
Abstract
Mesenchymal stem cells (MSCs) stimulate nerve and tissue regeneration and are primed for clinical translation. Application of autologous MSCs is limited by requirements for expedient harvesting procedures, proliferative expansion to increase number of cells, and reduced regenerative potential due to aging or pathological conditions. Because MSCs are immune privileged, allogeneic MSCs may serve as “off-the-shelf” cell-based reconstructive treatments to support nerve repair. Therefore, we examined the safety and immune response parameters of allogeneic MSCs seeded on NeuraGen® Nerve Guides (NNGs) in a rabbit model. NNGs with or without allogeneic rabbit MSCs were applied to rabbit sciatic nerves. Randomly assigned treatment included group I (no surgery control, n = 3) or groups II and III (sciatic nerve wrapped with unseeded or allogeneic MSC-seeded NNGs; n = 5/group). Rabbits were euthanized after 2 weeks to monitor functional recovery by histological evaluation of sciatic nerves and tibialis anterior (TA) muscle. Host reactions to allogeneic MSCs were analyzed by assessment of body and tissue weight, temperature, as well as hematological parameters, including white blood cell count (WBC), spleen histology, and CD4+ and CD8+ T lymphocytes. Histological analyses of nerves and spleen were all unremarkable, consistent with absence of overt systemic and local immune responses upon allogeneic MSC administration. No significant differences were observed in WBC or CD4+ and CD8+ T lymphocytes across unseeded and seeded treatment groups. Thus, allogenic MSCs are safe for use and may be considered in lieu of autologous MSCs in translational animal studies as the basis for future clinical nerve repair strategies.
Impact statement
Autologous mesenchymal stem cells (MSC) have been reported to enhance nerve regeneration when used in conjunction with nerve graft substitutes. However, autologous stem cell sources delay treatment and may be susceptible to age- or disease-related dysfunctions. In this study, we investigated the safety of allogeneic MSCs and the optimal number of cells for nerve conduit delivery in a rabbit model. When compared with unseeded nerve conduits, allogeneic MSC-seeded conduits did not induce a systemic or local immune response. The findings of this study will ultimately facilitate the clinical translation of a universal donor cell-based treatment option for nerve defects.
Introduction
Mesenchymal stem cell (MSC) transplantation is a promising and innovative therapeutic modality in the field of peripheral nerve regeneration. Several distinct properties of MSCs make them ideal candidates for clinical translation. 1 MSCs are multipotent stem cells capable of differentiating into various cell types, including Schwann cells, and may consequently repopulate the degenerated nerve tissue.2,3 They have also been shown to promote nerve regeneration through their trophic and immunomodulatory properties.4,5 Perhaps the most appealing characteristic for clinical use is their retention of functionality after expansion and cryopreservation. 6
Currently, autologous MSCs are often used as a biological adjunct to nerve graft substitutes, such as acellular nerve allografts and conduits, in experimental animal studies.7,8 The application of autologous MSCs is, however, limited secondary to the requisite surgical harvest of the patients' own tissue to obtain cells. A mandatory delay between harvest of tissue and application is necessary as cell expansion in tissue culture and quality control testing can take several weeks. 9 There is also evidence indicating a decline in the regenerative potential of MSCs with advanced age and in certain systemic disorders (e.g., diabetes), which would limit the use of autologous MSCs in specific patient populations.10–14
Several studies have suggested that MSCs are immune privileged or immune evasive as they express low levels of major histocompatibility complex molecules, and their paracrine cues can suppress the activation and proliferation of immune cells.9,15,16 The use of allogeneic MSCs would thus be a more appealing alternative to autologous MSCs both in large animal models and in human clinical use, as the low immunogenicity would allow for an off-the-shelf universal donor cell-based reconstructive treatment.
Preclinical and clinical studies in various disciplines, however, demonstrate conflicting results on the safety of allogeneic MSCs, with some studies demonstrating a rejection in vivo whereas others observed an immunosuppressive effect through alterations in lymphocyte reactivity.17–26 Most of these studies were either performed in in vitro models or investigated systemic administration of allogeneic MSCs, however, little is known about local transplantation in the vicinity of peripheral nerves.
It has previously been shown that MSCs can be efficiently seeded and remain viable on NeuraGen® Nerve Guides (NNGs) using the dynamic seeding protocol described by Rbia et al.27,28 NNGs are biodegradable conduits made of purified bovine type I collagen that have been approved by the Food and Drug Administration (FDA), making them suitable candidates as carriers for allogeneic MSC transplantation. The purpose of this study was, first, to determine the optimal cell seeding concentration for NNGs using allogeneic MSCs and, second, to confirm that NNGs seeded with allogeneic MSCs do not elicit an adverse immune response in vivo in a rabbit model.
Materials and Methods
All experiments were approved by the institutional animal care and use committee (IACUC A-575320). Seven-month-old outbred New Zealand white male rabbits with a mean weight of 3.45 ± 0.19 kg (SD) were used. The rabbit model was specifically chosen as they stand at the closest trunk of resemblance to human phylogeny after primates. 29 Moreover, rabbits are often used in preclinical studies evaluating the effect of various reconstructive options for large motor nerve defects. 30 Animals were housed in separate cages in an animal care facility with a 12-h light–dark cycle. They received water and food ad libitum and were allowed to move freely.
Experimental design
After isolation and characterization of allogeneic MSCs, the optimal cell density to dynamically seed MSCs onto a 10-mm NNG was determined by in vitro comparison of different cell concentrations. The cell seeding density yielding the highest seeding efficiency was subsequently used for the in vivo experiment (see optimal seeding density section). Thirteen rabbits were randomly divided into three groups. Animals in group I served as the control group (n = 3) and did not undergo surgery. In groups II and III (n = 5 per group), the left sciatic nerve was wrapped with unseeded NNGs and allogeneic MSC-seeded NNGs, respectively.
