
Abstract
Despite their large societal burden, the development of therapeutic treatments for neurodegenerative diseases (NDDs) has been relatively unsuccessful. This is, in part, due to a lack of representative experimental models that reveal fundamental aspects of human brain pathology. Recently, assays for in vitro modeling of the human central nervous system (CNS) have significantly improved with the development of brain and spinal cord organoids. Coupled with induced-pluripotent stem cell and genome editing technologies, CNS organoids are a promising tool for studying neurodegeneration in a patient-specific manner. An extensive array of protocols for the generation of organoids for different brain regions has been developed and used for studying neurodegenerative and other brain diseases. However, their application in the field of motor neuron disease (MND) has been limited due to a lack of adequate organoid models. The development of protocols to derive spinal cord and trunk organoids and progress in the field of assembloids are providing new opportunities for modeling MND. In this study here we review recent advances in the development of CNS organoid models, their application in NDDs, and technical limitations. Finally, we discuss future perspectives for the development of organoid-based systems for MND and provide a framework for their development.
Impact statement
Animal models and two-dimensional cultures are currently the main platforms for studying neurodegenerative diseases (NDDs). However, central nervous system (CNS) organoid technology offers novel possibilities for studying these diseases. Organoid modeling in combination with emerging organ-on-a-chip approaches, induced-pluripotent stem cell technology, and genome editing render in vitro modeling of NDDs more robust and physiologically relevant. In this study, we review the principles underlying CNS organoid generation, their use in NDD research, and future perspectives in organoid technology. Finally, we discuss how advances in different fields could be combined to generate a multisystem organoid-on-a-chip model to investigate a specific class of NDDs, motor neuron diseases.
Introduction
Neurodegenerative diseases (NDDs) are a heterogeneous group of diseases characterized by the progressive loss of specific neuronal subsets leading to motor, sensory, and/or cognitive defects. 1 The prevalence of NDDs is increasing rapidly in our aging population, causing an increased impact on health care and society in general. Despite an urgent need for the development of new therapies, the identification of treatments for NDDs has been rather slow. 2 A main reason for this void in therapy development is that currently available animal or in vitro models often do not accurately translate human pathology. 3 This is reflected, for example, by the failure of many clinical trials for neurodegenerative and psychiatric diseases.4–6 Therefore, new models are needed to further dissect the mechanisms underlying NDDs and to identify clinically relevant therapeutic targets.
Although several two- and three-dimensional (2D, 3D) in vitro systems have become available for studying the central nervous system (CNS) in the past years,7–9 here we focus on organoid models. Despite the fact that brain organoids represent the majority of CNS organoid models, for the purpose of this review we will use the term CNS organoids except when discussing brain organoid-specific features. CNS organoids are pluripotent stem cell-derived self-organizing structures that recapitulate CNS development and allow the in vitro generation of 3D tissue that resembles the CNS. CNS organoids retain several features of the human embryonic CNS in the first and second trimester, including transcriptomic and epigenetic signatures and tissue architecture and structure.10–12
Furthermore, both whole-brain and region-specific organoids contain different CNS cell types, which emerge following a developmental timeline astonishingly similar to the in vivo process and that retain human-specific features. 13 In addition, CNS organoids can develop functional synapses, 14 show spontaneous neural activity15,16 and oscillatory events, 17 and can support interneuron migration or formation of axonal projections when fused together.18,19
Although animal models allow for monitoring complex behaviors and disease-relevant interactions between different organs, humanized models, such as CNS organoids, have the advantage of carrying the patient's genetic information and of displaying human-specific features such as the presence of outer radial glia. 20 While initially CNS organoid modeling focused on the brain or specific brain regions, recent progress has been made in the development of protocols for structures such as the spinal cord, neuromuscular junction (NMJ), and corticospinal tract (CST).21–26 Combined with the discovery of induced-pluripotent stem cells (iPSCs) 27 and the adaptation of the clustered regularly interspaced short palindromic repeats/Cas9 (CRISPR/Cas9) system for gene editing,28,29 human CNS organoids comprise an invaluable tool for modeling neural development and disease.
