (Fig. 3, second lane).

Reconstructed 3D vaginal mucosa tissues share similar native tissue architecture and glycogen production, which declines with age—using MT and PAS staining. Polysaccharides such as glycogen, glycoproteins, glycolipids, and mucin content in the epithelium were stained bright magenta using the PAS technique. Native human vaginal cross-section ( × 100) was compared with in vitro reconstructed tissue using donor-specific epithelial and stromal cells from various age groups. Reconstructed autologous tissue from 3-year-old labia and vaginal biopsy were analyzed for the general architecture of the stroma and epithelial cell differentiation, and compared with tissue extracted from a premenopausal 32-year-old donor and postmenopausal 64-year-old donor. The importance of the organ-specific crosstalk between fibroblasts and epithelial cells was illustrated using EvaX32 seeded on human dermal fibroblasts. N > 5; scale bar: 50 μm. 3D, three dimensional. Color images are available online.
Reconstructed 3D vaginal mucosa tissues from premenopausal donors showed good mechanical properties for in vitro use
The reference values for native vaginal tissue are 5.5 Mpa and 0.5 N for elastic modulus and maximal strength, respectively. 43 Nevertheless, a graft of urological tissues presenting maximal strength ∼0.3 N was successful. Tissues could be easily used for mechanical or permeation tests with a maximal strength ∼0.1 N. All the reconstructed tissues had maximal strength ≥0.2 N with an elastic modulus inferior to the reference value (then more elastic). They could be easily handled in the laboratory. Maximal strength values and elastic modulus were obtained