The systemic immune response to allogeneic MSCs was assessed daily by the presence of toxicity signs (tremors, convulsions, diarrhea, and lethargy) and measurement of rectal temperature and body weight. The surgical incision was inspected daily for signs of infection. Peripheral blood was collected before surgery and on postoperative days 3, 7, and 14 to perform a complete blood count and determine the lCD4+ and CD8+ T lymphocytes counts using flow cytometry analysis. All animals were euthanized 2 weeks after surgery at the peak of the acute inflammatory response.31,32 During the sacrifice, the TA muscle and organ weights were measured. The sciatic nerves and spleen were harvested and histologically evaluated by a veterinarian pathologist.
Isolation and characterization of allogeneic MSCs
As New Zealand white rabbits are maintained as an outbred colony, allogeneic MSCs were isolated from the inguinal fat pad of one donor New Zealand rabbit according to a previously described protocol. 2 Cells were expanded in advanced minimum essential medium (Thermo Fisher Scientific, Waltham, MA) supplemented with 5% (vol/vol) platelet lysate (PLTMax®; Mill Creek Life Sciences, Rochester, MN), 1% (vol/vol) penicillin/streptomycin (Penicillin-Streptomycin 10,000 U/mL; Thermo Fisher Scientific, MA), 1% (vol/vol) GlutaMAX (GlutaMAX supplement 100X; Thermo Fisher Scientific), and 0.2% (vol/vol) heparin (Heparin Sodium Injection, 1000 USP units per mL; Fresenius Kabi, Melrose Park, IL).
Medium change was conducted at 72-h intervals. Cultures were maintained at subconfluent levels in a 37°C incubator with 5% CO2 and passaged with TrypLE (Thermo Fisher Scientific/Invitrogen, Waltham, MA).
Flow cytometry was performed in passage 2 for the detection of canonical MSC surface markers CD90 (FITC mouse antihuman CD90 – 5E10; BD Biosciences, Franklin Lakes, NJ) and CD105 (APC mouse antihuman CD105 – 266; BD Biosciences) and hematopoietic stem cell markers CD34 (PE mouse antihuman CD34 – 581; BD Biosciences) and CD45 (FITC mouse antihuman CD45 – HI30; BD Biosciences). Fluorophore-conjugated isotype controls (BD Biosciences) were used as negative controls.
In brief, MSCs were washed with phosphate buffered saline (PBS) and resuspended in 2% bovine serum albumin (BSA; Sigma Aldrich, St. Louis, MO) blocking buffer for 30 min at room temperature. Cells were centrifuged, washed twice with eBioscience™ Flow Cytometry Staining (FACS) Buffer (Thermo Fisher Scientific), and incubated with the antibodies for 1 h at 4°C in the absence of light. After two more washes with FACS buffer, cells were stained with 7-aminoactinomycin D (7-AAD; Thermo Fisher Scientific) to exclude dead cells and analyzed using a BD FACSCanto™ flow cytometer (BD Biosciences).
MSCs were further characterized by their multilineage differentiation potential through adipogenic (StemPro™ adipogenesis differentiation kit; Thermo Fisher Scientific) and osteogenic differentiation (StemPro osteogenesis differentiation kit; Thermo Fisher Scientific).
Preparation of NNG
Ten-millimeter NNGs were aseptically longitudinally cut in half and reconnected with two interrupted 10-0 nylon sutures (Ethicon, Inc., Somerville, NJ) (Fig. 1A). NNGs were subsequently sterilized using γ-irradiation at a dose of 2.5 kGy. Before cell seeding, NNGs were soaked in culture medium for 2 h.

Schematic overview of dynamic cell seeding of allogeneic MSCs onto NNG and surgical procedure in a rabbit model.
Dynamic cell seeding of NNG
NNGs were dynamically seeded with various concentrations of MSCs using a modification of a previously described and validated technique. 28 In brief, 1.5 mL Eppendorf tubes were coated with pHEMA (poly 2-hydroxyethyl methacrylate; Sigma Aldrich) to prevent adhesion of MSCs to the tube. MSCs were subsequently combined with the NNG in the Eppendorf tube containing 1 mL of culture medium supplemented with 25 mM HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, Fisher Scientific) to increase the buffering capacity (Fig. 1B). The tubes were placed in a bioreactor rotator system with a fixed speed of 18 rpm in an incubator at 37°C and 5% CO2 for 12 h.
Optimal cell seeding density
To determine the optimal cell seeding density, nine NNGs were seeded with 1 × 106, 3 × 106, and 5 × 106 cells/mL (n = 3 per group). After dynamic seeding, cell counts of free-floating cells in the remaining medium were performed as an indirect measurement of the seeding efficiency. To account for anoikic cell death, seeding efficiency was also determined using a spectrophotometer (SpectraMax Plus; Molecular Devices, San Jose, CA) by subtracting the optical density (OD) of the remaining medium from the OD of the original cell concentration at an absorbance wavelength of 560 nm.
In addition, seeded NNGs were stained with Hoechst (Hoechst 33342; Thermo Fisher Scientific) and visualized with a confocal microscope (Zeiss LSM 780 confocal microscope; Zeiss, Thornwood, NY). The fluorescent intensity of the Hoechst staining was quantified using a microplate reader (Synergy H1, Biotek, Winooski, VT) at an absorbance wavelength of 361/497 nm.
The optimal cell seeding concentration was subsequently used for dynamic seeding of NNGs in experimental group III. To maintain similar conditions in group II, NNGs without MSCs were also placed in pHEMA-coated Eppendorf tubes containing 1 mL of culture medium supplemented with 25 mM HEPES in a rotator system and incubator at 37°C and 5% CO2 for 12 h.
Surgical procedure
All surgical procedures were performed under sterile conditions. Anesthesia was induced with an intramuscular injection of ketamine (35 mg/kg) and xylazine (5 mg/kg). Animals were intubated and ventilated with 1–3% isoflurane during the procedure. Body temperature was maintained at 37°C with a heating pad and intravenous lactated Ringer's was administered to maintain an adequate hydration status. In lateral position, a 4-cm incision was created in the skin of the posterolateral thigh (Fig. 1C).