Brain organoids have already been used to model several NDDs and psychiatric disorders such as Alzheimer's disease (AD), Parkinson's disease (PD), Huntington's disease (HD), autism spectrum disorder, and schizophrenia.30–39 A notable exception are motor neuron diseases (MNDs) for which the application of organoids is still in its infancy. MNDs are neuromuscular diseases that affect upper motor neurons (UMNs) in the motor cortex and/or lower motor neurons (LMNs) in the brainstem and ventral horn of the spinal cord. 40
MNDs lead to degeneration of affected motor units consisting of the motor neuron (MN), NMJ, and innervated muscle fibers. Most MNDs, including progressive muscular atrophy, spinal muscular atrophy (SMA), and progressive bulbar palsy, are LMN diseases, whereas primary lateral sclerosis and pseudobulbar palsy primarily affect UMNs. Amyotrophic lateral sclerosis (ALS) is the most common MND and affects both UMNs and LMNs. In addition to motor problems, 50% of ALS patients present cognitive or behavioral symptoms, including frontotemporal dementia (FTD).40,41 Most ALS cases, and NDD cases in general, are caused by a combination of genetic, epigenetic, and other factors. Furthermore, ALS patients often differ in the age- and site-of-onset of their symptoms and in disease progression. 42 It has proven difficult to create models that adequately reflect this complexity, highlighting the need for improved patient-derived models and personalized medicine approaches. So far, this advanced modeling mostly relied on iPSC-derived 2D MN and muscle (co-) cultures43–45 and would undoubtedly benefit from the inclusion of 3D organoid models.
In this study, we review the general principles of CNS organoid generation and how these models have been used in NDD research. We address outstanding limitations and future perspectives in the field. Finally, we highlight progress made and discuss challenges for the development of advanced organoid-based in vitro systems for MNDs.
Principles of CNS organoid generation
Methods for CNS organoid generation can be classified in directed and nondirected protocols. Nondirected methods, such as the cerebral organoid protocols,20,46 solely rely on cell–cell interactions and self-organization in the absence of extrinsic patterning factors. 47 In this case, embryoid bodies are grown in serum-free medium, followed by embedment in Matrigel. These organoids contain several brain regions within the same organoid such as forebrain, midbrain, hindbrain, retina, and choroid plexus. 20 Although this approach favors the formation of neuroectoderm, it allows differentiation of cells from other germ layers. For instance, mesoderm progenitors,10,15 which can differentiate into microglia, have been reported in this model. 48 The stochastic nature of the events that lead to cerebral organoid formation results in high interbatch variability, which is a major challenge when using these protocols, especially in combination with patient-derived iPSCs, which also show high variability.49–51
The use of protocols that direct organoid differentiation through the addition of specific molecular factors aids in reducing brain organoid variability. In these protocols, patterning factors are used to either stimulate or inhibit specific morphogen pathways to mimic in vivo development and achieve regional identity. Another common feature of these protocols is the directed generation of neuroectoderm by inhibition of the bone morphogenetic protein (BMP) and transforming growth factor beta (TGF-β) pathways (known as dual SMAD inhibition 52 ). These approaches typically generate organoids with a more consistent cellular composition. 53
However, this more restricted cellular composition is also a disadvantage. Fewer cell types are present, and these organoids generally lack cells originating from other brain areas. This problem can, in part, be solved by fusing different region-specific organoids.18,19,54–56 For instance, dorsal forebrain organoids contain radial glia and glutamatergic neurons of the different cortical layers but lack interneurons that are born in the medial ganglionic eminence and that migrate into the dorsal cortex.18,55 Assembloids of dorsal and ventral brain organoids create a dorsoventral axis that enables the study of interneuron migration and integration.18,55,56 Another example is the assembly of thalamic and cortical organoids (CO) to study thalamocortical and corticothalamic projections. 19 In addition, cell types that are typically absent from brain organoids, such as microglia and endothelial vasculature, can be provided to organoids.57–62 The idea of CNS assembly using this type of approach is promising for MND modeling where multi-organ systems are needed to accurately model the disease.