with tissues from the 32-year-old donor [Fig. 4 (1–3)]. For comparison, reconstructed tissues using dermal fibroblasts showed better maximal strength but lacked the elasticity of native vaginal mucosa [Fig. 4 (4)]. It also had the thickest stroma but the less differentiated epithelium, as seen in Figures 1–3.
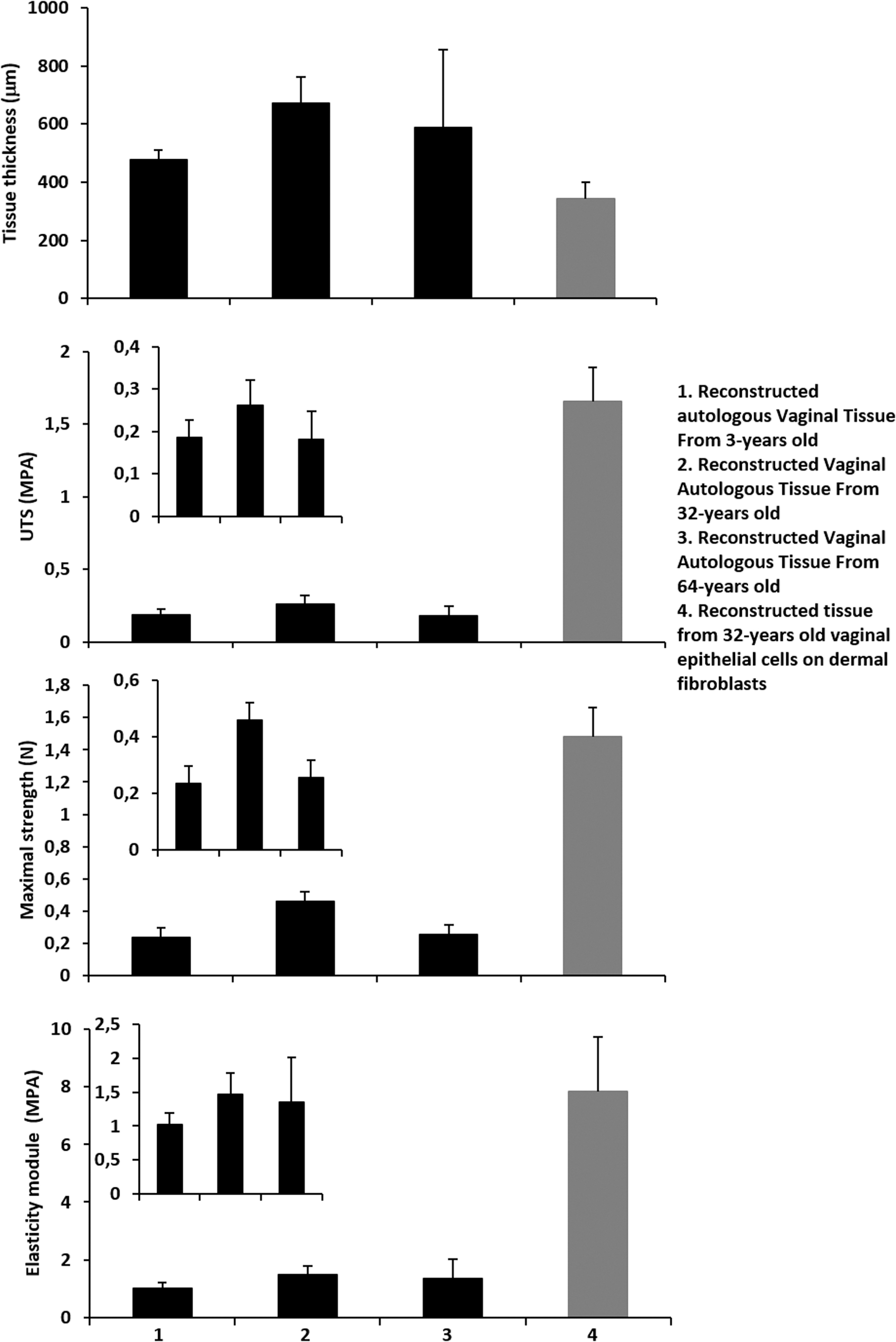
Reconstructed 3D vaginal mucosa tissues from premenopausal donors show optimal mechanical properties for in vitro use. The mechanical characteristics of (1) reconstructed autologous vaginal tissue from 3-year-old, (2) 32-year-old, (3) 64-year-old, and (4) reconstructed tissue from 32-year-old vaginal epithelial cells on dermal fibroblasts were evaluated for the thickness of tissue (μm), UTS (Mpa), maximal strength (N), and elastic modulus (Mpa). All values were calculated as means ± standard deviation. N = 3 in triplicate for each experiment.
A 10 nM supply of estrogen during the stroma culture provides sufficient hormonal levels to ensure the proliferation and maturation of the vaginal epithelium and an adequate glycogen supply
To evaluate the effect of hormonal supply on the differentiation of the vagina mucosa model at different points of the production process, estrogen was (1) omitted, (2) added during the stroma reconstruction only, (3) added during the air/liquid maturation phase only, or (4) throughout the process. The stroma and epithelium thickness was enhanced when stroma was reconstructed in the presence of estrogen but not when estrogen was added only during the epithelial maturation phase. In contrast, in this culture condition, the epithelium thickness was reduced (Fig. 5). Also, glycogen accumulation was correlated in the same manner.