The biceps femoris muscle was separated from the gluteus medius and vastus lateralis muscle to expose the underlying sciatic nerve (Fig. 1D). A 1-cm segment of the sciatic nerve was dissected and wrapped with either unseeded (group II) or allogeneic MSC-seeded (group III) NNGs. The NNGs were closed with one interrupted 10-0 nylon suture (Fig. 1E). Wounds were closed in layers using 4-0 and 5-0 absorbable sutures (Ethicon, Inc.) for fascia and skin, respectively.
Preoperatively, animals received 5 mg/kg enrofloxacin (Baytril; Bayer Animal Health, Shawnee, KS), 4 mg/kg carprofen (Rimadyl; Pfizer Animal Health, New York, NY), and 0.18 mg/kg buprenorphine (Buprinex; Reckitt Benkiser Pharmaceuticals, Inc., Parsippany, NJ) subcutaneously to prevent infection and provide analgesia. Carprofen was administered daily for 5 days postoperatively. After surgery, an Elizabethan collar (E-collar) was placed on all rabbits in groups II and III for 1 week.
General vital sign observations
Animals were observed for general behavior, food intake, signs of wound infection, and general toxicity (tremors, convulsions, diarrhea, and lethargy). Body weight and rectal temperature were monitored daily and temperatures between 38.6°C and 40.1°C were considered normal. 33
Peripheral blood collection and immune cell phenotyping
Before surgery and on postoperative days 3, 7, and 14, two whole blood samples (1.5 mL) were collected from the central auricular artery of each rabbit in EDTA tubes for white blood cell (WBC) count analysis and percentage lymphocytes and granulocytes (0.5 mL; IDEXX Laboratories, Inc., Westbrook, ME) and flow cytometric analysis of immune cell phenotypes (1 mL). Topical lidocaine 4% (Ferndale Laboratories, Inc., Ferndale, MI) was applied on the ears before blood collection as a local anesthetic and to prevent arteriospasm. The specific days were chosen to evaluate the peak acute inflammatory responses.32,34,35
For immune cell phenotyping, blood samples were combined with ammonium–chloride–potassium (ACK) lysing buffer (Thermo Fisher Scientific) and incubated for 5 min at room temperature. After centrifugation at 300 x g for 5 min, the WBCs were collected from the cell pellet and washed twice with PBS. Cells were then incubated in antibody solutions in FACS Buffer (Thermo Fisher Scientific) for 30 min at 4°C. Helper T cells (CD4+) and cytotoxic T cells (CD8+) were detected by staining with surface markers CD4 (FITC mouse antirabbit CD4 – KEN-4; Bio-Rad Antibodies, Hercules, CA) and CD8 (mouse antirabbit CD8 – 12.C7; Bio-Rad Antibodies), respectively.
In separate samples, negative IgG1 (FITC mouse IgG1 negative control; Bio-Rad Antibodies) and IgG2a (FITC mouse IgG2a negative control; Bio-Rad Antibodies) fluorescein-conjugated isotype controls were added. Stained cells were washed twice with FACS buffer and fixed with 1% paraformaldehyde.
Each sample, individual antibody fluorescence signal compensation control, unstained control, and negative isotype controls were run on a flow cytometer (BD FACSCanto, BD Biosciences). Lymphocytes were gated based on their forward scatter area (FSC-A) and side scatter area (SSC-A). Debris and duplets were subsequently excluded, and cells were gated based on their expression of CD4 or CD8 (Supplementary Fig. S1). The CD4/CD8 ratio was calculated and a ratio <1 was defined as an allograft rejection. 36
Nonsurvival procedure
After 2 weeks, animals were anesthetized and subsequently euthanized with an overdose of intravenous pentobarbital. The spleen, thymus, brain, heart, liver, and kidneys were harvested and directly weighed in grams and served as a measure for general toxicity. To evaluate motor nerve function, tibialis anterior (TA) muscles were dissected and weighted.
Histology
Ten-millimeter segments of the sciatic nerves were harvested and split into 5-mm segments for longitudinal and cross sections. Approximately, 2–3 mm of the spleen midsection was harvested. All samples were fixed in 10% formalin for 48 h at room temperature. Samples were subsequently embedded in paraffin, cut into 5-μm sections, and hematoxylin and eosin (H&E) stained. Sections were evaluated by an independent veterinarian pathologist and nerve samples were scored according to a previously described histological inflammation grading scale. 37 The pathologist was not blinded due to the importance of comparing treated groups with the controls.
Statistical analysis
Flow cytometry data were analyzed using FlowJo™ software 10.6.2 (Treestar, Ashland, OR) and cell counts for seeding efficiency were performed three times per sample. All results are expressed as mean ± standard error of the mean (SEM) with the exception of the histological score for which median and interquartile range (IQR) were used. Weight, WBC, percentage lymphocytes, and granulocytes are expressed as the mean change compared with baseline levels. Organ weights were normalized to the total body weight at sacrifice and TA muscle weight of the operated side was normalized to the value of the contralateral nonoperated side to correct for biological inter animal variation.
One- and two-way analysis of variance (ANOVA) followed by post hoc Bonferroni correction for multiple comparisons were carried out to compare groups. For histological scores, a Mann–Whitney U test was used due to the ordinal nature of the variable. Significance was set at α ≤ 0.05.
Results
Characterization of MSCs from the perivascular fraction of adipose tissue
Flow cytometry analysis demonstrated that our rabbit allogeneic MSC preparations are depleted of hematopoietic markers CD45/PTPRC (0.4%) and CD34 (0.8%) and positive for the classical MSC markers CD105/ENG (98.8%) and CD90/THY1 (92.5%). The multilineage differentiation potential of rabbit MSCs was confirmed by Alizarin Red and Oil Red staining for osteogenic and adipogenic differentiation, respectively (Supplementary Fig. S2). Hence, the rabbit MSCs used in this study represent a conventional population of MSCs.