Progress in the field of directed differentiation has led to generation of organoids for many different brain regions19,21,22,26,46,56,63–71 (Fig. 1). And recently, protocols to generate trunk and spinal cord organoids (SCO) became available21,22,26,72 (Fig. 1). A recent study generated organoids containing ventral, intermediate, and dorsal spinal cord domains. 26 In this study, a combination of patterning factors was used that is also applied for the differentiation of spinal MNs in 2D. However, BMP inhibitors were omitted to allow the generation of dorsal identity. This resulted in roof plate-like structures and the presence of somatosensory interneurons.

Overview of CNS regions for which organoid protocols have been developed and used in research on NDDs. Schematic of the developing brain and spinal cord indicating regions for which organoid protocols have been developed (representative references in black). Studies on the use of organoid models in NDD research are indicated in green. CNS, central nervous system; NDDs, neurodegenerative diseases.
Another remarkable advance was the generation of trunk organoids containing spinal MNs and skeletal muscle. 22 In this study, human PSCs (hPSCs) were initially differentiated into bipotent neuromesodermal progenitors, which were then grown as 3D aggregates. These aggregates recapitulated morphogenetic movements and displayed distinct neuroectodermal and mesodermal domains. After 50 days in culture, axonal tracts were present which innervated mature muscle fibers and formed functional NMJs.
More recently, Andersen et al. developed a cortico-motor unit by fusing CO, SCO, and skeletal muscle organoids. 73 These three-component assembloids contained corticofugal projections (from CO to SCO) and NMJs (spinal cord MNs innervating the muscle). Furthermore, the functional assembly of cortico-motor units was shown by muscle contraction and calcium spikes in muscle fibers after optogenetic stimulation of CO. Altogether, these models provide excellent opportunities for developing advanced 3D model systems for studying MNDs.
The variety and complexity of CNS organoid models have increased exponentially as have their use for studying disease mechanisms of neurological disorders, including NDDs.
Application of brain organoids for studying NDDs
With the development of different CNS organoid protocols, the use of organoids for dissecting the mechanisms underlying NDDs is increasing rapidly (Fig. 1). In this review, we provide a few examples of how CNS organoids have been used to study different NDDs.
AD is the most prevalent form of dementia, accompanied by cognitive decline and memory impairment. The pathophysiology of the disease is characterized by the extracellular accumulation of misfolded β-amyloid (Aβ) and tau proteins, which form amyloid plaques and neuronal tangles in cortical and subcortical regions of the brain. 74 In 2014, the first 3D AD model was generated. 75 In this study, mutant amyloid precursor protein (APP) or Presenilin 1 (PSEN1), associated with familial AD (fAD), were expressed in a neural stem cell line cultured as a 3D aggregate in Matrigel. This configuration favored the extracellular accumulation of plaques and tangles, two pathological hallmarks of AD that 2D in vitro systems and many animal models had failed to recapitulate.76,77 These findings set the stage for the further development of organoid models using iPSCs from fAD or sporadic AD (sAD) patients. Several subsequent studies described aggregation of Aβ and tau proteins in AD patient-derived iPSC-generated brain organoid cultures, demonstrating the ability of these models to recapitulate key features of AD pathology.30,78
The amount of Aβ production and its toxic species is determined by β- and γ-secretases. Therefore, modulation or inhibition of the activity of these enzymes is one of the best-studied therapeutic strategies for AD. 79 One study used CO derived from different AD patients to validate the effectiveness of several secretase inhibitors. Aβ production could be decreased when organoids were exposed to the inhibitors. 80 Recently, brain organoids generated from several fAD patient-derived iPSCs carrying a mutation in APP were used as a platform for examining the ratio of secreted Aβ peptides. This work showed how APP processing, and thus Aβ production, can vary in the presence of different APP mutations. These results highlight the importance of using patient-specific systems for disease modeling and drug screening. 81
The main genetic risk factor for AD is Apolipoprotein E (APOE), a gene involved in lipid homeostasis. 82 APOE has three alleles—ɛ2, ɛ3, and ɛ4—of which ɛ4 has been identified as the strongest genetic risk for AD. ɛ2 has a protective effect. 83 A recent study assessed the role of APOE variants in different cell types by combining CRISPR/Cas9 and brain organoid technology. 32 APOE polymorphisms were found to affect the ability of microglia to phagocyte Aβ aggregates in cerebral organoids derived from iPSC lines harboring fAD mutations. Furthermore, organoids derived from iPSCs carrying the APOE4 variant spontaneously developed features of AD pathology, including increased Aβ aggregates and tau phosphorylation.