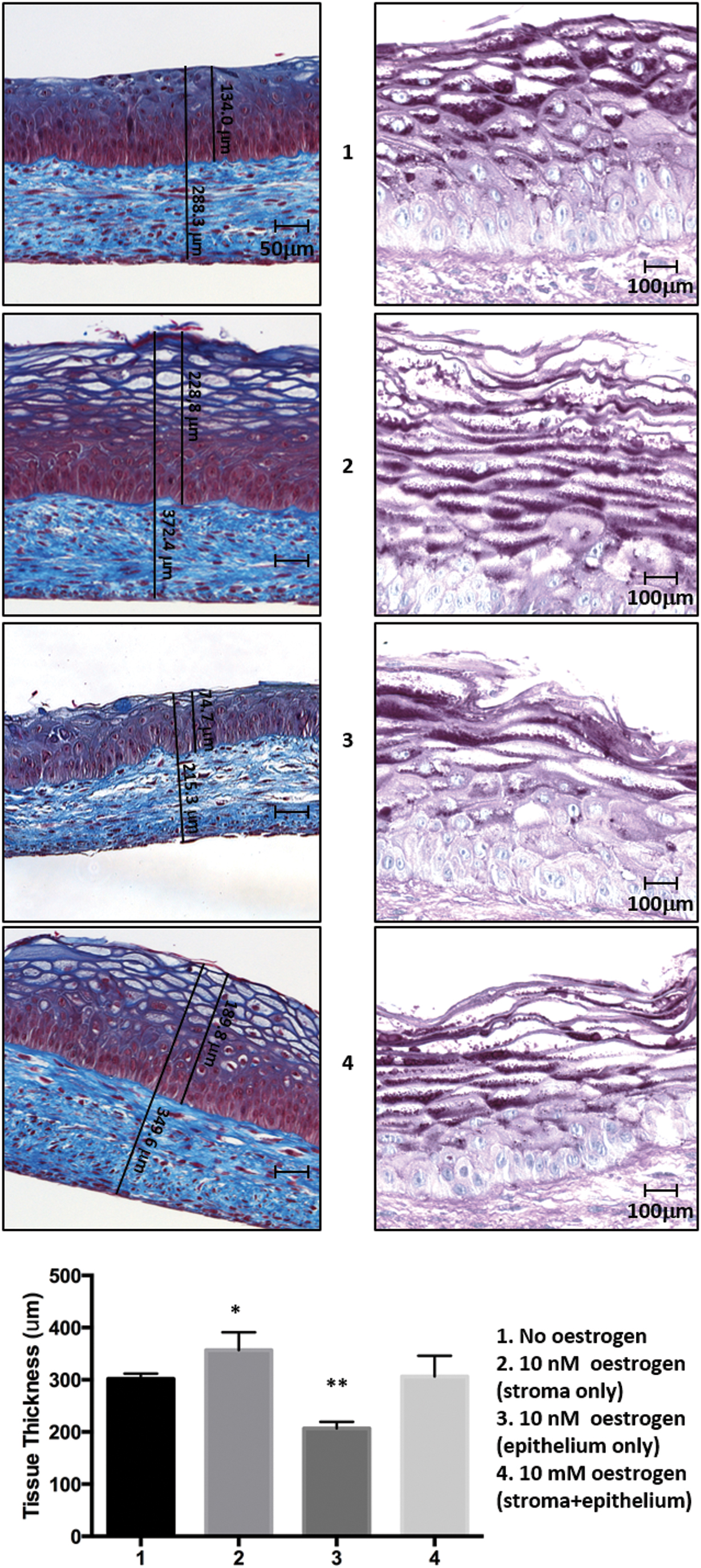
A 10 nM supply of estrogen during the stroma culture provides sufficient hormonal levels to ensure the proliferation and maturation of the vaginal epithelium and an adequate glycogen supply. MT and PAS staining were performed on reconstructed vaginal tissues from the 32-year-old donor. Culture conditions involved (1) no additional estrogen in the media, (2) 10 nM estrogen during stroma culture only, (3) 10 nM estrogen during epithelium maturation only, and (4) ten millimeter estrogen throughout the culture of stroma and epithelium maturation. Stroma thickness, the well-differentiated epithelium (MT), and glycogen accumulation (PAS) were observed. N = 3 in triplicate for each experiment. Scale bar: 50 μm or 100 μm. *p = 0.05; **p = 0.01. Color images are available online.
Estrogen addition had no impact on the tissue mechanical properties
Despite the increasing thickness of stroma when estrogen was added to the cell culture medium (during stroma production only or throughout the process), the mechanical properties of the tissue remained unchanged (Fig. 6).

Estrogen addition has no impact on the tissue mechanical properties. Tissues were evaluated for thickness (μm), UTS (Mpa), maximal strength (N), and elastic modulus (Mpa). Vaginal tissues were reconstructed from the 32-year-old donor cells. Culture conditions involved (1) no additional estrogen in the media, (2) 10 mM estrogen throughout the culture of stroma and epithelium cells, and (3) 10 nM estrogen during stroma culture only. All values were calculated as means ± standard deviation. N = 3 in triplicate for each experiment.