Optimization of cell seeding density
There were no significant differences in indirect cell count seeding efficiencies between cell concentrations 1 × 106 (81.8% ± 3.6%), 3 × 106 (85.4% ± 1.5%), and 5 × 106 cells/mL (82.0% ± 3.6%) (Fig. 2A). The seeding efficiency measured by spectrophotometry showed a significantly higher percentage in 3 × 106 (68.9% ± 2.4%; p = 0.02) and 5 × 106 cells/mL (66.9% ± 1.8%; p = 0.05) than in 1 × 106 cells/mL (58.7% ± 0.4%) (Fig. 2B). Hoechst staining of seeded NNGs demonstrated cell attachment in all concentrations with visually more cell engraftment in 3 × 106 (Fig. 2D) and 5 × 106 cells/mL (Fig. 2E) than in 1 × 106 cells/mL (Fig. 2C).
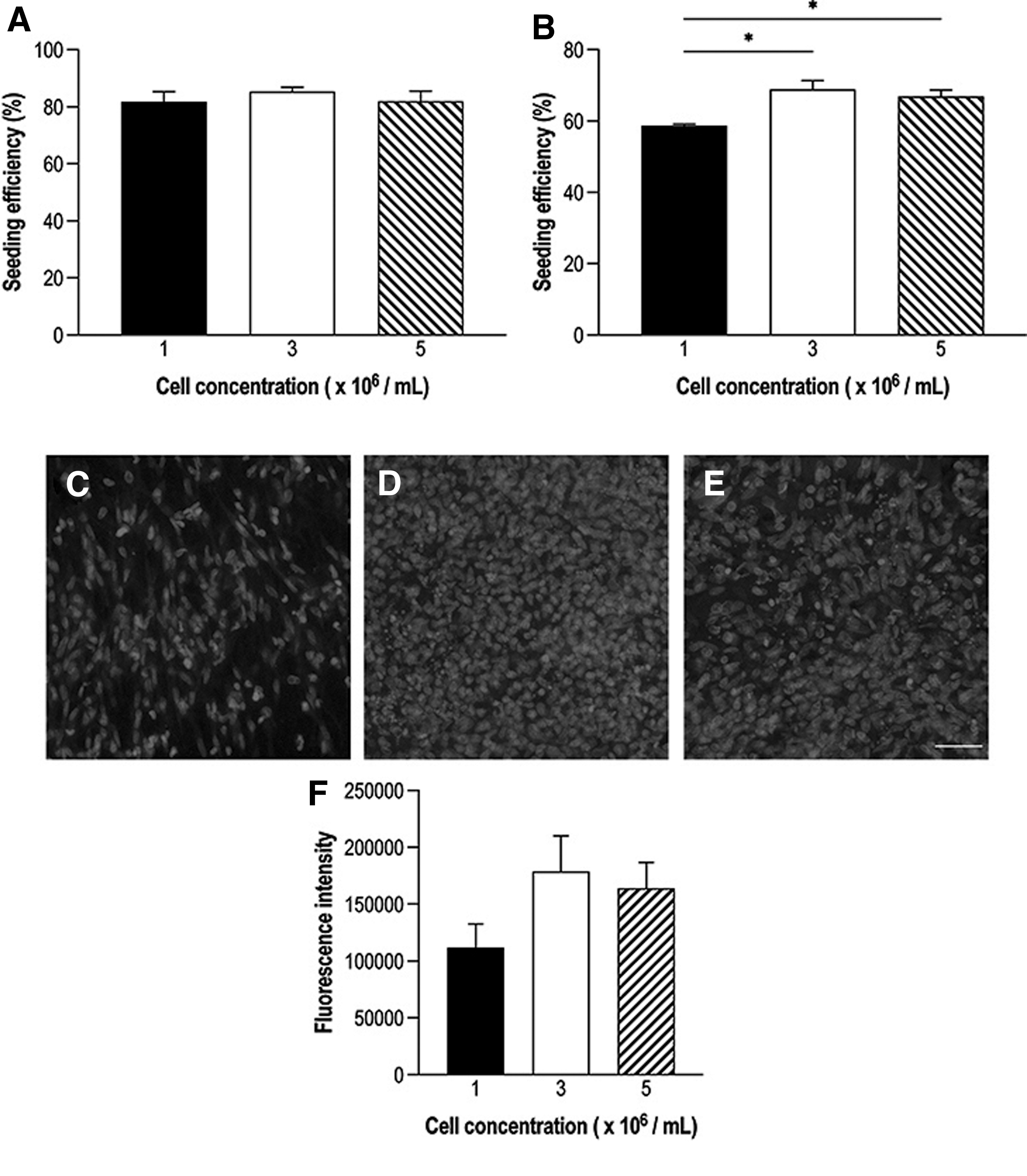
The optimal MSC seeding density of NNGs was investigated. Seeding efficiency was determined through
Quantification of Hoechst fluorescence intensity showed a trend toward higher intensities in 3 × 106 (1,78,805 ± 31,267) and 5 × 106 cells/mL (1,64,025 ± 22,771) than in 1 × 106 cells/mL (1,11,775 ± 20,928) (Fig. 2F). These differences were not statistically significant. Based on these findings, 3 × 106 cells/mL were used for dynamic seeding of allogeneic MSCs in group III.
Analysis of general vital signs
All animals survived the procedures and there were no adverse events or deaths during the survival period. Rabbits did not develop a fever and there were no signs of wound infection or toxicity. There was a decrease in weight in groups II and III compared with baseline weight between postoperative days 1 and 7 (Fig. 3). After removal of the E-collar on postoperative day 7, body weight of animals in groups II and III increased. The weight change in the treated groups was significantly different compared with that in controls from postoperative day 4 until postoperative day 14. No significant differences in weight change were observed between unseeded and allogeneic MSC-seeded NNGs.

Mean weight change over time compared with baseline weight. *p < 0.05, **p < 0.01, compared with control.