More recently, studies have reported other AD-related pathological processes, such as mitochondrial dysfunction or tau acetylation, in brain organoid models.31,84 Interestingly, tau pathology has not only been recapitulated in brain organoids using iPSCs derived from AD patients but also from FTD patients. 85 Given that tau pathology is a common hallmark shared by different NDDs, including AD and FTD, brain organoids hold great promise as in vitro models of other NDDs.
PD is a movement disorder caused by the degeneration of dopaminergic neurons in the substantia nigra. 86 The possibility to generate midbrain organoids that contain midbrain dopaminergic neurons 67 has enabled the field to model this pathology in a more physiological setting. A few studies have so far shown that midbrain organoids derived from iPSCs carrying PD-associated Leucine-Rich Repeat Kinase-2 (LRRK2) mutations develop pathological hallmarks of PD, including increased aggregation and impaired clearance of α-synuclein, dopaminergic neuron death, and reduced neuronal complexity.33,34 Furthermore, transcriptome analysis showed that PD midbrain organoids resemble postmortem brain tissue from sporadic PD patients carrying the same mutation. 34
HD is caused by a CAG repeat expansion in the huntingtin (HTT) gene and characterized by degeneration of cortical and striatal areas. 87 A recent study generated cerebral organoids from HD patient-derived iPSCs and showed that HTT repeat expansions affect neuronal specification, maturation, and cellular organization, highlighting the neurodevelopmental component of the disease. 35 More recently, another study confirmed the effects of CAG repeat expansion in HTT on neuronal differentiation using cerebral organoids. 36
Thus far, most studies applying brain organoids in the field of NDDs have focused on AD, most likely due to its high prevalence and large societal impact. In contrast, for other NDDs, such as MNDs, organoid technology remains largely unexplored. SMA is a type of MND that affects spinal cord MNs and is caused by mutations in Survival of Motor Neuron (SMN) genes. 88 Recent work showed that SMA patient-derived iPSC-generated SCO contained a significant number of degenerating MNs demonstrating the potential of organoid cultures for studying neurodevelopmental and neurodegenerative features. 21
As illustrated by the examples above, organoid technology has great potential for increasing our understanding of the molecular and cellular basis of NDDs. However, further work is needed to generate organoid systems that can fully recapitulate neurodegeneration.
Limitations and solutions when working with CNS organoids
Despite recent advances, developing CNS organoid models that can fully recapitulate NDD pathology represents a challenge. This is particularly relevant in case of MNDs where multiple tissues are involved. Below, we highlight limitations of working with CNS organoids and discuss how current and future advances may help to generate relevant organoid model systems.
Variability and reproducibility
A limiting factor of working with iPSC-derived brain organoids is variability between organoids, both within and between individual experiments. This variability is, in part, caused by the starting material used (e.g., variability in iPSC lines and clones) but also by the currently available protocols for organoid development. In the last years, several studies have assessed organoid variability using single cell RNA sequencing. 89 Initial studies described variability within undirected brain organoids, which contain a rich cellular diversity at the expense of reproducibility.10,15 Directed protocols used to obtain brain region-specific organoids, namely forebrain and CO, showed greater organoid-to-organoid reproducibility both in terms of differentiation and cellular composition.90,91 The first protocols to generate SCO were only developed in the last 3 years.21,22,26,72 Consequently, SCO still lack the in-depth characterization that has been performed for other types of CNS organoids.
Despite being less variable compared to undirected organoids, the generation of the majority of directed organoids relies on bath application of recombinant morphogen proteins. This does not allow the generation of the tightly controlled spatiotemporal gradients that characterize in vivo morphogenesis. Furthermore, the use of recombinant proteins inevitably increases batch-to-batch variability and does not ensure stable morphogen concentrations over time, making small molecules a more reliable choice.