No active viral replication was observed when HIV-1-infected human MDMs were seeded on the apical side of the 3D reconstructed vaginal mucosa tissue
Free HIV-1 particles could not infect the epithelium or stromal compartment in a nonimmunocompetent human 3D vagina model (Supplementary Fig. S1). We, therefore, used HIV-1-infected MDMs seeded apically on human 3D vagina tissues. Contrarily to the mock uninfected condition, while MDMs remained at the tissue surface even after 72 h, infected MDMs (colocalized Alexa 594 and GFP signals) were detected in the stroma compartment after 72 h (Fig. 7A). The epithelial barrier remained intact until 72 h in the mock condition, whereas some degradation of the epithelium integrity could be observed when infected MDMs were seeded. The addition of infected MDMs leads to a disturbance of the basal layer cells after 48 h, which increased again after 72 h, allowing the passage of HIV-1-infected MDMs into the stroma (Fig. 7B). Nevertheless, despite the increase of viral p24 capsid amount in the cell culture supernatant, indicating the MDMs crossed the stroma in a kinetic correlating manner, also observed in IF (Fig. 7C), no viral replication was detected in the supernatant of infected tissues tested on TZM-bl indicator cell culture (Fig. 7D).

No active viral replication is observed when HIV-1-infected human MDMs are seeded on the apical side of the 3D reconstructed vaginal mucosa tissue. 2 × 105 human MDMs were exposed to PBS (U; Mock) or infected (I) with NL4-3 Bal HIV-1 GFP (MOI = 0.1) for 5 days, and then apically added to the reconstructed vaginal tissues from the 32-year-old donor. Double IF analyses (200 × ) of these tissues after 24, 48, or 72 h of incubation were performed using antibodies to specifically stain macrophage, CD163/Alexa 594, and the HIV-1 viral GFP
Viral replication and transcytosis were observed when immunocompetent 3D reconstructed vaginal mucosa tissue had incorporated MDMs into the stroma and was infected with free HIV-1 GFP viral particles