Analysis of immune responses in unseeded versus allogeneic MSC-seeded NNGs
We examined WBC counts, the percentage of lymphocytes and granulocytes, as well as the fraction of CD4+ helper T and CD8+ cytotoxic T cells to understand whether allogeneic MSCs provoke immune responses. The WBC change from baseline showed no significant differences between controls, unseeded NNGs, and allogeneic MSC-seeded NNGs over time (Fig. 4A). Unseeded NNGs demonstrated a significant decrease in percentage lymphocytes (−19.6% ± 3.5%) and an increase in percentage granulocytes (19.6% ± 3.5%) from baseline compared with controls (lymphocyte change 5.3% ± 4.9% and granulocyte change −5.3% ± 4.9%; p = 0.004) on postoperative day 3 (Fig. 4B, C).

These differences normalized on days 7 and 14. The lymphocyte and granulocyte change in allogeneic MSC-seeded NNGs was not significantly different compared with unseeded NNGs and controls at all time points. Flow cytometry analysis of levels of helper T cells (CD4+) and cytotoxic T cell (CD8+) demonstrated no significant differences between groups at all time points (Fig. 5A, B). The CD4/CD8 ratios were not inverted (<1) and not significantly different between groups over time (Fig. 5C).
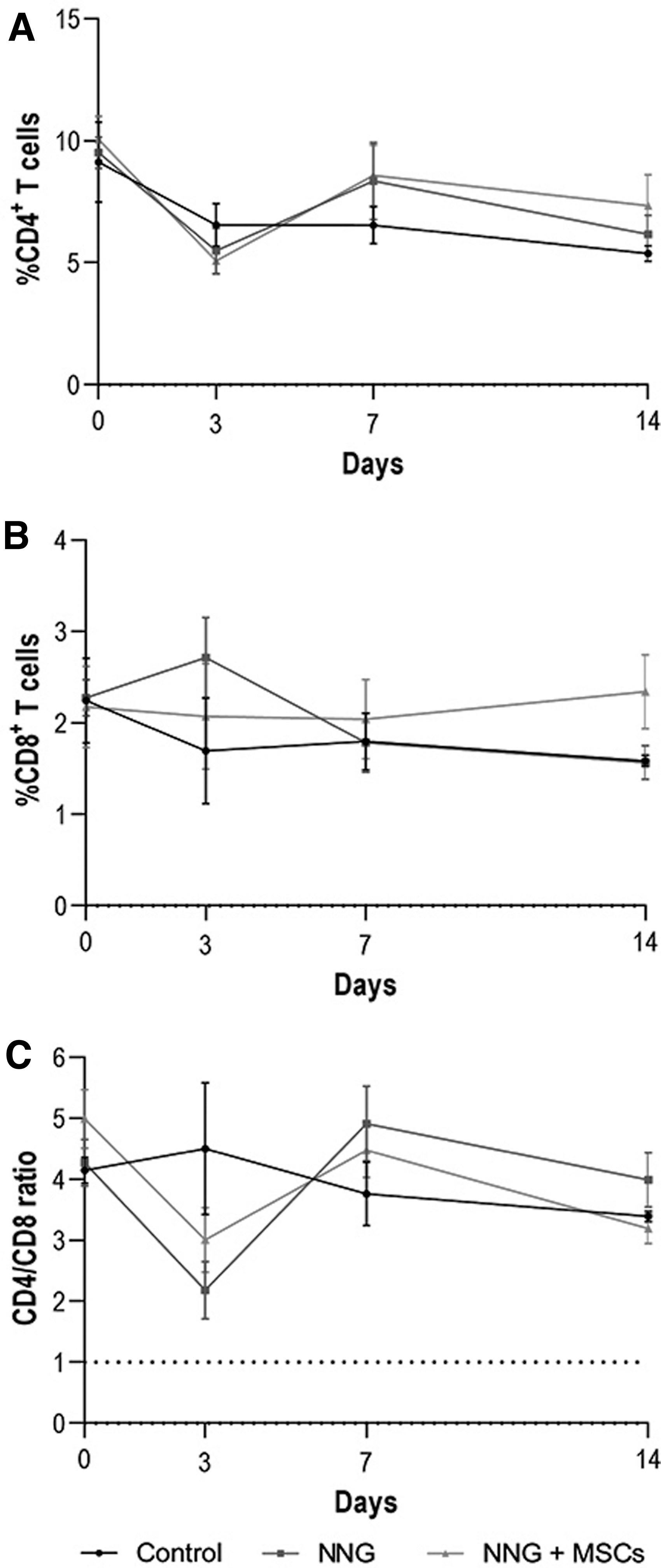
Immune cell phenotyping of
Evaluation of vital organs and TA wet muscle weight
No macroscopic pathological findings, such as hemorrhage, necrosis, tumors, or dysplasia, were observed upon visual examination of all vital organs. There were no significant differences in weights of spleen, thymus, brain, heart, liver, and kidneys as a percentage of body weight between groups (Fig. 6A).