To date, two distinct approaches have been tested to recapitulate in vivo-like morphogenesis. One approach consisted of embedding a mass of cells expressing a certain morphogen at one pole of the organoid, creating an artificial organizing center. 92 Other studies have used microfluidics technology to create a spatiotemporal morphogen gradient93,94 or even a combination of gradients in an orthogonal manner. 95 Both approaches were successful, as the acquisition of different cell fates within the same organoid correlated with the morphogen gradient in a highly reproducible manner. The generation of in vivo-like molecular gradients during organoid morphogenesis will not only improve the reproducibility of cultures but also allow the generation of more complex and accurate CNS tissues.
Variability and low reproducibility can also be addressed using a combination of bioengineering and biomaterial approaches. While animal-derived matrices (e.g., Matrigel) are the gold standard, their composition is highly variable and poorly defined. 96 The use of chemically defined synthetic hydrogels has emerged as an alternative that supports organoid growth and allows better control of biophysical and biochemical parameters. 97 In addition, synthetic materials are also used to create biophysical constraints that allow tight control of organoid morphology.
For example, micropatterned substrates create a geometrical confinement that has been shown to induce self-organization of ectodermal cells into forebrain and spinal cord tissue in a highly reproducible manner.98,99 Other biophysics-based strategies that improve reproducibility include micropillar array chips 100 or the use of microfilaments, 101 as reviewed elsewhere. 102
For future organoid modeling of MNDs, novel patterning strategies combined with organoid fusion and the use of adequate matrices will facilitate the generation of more reproducible and complex systems.
Cellular diversity, vasculature, and connectivity
The cellular composition of CNS organoids is, especially in region-specific organoids, limited. While most of the ectodermal brain cell types are present in CNS organoids, nonectodermal cell types such as microglia, endothelial cells, or pericytes are not present in most directed CNS organoid protocols.14,103,104 The absence of nonectodermal cell types is a direct consequence of restricting cell identity to a neuroectodermal fate, a principle behind all directed CNS organoid differentiation strategies. To circumvent this limitation, several studies have successfully integrated exogenous cells into organoids.
For example, microglia were provided to brain organoids,32,59,60 making use of recently developed protocols to generate hPSC-derived microglia.57,105 This approach still requires further optimization by, for example, defining the optimal time of microglia integration and number of cells, but also by assessing, and perhaps improving, average cell survival time after integration. Given the essential role of the immune system in MND, advanced MND organoid systems would greatly benefit from the presence of microglia.
Current brain organoid models lack endothelial cells and vasculature, which impairs oxygenation and nutrient delivery to the core of the organoid. This causes metabolic stress that affects cell type specific differentiation. 106 To date, two different approaches to induce brain organoid vascularization have been described. The first approach is based on in vitro integration of vasculature and has been accomplished by coating the organoid with endothelial cells, 58 by codifferentiating endothelial cells during neuronal fate acquisition of the organoid 61 or by coculturing the initial aggregate of cells with endothelial cells 62 or mesodermal progenitors. 107
Another study mixed ETS Variant Transcription Factor 2 (ETV2)-expressing human embryonic stem cells (hESC) that differentiate into endothelial cells with hPSCs. 108 These approaches successfully integrated a vascular system into the organoids and increased the diffusion of oxygen and nutrients, but could not induce flow. The other approach introduces vasculature into brain organoids by transplantation into the mouse cortex. 109 The grafted organoids were invaded by mouse vasculature, which formed functional blood vessels. Inclusion of vasculature will not only improve the quality of the organoid systems and facilitate longer-term cultures but also allow studies on the role of the blood–brain barrier in MND, both with respect to pathology and drug delivery.
As outlined above, hypoxia limits the development and maturation of complex structures, such as a multilayered cortex and large axon bundles, in brain organoids. This limitation could, in part, be overcome by advanced culturing techniques that ensure sufficient oxygen and nutrient diffusion in long-term cultures. Organotypic cultures of brain organoids have shown to improve neuronal survival, enhance long-range and functional connectivity, 23 and to allow the development of more complex cortical structures. 110
In addition, optogenetics could be used to trigger neural activity and circuit formation. For example, iPSC-derived MNs expressing a light-sensitive channelrhodopsin were shown to trigger muscle contractions upon light stimulation in a study that developed a 3D NMJ system containing interconnected skeletal muscle and MN spheroids. 111 More recently, a study showed how novel circuits can be established in a living organism using optogenetics. 112 Therefore, we anticipate that optogenetics will be used more broadly in organoids for stimulating the development of neuronal connections, eventually enabling complex neuronal activity patterns and the formation of functional circuits.