In a different setting, MDMs were first incorporated into the model during the stroma production, and the vaginal mucosa model was then infected using free HIV-1 virus particles. Forty-eight hours after infection, HIV-1 particles had crossed the epithelium and infected the MDMs, as shown by the colocalization of CD163 and GFP signals (Fig. 8A). Measurement of viral p24 capsid by ELISA shows that transcytosis can already be observed at 24 h (Fig. 8B). In this condition, the HIV-1 replication could also be observed by luciferase assay on TZM-bl indicator cell line (Fig. 8C).
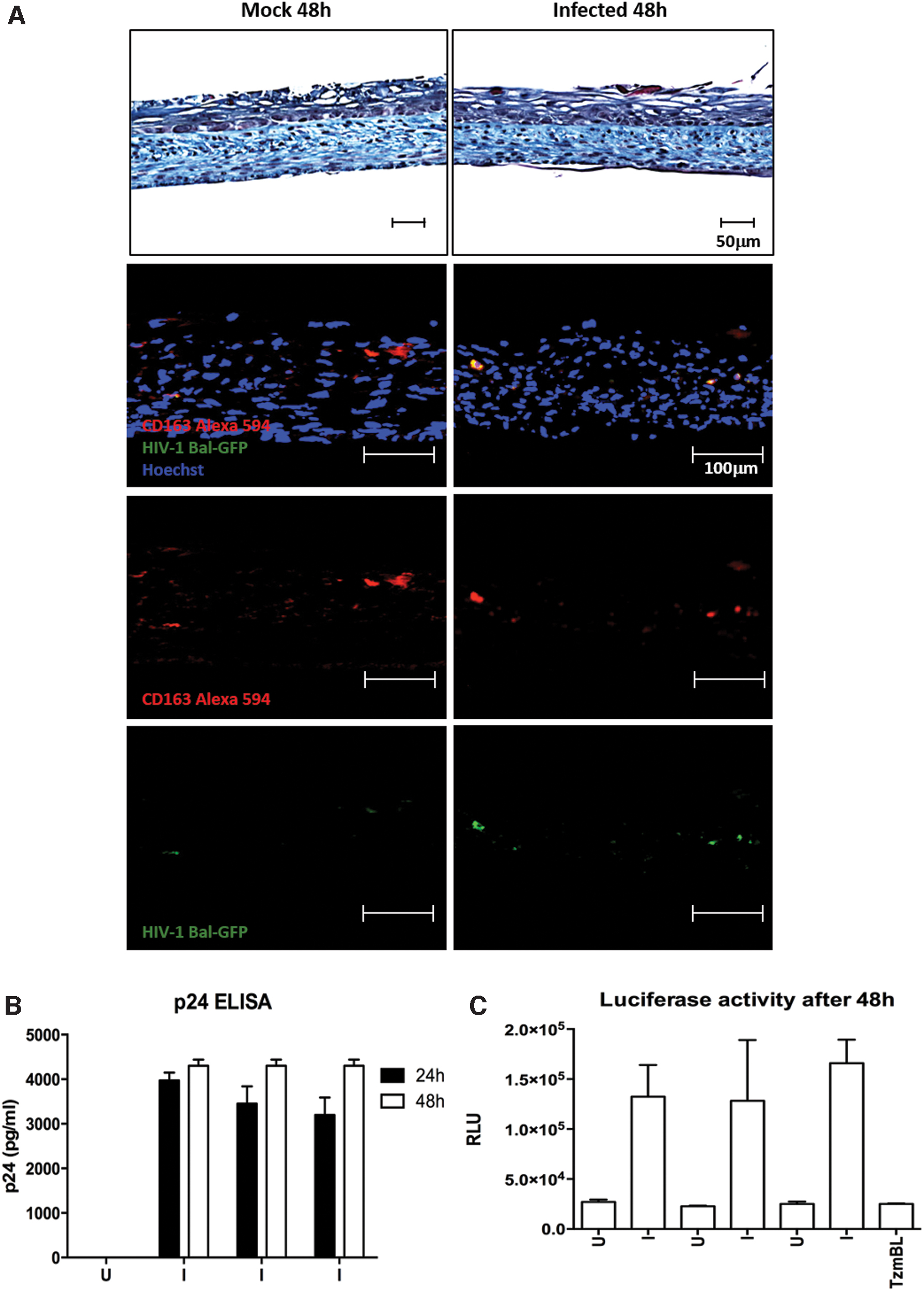
Viral replication and transcytosis are observed when immunocompetent 3D reconstructed vaginal mucosa tissues have incorporated MDMs in the stroma and are infected with free HIV-1 GFP viral particles. Cross-sections of immunocompetent vaginal tissues (100X) exposed to PBS uninfected (U; Mock) or 90 ng of NL4-3 Bal HIV-1 GFP (I) were histologically analyzed with MT staining to confirm intact epithelium and general tissue architecture. Each tissue was engineered with 5 × 103 MDM incorporated into the stroma. Double IF staining of specific macrophage marker CD163 and the HIV-1 viral GFP showed infection of stroma MDMs with HIV-1 GFP particles after 48 h
Discussion
This work aimed to produce a vaginal mucosa model, which could be useful not only to study HIV-1 infection of FRT but also to be a platform to test various drugs. The specific objectives were to obtain a reconstructed model histologically close to native tissue that was hormone responsive, mechanically resistant to being readily used in vitro, and able to sustain a macrophage population infected or not by HIV-1. The model produced using organ-specific stromal and epithelial cells showed the closest histology to the native tissue (Fig. 1) and a better accumulation of glycogen in the tissue, indicating a high degree of epithelial differentiation (Fig. 2). The effect of the donor's hormonal status, prepubescent, premenopausal, or postmenopausal, clearly showed results mimicking the in vivo physiological response (Fig. 3). This result suggested a hormone-responding model, which was confirmed by the decreased thickness of the epithelium when estrogen was omitted in the cell culture medium (Fig. 5). All the models produced, regardless of the donor's age or the presence or absence of estrogen, presented mechanical properties compatible with use in the laboratory, especially for permeation tests (Figs. 4 and 6). The vaginal model could be invaded by HIV-1-infected MDMs or free HIV-1 particles (Figs. 7 and 8).