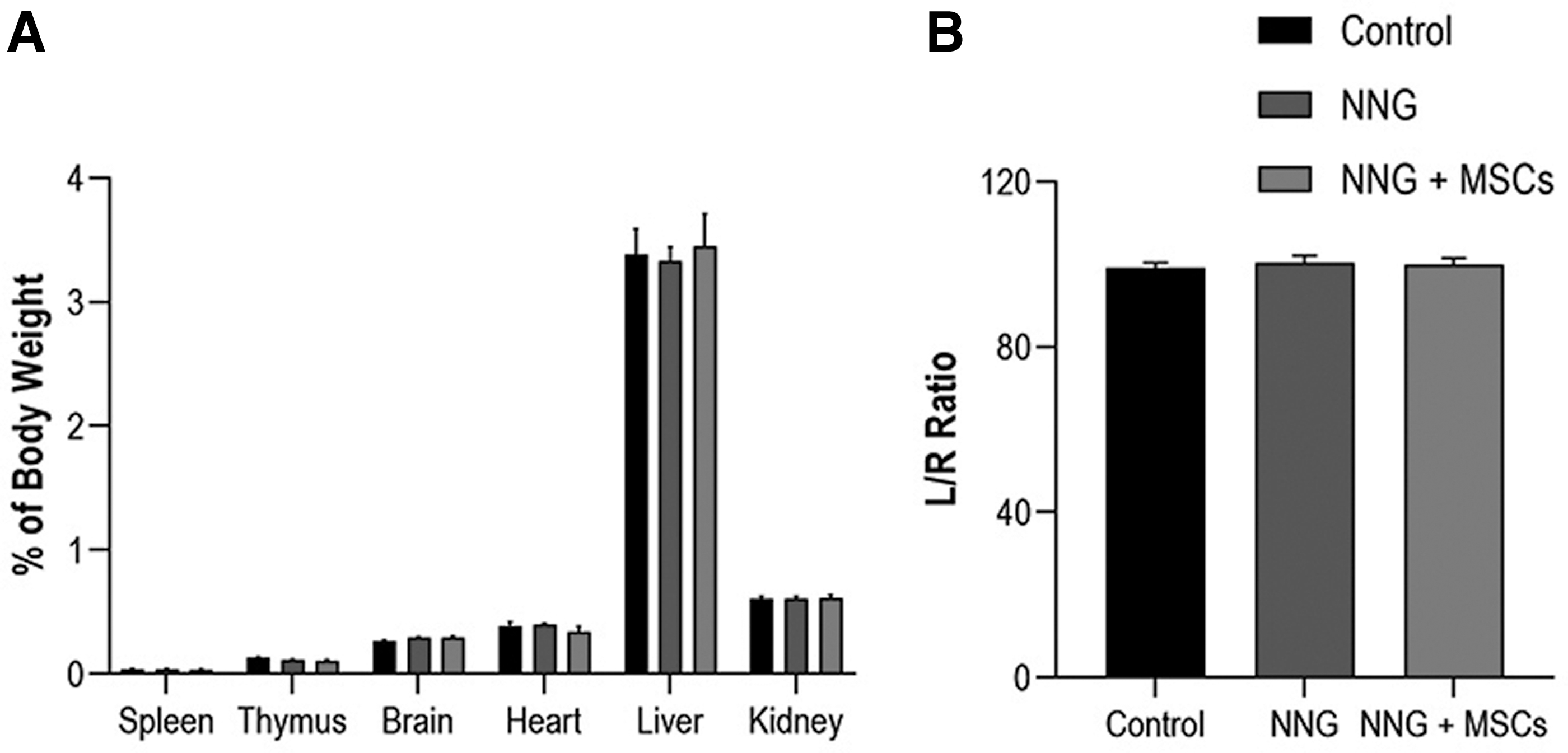
The TA muscle weight expressed as a percentage of the contralateral nonoperated side was 99.1% ± 1.2% in controls, 100.4% ± 1.8% in unseeded NNGs, and 99.9% ± 1.5% in allogeneic MSC-seeded NNGs (Fig. 6B). No significant differences between groups were found.
Analysis of spleen and nerve histology to assess systemic and local immune responses
Histological examination of H&E-stained sections of the spleen revealed a mildly increased splenic marginal zone (Fig. 7, blue dashed circles), with a mild increase in lymphocytes and macrophages, in treated rabbits compared with control rabbits. There was a mild increase in number of lymphocytes with germinal centers (Fig. 7, red circles) compared with control rabbits. The red pulp was increased in the treated rabbits, with increased number of lymphocytes and hematopoietic cells, compared with control rabbits. There were no differences in plasma cells, dendritic cells, or mast cells between the control and treated rabbits, and there was no evidence of fibrosis in all groups. No differences were seen between the unseeded and allogeneic MSC-seeded NNGs group.
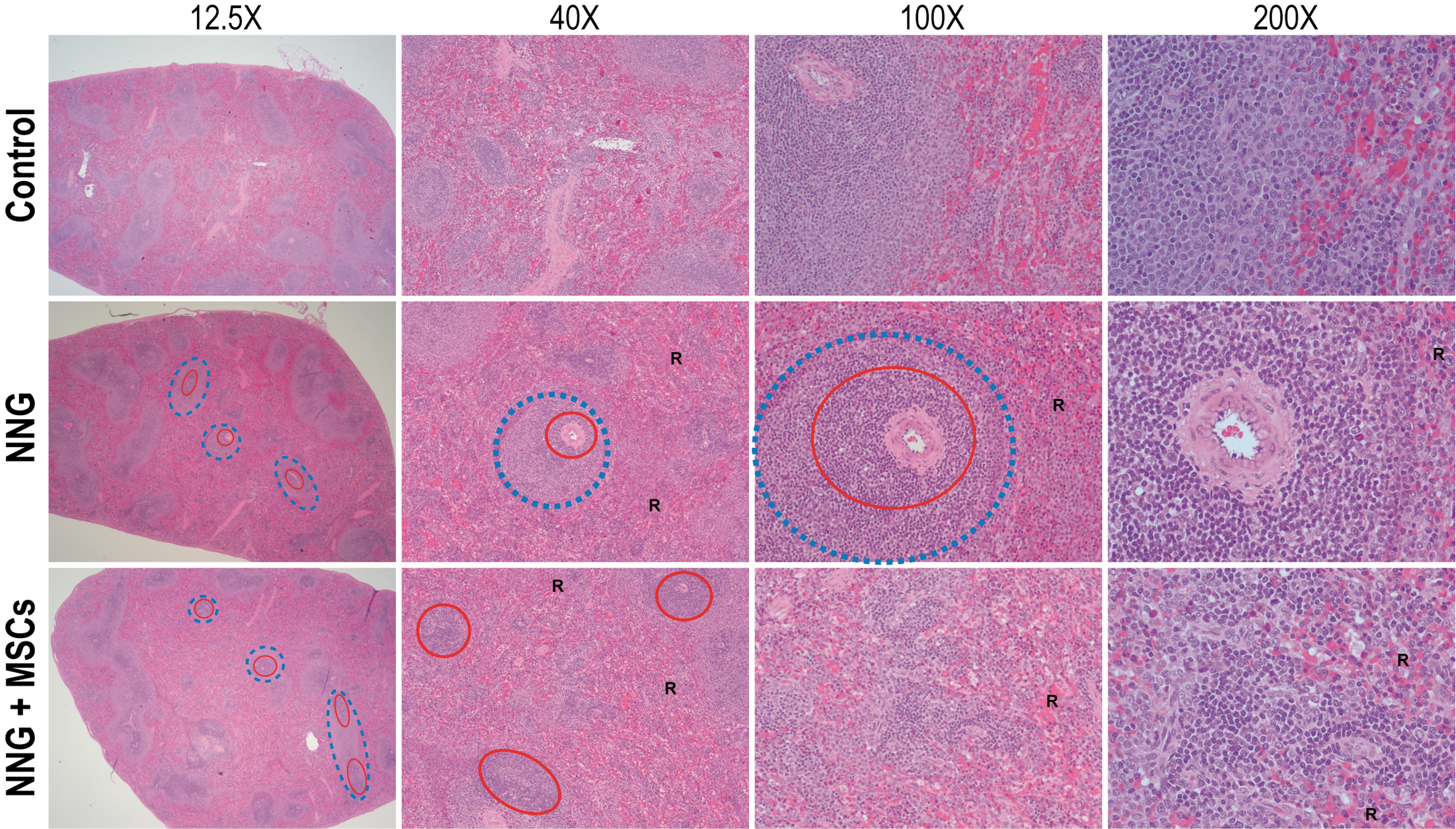
Cross sections of the spleen stained with H&E and imaged at 12.25 × , 40 × , and 100 × or 200 × . Unseeded NNGs and allogeneic MSC-seeded NNG (NNG+MSC) demonstrated a mildly increased splenic marginal zone (blue dashed circles