In conclusion, reduced hypoxia and enhanced neuronal connectivity will be key in developing advanced MND organoid systems for long-term cultures that contain functional long-range corticospinal and NMJ connections.
Modeling neurodegeneration and disease progression in a neurodevelopmental system
CNS organoids present an evident limitation when used to model neurodegeneration, as they are a neurodevelopmental system that reaches a maturational plateau that corresponds to mid-gestational stages.10,113 In addition, reprogramming of adult cells into iPSCs induces cell rejuvenation and partial loss of epigenetic signatures, which can be crucial for disease modeling. 114 Given that aging is a primary risk factor for the majority of NDDs, 115 several strategies for inducing aging-related processes have been exploited, including mutating or overexpressing progeric genes116,117 and inducing reactive oxygen species and DNA damage.118,119
The artificial induction of aging by manipulation of progeric genes has proven successful, as it has enabled the emergence of disease phenotypes otherwise not present in iPSC-derived models. In addition, direct conversion strategies to bypass the iPSC stage could eventually be implemented into the CNS organoid field using an aggregate of neural stem cells directly differentiated from somatic cells as starting material.114,120 Likewise, the introduction of epigenetic modifications related to aging could be an approach to induce neurodegeneration in a more physiological manner.121,122
In addition to the induction of aging and neurodegeneration, it is crucial to develop organoid-based systems that can model disease progression. Recent advances in CNS organoid technology offer the possibility to extend the life span and maturity of these models. As the organoid increases in size, its core becomes necrotic due to a lack of oxygen and nutrients. To date, this limitation has been overcome by either generating organotypic slices or through vascularization of the organoid (as described in the previous section), both of which have been shown to enhance survival and increase neural maturation.23,123 Likewise, the use of growth factors (e.g., brain-derived neurotrophic factor [BDNF], glial cell-derived neurotrophic factor [GDNF]) promotes maturation and survival. 15 Although these improvements are not yet sufficient to increase the gestational age of CNS organoids, they extend the degree of resemblance to their in vivo counterparts. Consequently, it allows more reliable modeling of the neurodegenerative process.
The emergence and development of disease phenotypes in CNS organoids grown from patient-derived iPSC also depend on the mutation(s) they harbor and the type of NDD. For example, fAD/sAD patient iPSC-derived brain organoids require several months of culture to develop Aβ and tau pathology,30,124 whereas SMA or HD patient iPSC-derived CNS organoid models develop disease phenotypes following shorter periods of culture.21,35 To generate a model where disease progression occurs, especially when disease phenotypes appear while the organoid is still developing, introducing mutations in a time-specific manner (e.g., once the organoid has reached a maturational plateau) could be useful. 125 This would require implementation of inducible genetic systems in iPSCs and CNS organoids. 126
In summary, CNS organoids can be used to model neurodegeneration although they are inherently a neurodevelopmental system. While several strategies for inducing aging have been utilized, modeling disease progression of NDDs that are diagnosed at later stages of adulthood represents an outstanding challenge. In particular, MND modeling could be greatly benefited from organoid models where disease starts progressing in a specific tissue (e.g., spinal cord) rather than the whole system.
A multisystem organoid-on-a-chip model for MND
As discussed above, current CNS organoid models are characterized by variability and other limitations, but several solutions are available to tackle these issues in the future. Implementation of these solutions will help the design of more physiological multisystem organoid platforms for MNDs. Below we discuss how such an advanced organoid system could be assembled. The model that we propose has all the essential components of the neuromuscular system, but optimization of parts of this system, for example, further development of SCO, would already greatly progress the field of MND research.