Nevertheless, in the experimental setting, when HIV-1-infected MDMs were seeded on the epithelium, no viral replication was measured (Fig. 7). In contrast, free HIV-1 impairs quickly mucosal epithelial barrier integrity, 54 and can infect and replicate in stroma-embedded MDM (Fig. 8). The maximum amount of measured viral p24 capsid after 72 h into the cell culture supernatant of the experiment done with pre-infected MDMs is 200 pg/mL (Fig. 7C), whereas 4000 pg/mL of viral p24 capsid (Fig. 8B) were measured 24 h after free HIV-1 infection of the immunocompetent vaginal mucosa model. Very few HIV-1-infected MDMs cross the basal lamina to invade the stroma, whereas many free HIV-1 particles did. The MDMs incorporated into the tissue may be stimulated by stromal cells and ECM components for 2 months. The ones cultivated without contact with the vaginal stromal cell or ECM can present different phenotypes.55–57 It has been reported that mucosal stromal fibroblasts enhance HIV-1 infection. 58 An assay using decellularized stroma could be performed to investigate this point further. It is also noticeable that MDMs remained alive in the tissue for >2 months (the stroma production and the epithelial differentiation phase).
This model could be of interest to investigators wanting to study HIV-1 infection of FRT or test new drugs such as microbicide gels. This model showed an apparent accumulation of glycogen in the superficial cells of the vaginal epithelium. This setting could indicate that an adequate supply of glycogen, a source of nourishment for the vaginal microbiota, including the lactobacilli, can directly impact the modeling of HIV-1 infection in vitro. The bacteria colonization induced a drastic pH change into the vaginal mucus, from basic during the prepubescent phase to acid during the premenopausal phase, to return to basic during the postmenopausal phase. 59 The presence of lactic acid can negatively impact HIV-1 infection.60–62 The production of vaginal microbiota may, in turn, enhance proinflammatory signals mediated by some bacteria and enhance HIV-1 transmission. For example, the proinflammatory milieu can increase CD4+ target cells or damage the epithelial barrier through disruption of cell junction. 63 Also, vaginal lactobacilli induce differentiation of monocytic precursors toward Langerhans-like cells. 64 This relationship between HIV-1 infection and microbiota could be investigated in vitro. It is especially relevant to test microbicide. 65 Our model can then be improved by adding microbiota in the same manner as it was done by Pandit et al. 37 The semen's role, a complex mix of cells and macromolecules, is also discussed for HIV-1 transmission into the vagina. 66 For example, sulfogalactosylglycerolipid interacts with HIV-1 gp120. The alkaline pH of the semen can also be favorable to HIV-1 infection by neutralizing the vagina mucus' acidic pH protecting the virus. 67
We could tailor our model to mimic the menstrual cycle by adding estrogen and progesterone because this hormone (and analogs used for contraception 28 ) correspond to the peak of HIV-1 infection susceptibility. 68 Sex hormones are also related to the vaginal microbiome and vaginal immunity. 16 Because we can control the surface quality of the epithelium and the thickness of our stroma, expanding the culture model to mimic immune-competent mucosae, microbiota, pH, and other conditions could help understand how the response of the FRT is modulated during HIV-1 infection, with or without antiviral, microbicides, or other treatments.
Conclusion
This vaginal mucosa model is histologically close to native tissue, hormone responsive, mechanically resistant, and can sustain a macrophage population infected or not by HIV-1 during ∼2 months. It offers a tool to explore viral load and HIV-1 transmission in an environment that may contribute to propagating the virus and new antiviral treatments in vitro. Future perspectives include its improvement to render it even more physiologically relevant.
Footnotes
Authors' Contributions
I.S., C.B., H.O., M.P., M.J.T. and S.B. conceived and designed the experiments; I.S., C.B., N.R., M.L., W.J. and A.L. performed the experiments; I.S., C.B., S.C., N.R., M.L., W.J. and A.L. analyzed the data; M.J.T. and S.B. supervised the project; I.S., C.B. and S.C. wrote the article; S.C. and S.B. proofread the article; All authors read and approved the final version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the “Fonds de Recherche du Québec—Santé (FRQS) and the MerckSchering-Plough/Université Laval Foundation.” S.B. was a recipient of the Canadian Urological Association Scholarship Fund. The project is also funded by the Quebec Cell, Tissue and Gene Therapy Network—ThéCell (a thematic network supported by the Fonds de recherche du Québec—Santé).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.