indicate a subset of expanded marginal zones), with a mild increase in lymphocytes and macrophages, and a mild increase in number of lymphocytes with germinal centers (red circles indicate a subset of expanded follicles) compared with control rabbits. The red pulp (R) was increased in NNG and NNG+MSC compared with control rabbits. H&E, hematoxylin and eosin. Color images are available online.
Both treatment groups demonstrated infiltration of heterophils and multinucleated giant cells (Fig. 8, black circles). The nerve bundles (endoneurium) were not infiltrated, preserving the normal axonal architecture. The NNGs were evident around the treated nerves. No significant differences were found between unseeded NNGs (median[IQR], 17.0[17.0–18.0]) and allogeneic MSC-seeded NNGs (17.0[17.0–19.0]) in histological inflammation scores (p = 0.68).
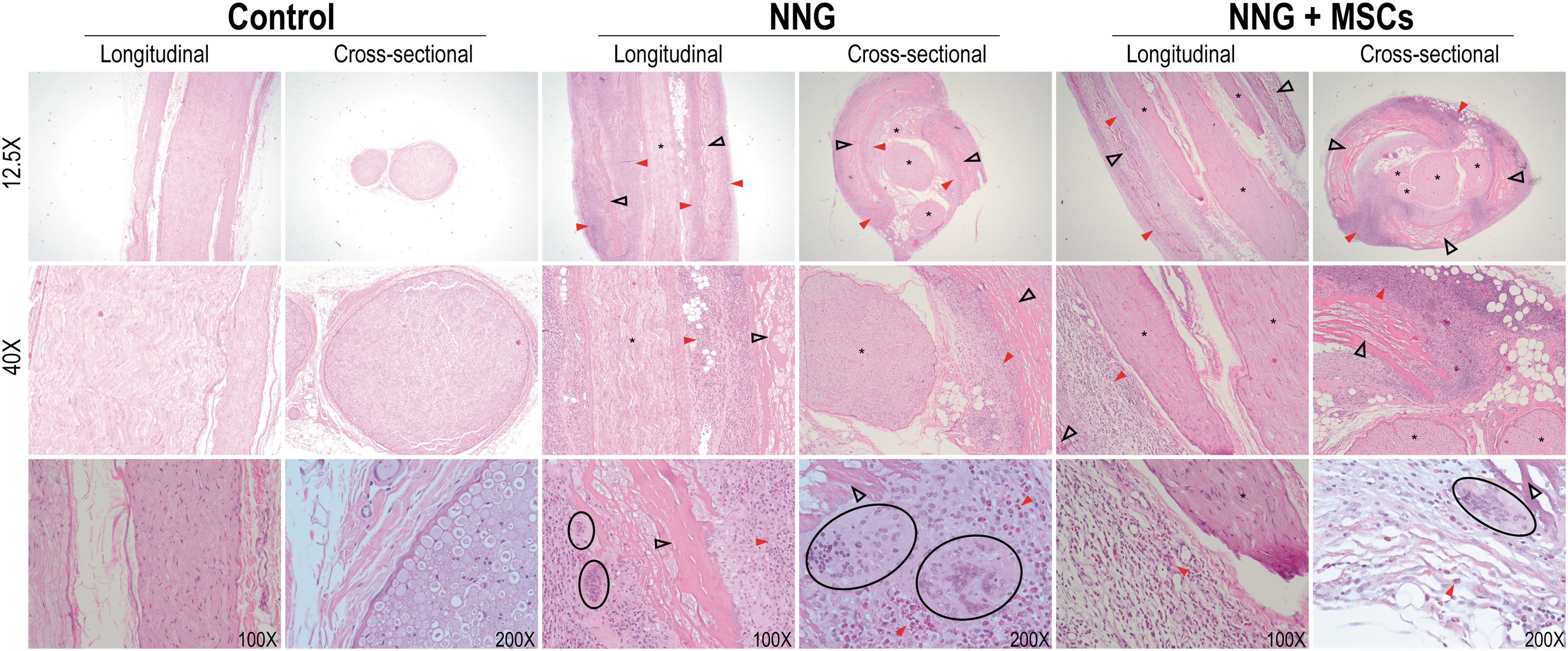
Longitudinal and cross sections of the sciatic nerve were stained with H&E and imaged at 12.25 × , 40 × , and 100 × or 200 × . In both unseeded NNGs and allogeneic MSC-seeded NNG (NNG+MSC), remnants of the conduit (black empty arrows) were visible. Both experimental groups demonstrated inflammatory infiltrate adjacent to the conduit (red arrow) composed of multinucleated giant cells (black circles). The sciatic nerve bundles (asterisks) were not infiltrated by the inflammatory infiltrate, preserving the normal axonal architecture.