An optimal MND organoid model would require the integration of multiple tissues, including SCO and CO, and muscle fibers. Organ-on-a-chip (OOC) technology is able to facilitate the inclusion of different tissues and modalities in one system. OOC uses microfluidics to create physiological systems, which allow the coculture of different tissues or organs in a tightly controlled environment. 127 The engineering of OOC platforms can help to design highly complex and robust organoid systems, as reviewed elsewhere,95,128,129 or to combine different organoids into a multisystem or “whole body” systems.130–133 The design of microfluidic chambers can be tailored to accommodate a variable number of chambers and inlets, as well as connections, between them providing a highly adaptable system.134–136
Such systems also allow the use of different defined hydrogels in separate chambers, which is advantageous when integrating organs with different extracellular matrix (ECM) requirements. 137 Furthermore, it is possible to add engineered features such as multiple electrode arrays (MEA), pillars, and other biosensing approaches. 130 Therefore, OOC offers a promising foundation for an organoid-based MND model. For this particular approach we envision a pumpless system, which uses gravity to generate a bidirectional flow through the use of a rocker platform (Fig. 2). The main reason for this choice is that these types of systems are easier to scale up for high-throughput purposes. 132
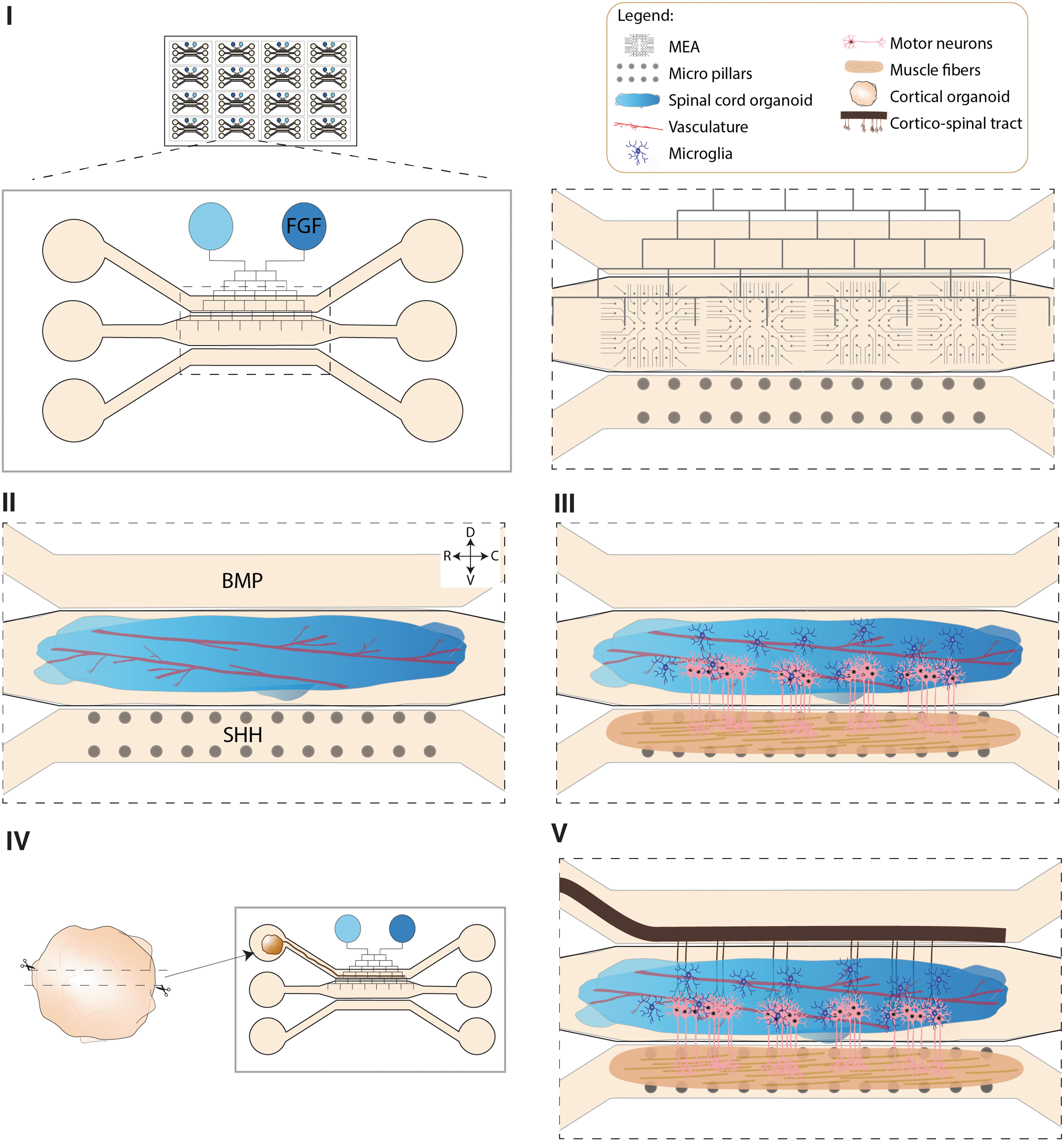
An advanced multisystem organoid-based model for MND.