Discussion
The use of MSCs is of great interest in regenerative medicine due to their multipotency, high self-renewal capacity, and low immunogenicity. 4 More notably, MSCs appear to enhance axon regeneration and myelin formation by regulating the microenvironment. 7 Optimal clinical translation of MSCs, however, necessitates tackling several issues. Existing research on MSCs in the field of nerve regeneration primarily centers around autologous resources, with a lesser focus on allogeneic MSCs. In contrast to autologous MSCs, allogeneic MSCs are readily available and are not susceptible to disease- or age-related dysfunctions. 38
Concerns regarding the in vivo immunogenicity of allogeneic MSCs in different biological contexts remain to be addressed in greater detail.17–20,39 In this study, we demonstrated that local delivery of allogeneic MSCs to peripheral nerves using NNGs did not elicit any systemic adverse effects, and was safe for use in a rabbit model.
Improving the therapeutic effect of MSC-based therapies for peripheral nerve regeneration requires optimal delivery of a sufficient number of cells to the nerve graft substitutes. Dynamic cell seeding has been described as an effective method to deliver stem cells to both nerve allografts and conduits.27,28 The advantage of the dynamic cell seeding technique is that the cells are integrated with the nerve conduit before transplantation, reducing the chance of cell death due to inability to adhere to the extracellular matrix after transplantation. Prior studies have investigated the optimal cell seeding duration, but not the optimal number of cells.27,28
In the in vitro experiments described here, the optimal cell seeding density to completely cover a 10-mm conduit was found to be 3 × 106 cells. Further increasing the seeding density to 5 × 106 appeared to reduce the number of adherent cells, as cells formed attachments to one another. Achieving the maximum number of adherent cells is crucial for optimizing the therapeutic efficacy of MSCs, as cell transplantation is inevitably associated with some degree of cell death due to a combination of cellular and host factors. 40 The use of allogeneic MSCs enables the cultivation of such large number of cells before transplantation without delaying nerve repair as would be the case with autologous MSCs.
When comparing unseeded NNGs and allogeneic MSC-seeded NNGs, we found no significant difference in systemic immune response as demonstrated by peripheral blood analyses and spleen histology. On postoperative day 3, both groups demonstrated lymphopenia and granulocytosis, with only the unseeded NNG group being significantly different from controls. This immune response is most likely attributable to the anesthesia and surgical stress, which have been shown to cause a redistribution of lymphocytes and granulocytes.41,42
The attenuated immune response observed in the allogeneic MSC-seeded NNG group may be a result of the immunosuppressive properties of MSCs. Locally, the NNG elicited an immune response consisting of multinucleated giant cells, which are frequently present in the implantation bed of resorbable biomaterials to support the degradation. 43 The role of biomaterial-induced multinucleated giant cells in the regeneration process and their interaction with MSCs are not well understood. 44
One limitation of this study is that we did not perform a direct comparison between allogeneic and autologous MSC-seeded NNGs. Based on economic (i.e., animal cost and personnel effort) and veterinary considerations (i.e., minimization of number of animals and reduction in animal suffering), we deprioritized this comparison initially. Our results established the safety of allogeneic MSCs in comparison with unseeded NNGs, thus obviating the need to examine autologous MSCs that are predicted to be safer than allogeneic MSCs in this experimental context. Other limitations include the small sample sizes, lack of in vivo cell survival measurements, and the absence of a nerve transection or defect.
MSCs have been shown to remain viable for up to 29 days in a rodent model. 45 Existing hypotheses regarding the efficacy of MSC therapy in nerve repair continue to hold that MSCs primarily serve a paracrine function and stimulate host growth factor production and tissue regeneration, which would not require permanent cell integration. 46 Our experimental cohort represents an adequate pilot project that suffices to address initial concerns with the safety of allogeneic MSCs in nerve repair. Future studies will be expanded with sufficient subjects to assess survival and potential beneficial effects of allogeneic MSC treatments and occurrence of any rare adverse events.
TA muscle weight and histological analysis of the nerve segments revealed no pathological effects of allogeneic MSCs to peripheral nerves, indicating that they are safe for further experimentation. The limitations notwithstanding, the outcomes of this study provide a basis for further research on allogeneic MSCs in peripheral nerve regeneration. Future studies should directly compare the effect of autologous and allogeneic MSCs on functional motor outcomes and gene expression profiles after reconstruction of motor nerve defects.
Conclusions
MSCs exhibit a multitude of beneficial properties, such as the secretion of trophic factors and an immune-privileged status. The use of MSCs as an adjunct to nerve graft substitutes has demonstrated promising results in peripheral nerve regeneration studies. However, autologous MSCs limit the ability to treat peripheral nerve injuries in a timely manner, which is crucial for early muscle reinnervation and subsequent motor function. In this study, we demonstrated that the local and systemic immune response elicited by allogeneic MSC-seeded NNGs did not significantly differ from unseeded NNGs, indicating that they are safe for use in future experiments aimed at improving motor nerve regeneration.
Footnotes
Acknowledgments
Authors' Contribution
M.B. contributed to conceptualization, methodology, formal analysis, investigation, data curation, funding acquisition, and writing—original draft. P.F.F. was involved in investigation. A.J.W. carried out data curation and writing—review and editing. A.Y.S. was in charge of conceptualization, methodology, funding acquisition, data curation, writing—review and editing.
Disclosure Statement
The authors have no conflict of interest to disclose. The NeuraGen® Nerve Guides used in this study were provided by Integra LifeSciences Corporation, Princeton, NJ.
Funding Information
Research reported in this publication was supported by the Mayo Clinic Orthopedic Research Review Committee (ORRC) and the National Institute of Neurological Disorders and Stroke of the National Institutes of Health under Award Number RO1 NS 102360. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.