The core of any advanced MND model should consist of CNS organoids (cortex/spinal cord) that consistently contain relevant MN subtypes, as well as other neuron subtypes such as interneurons. In this study, we focus on systems that have SCO at their core. To generate more representative spinal cord models, hPSCs would be seeded in an elongated chamber and subjected to an orthogonal gradient to generate caudal–rostral and dorsal–ventral axes for spinal cord patterning (Fig. 2). To induce a caudal–rostral axis, it would be necessary to add a channel network that runs on top of the main chambers connecting two inlets. 94 In this case the caudal inlet would contain fibroblast growth factors (FGF) for caudal identity,138,139 and the other would have only medium creating a regulated gradient in the central chamber with high concentration of FGF at the caudal side and no FGF on the rostral part.
For dorsoventral patterning, the side chambers would be used to deliver BMP and Sonic hedgehog (Shh) on the top and bottom chambers, respectively. Vascularization of the spinal cord tissue could be achieved by adding mesodermal progenitors or ETV2-expressing cells at the time of seeding.107,108 During the patterning period the culture would be static and it would be moved to a rocker afterward.
Once spinal cord patterning is concluded, the bottom chamber would be used to seed myogenic progenitors and generate skeletal muscle fibers 140 (Fig. 2). As specification of MN subtypes is also dependent on the types of muscle fibers these neurons innervate, 141 and given the observation that MN susceptibility is, in part, dependent on the muscle fibers MNs innervate,142,143 a mature and robust muscle model is critical for MND modeling.
The presence of a CST is a valuable addition for many MND-related research questions. To achieve this, CO differentiated separately could be sectioned and cultured in an air–liquid interface in an inlet connected to the top chamber where the axonal tract would grow and innervate the SCO (Fig. 2). Finally, microglia would be differentiated separately and integrated into the system, as previously described.32,59,60 For experimental readouts, the chip should be equipped with pillars to measure muscle contraction in the muscle chamber and MEA in the SCO and CO section inlet. Moreover, for imaging purposes the plate should be transparent.
In summary, for future MND studies we envision an organoid-on-a-chip based multisystem model consisting of spinal cord that is innervated by a CST and forms functional NMJ with muscle fibers. Inclusion of MEA and pillars is needed to assess functional readouts in muscle and neuron populations, respectively (Fig. 2).
Conclusion
In vitro modeling of NDDs greatly benefits from the development of CNS organoid technology. The development of protocols for different CNS regions and the ability to assemble them together allow the generation of complex models with reduced variability, which will further benefit their use for NDD research. Despite the limitations of current organoid models, the CNS organoid field is evolving rapidly, and more reproducible and in vivo-like systems are becoming available at a rapid pace. These advances will, in combination with progress in the areas of microfluidics, biomaterials, and tissue engineering, eventually allow the development of advanced multisystem organoid-based platforms for MNDs (Fig. 2). The organoid system we discuss includes vascularized SCO and CO and muscle tissue interconnected in a microfluidic chip that allows functional readouts and high-throughput applications in a tightly controlled environment. Implementation of this and other advanced systems will significantly further our understanding of the etiology of MNDs and will be invaluable for drug development and testing.
Footnotes
Authors' Contributions
R.V.d.S. and M.C.L. prepared a first draft of the article and prepared figures. R.J.P. provided a conceptual framework for the review, supervised R.V.d.S. and M.C.L., and edited text.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Stichting ALS Nederland (TOTALS, ALS-on-a-Chip), Stichting Parkinson Fonds, the MAXOMOD and INTEGRALS consortia under the frame of E-Rare-3, the ERANet for Research on Rare Diseases, and by the EU Joint Program Neurodegenerative Diseases (JPND; TRIAGE) (to R.J.P.).