
Abstract
Mesenchymal stromal cells (MSCs) have immense potential for use in musculoskeletal tissue regeneration; however, there is still a paucity of evidence on the effect of tenogenic MSCs (TMSCs) in tendon healing in vivo. This study aimed to determine the effects of growth differentiation factor 5 (GDF5)-induced rabbit MSCs (rbMSCs) on infraspinatus tendon healing in a New Zealand white rabbit model. In this study, bone marrow-derived rbMSCs were isolated, and 100 ng/mL GDF5 was used to induce tenogenic differentiation in rbMSC. The effects of GDF5 on rbMSC in vitro were assessed by total collagen assay, gene expression analysis, and immunofluorescence staining of tenogenic markers; native tenocytes isolated from rabbit tendon were used as a positive control. In in vivo, a window defect was created on the infraspinatus tendons bilaterally. After 3 weeks, the rabbits (n = 18) were randomly divided into six groups and repaired with various interventions: (1) surgical suture; (2) fibrin glue (FG); (3) suture and FG; (4) suture, FG, and rabbit tenocytes (rbTenocyte); (5) suture, FG, and rbMSCs, and (6) suture, FG, and TMSC. All animals were euthanized at 6 weeks postoperatively. The in vitro GDF5-induced rbMSCs (or TMSC) showed increased total collagen expression, augmented scleraxis (SCX), and type-I collagen (COL1A1) mRNA gene expression levels. Immunofluorescence showed similar expression in GDF5-induced rbMSC to that of rbTenocyte. In vivo histological analysis showed progressive tendon healing in the TMSC-treated group; cells with elongated nuclei aligned parallel to the collagen fibers, and the collagen fibers were in a more organized orientation, along with macroscopic evidence of tendon callus formation. Significant differences were observed in the cell-treated groups compared with the non-cell-treated groups. Histological scoring showed a significantly enhanced tendon healing in the TMSC- and rbMSC-treated groups compared with the rbTenocyte group. The SCX mRNA expression levels, at 6 weeks following repair, were significantly upregulated in the TMSC group. Immunofluorescence showed COL-1 bundles aligned in parallel orientation; this was further confirmed in atomic force microscopy imaging. SCX, TNC, and TNMD were detected in the TMSC group. In conclusion, GDF5 induces tenogenic differentiation in rbMSCs, and TMSC enhances tendon healing in vivo compared with conventional suture repair.
Impact Statement
Tendon tears and degeneration are debilitating clinical conditions. To date, the suture method is the only gold standard for repairing tendons. Mesenchymal stromal cells (MSCs) have been suggested for many years for their potential in tissue regeneration, especially in tendon-degenerative conditions. Growth differentiation factor 5 (GDF5) has been reported to induce human MSC into a tenogenic lineage (or TMSC), hence a potential cell source for tendon regeneration. This study reported on the potential of rabbit MSC to differentiate into TMSC via GDF5 induction and the potential of TMSC in tendon healing in a New Zealand white rabbit infraspinatus tendon model fulfilled with the 3R principle (reduce, reuse, and replace).
Introduction
Tendon is a fibrous connective tissue that contains resident cells known as tenocytes, which are responsible for tissue repair and remodeling. However, with age, the number of tenocytes in the tendon reduces dramatically. This constitutes a major contributing risk factor for tendon degeneration and poor tendon healing. 1 It has been suggested that this observation is the result of reduced extracellular matrix (ECM) production by tenocytes, which contains the proteins that provide the tendon with its strength.2,3 Tendon tears and degeneration are phenomena observed clinically, that is, in rotator cuff tears. 4 To overcome this issue, methods that involve the increase in the pool of tenocytes in tendons have been proposed, which could inevitably stimulate tendon healing (via the increase in ECM protein production) and lead to tendon regeneration, 5 as opposed to mere scar formation in normal tendon healing. This gave birth to the concept of cell-based therapies that promise superior tendon healing outcomes than those produced using conventional therapies. Several sources of cells have been considered for use in treating damaged tendons. These have included mesenchymal stromal cells (MSC or mesenchymal stem cells)6–8 and tenocytes. 9 Of these, MSCs enriched from bone marrow or fat tissue have been the preferred source due to various reasons. This includes the ease of recovery, 10 minimal donor site morbidity,11,12 and the possibility of using both autologous and allogeneic sources. 13 However, the direct application of MSCs in damaged tendons has shown to produce ectopic bone formation 14 and, in more adverse cases, tumor formation.15–17 To avoid the unintended lineage differentiation of MSCs and thus ectopic tissue formation, the use of predifferentiated MSCs before implantation has therefore been preferred in many studies.18–20
The use of biochemical, growth, and hormonal factors in MSC cultures to induce lineage differentiation is well established in many protocols.18,19 Among these, factors such as bone morphogenetic proteins (BMPs) have been used in many studies to induce MSC differentiation.18,21 One of the BMPs from the transforming growth factor beta (TGFβ) superfamily, growth differentiation factor 5 (GDF5), has been demonstrated as a pivotal factor in neo-tendon formation in in vitro experiments.22–25 Its use has also been shown to augment tendon healing when used in preclinical models.26,27 However, the tendency of GDF5 to also enhance cartilage 28 and bone 29 formation in vivo has hampered the direct use of GDF5 for tendon regeneration. Hence, indirect use of GDF5 for tendon repair, that is, the GDF5-induced MSCs, is postulated to be a better application for clinical tendon repair. To date, studies investigating the effect of GDF5 on MSC tenogenic differentiation in vitro or its subsequent effect on tendon defects in vivo appear to be limited to either immortalized cell lines, 30 rodent adipose tissue-derived MSCs, 24 or harvested non-immortalized human bone marrow-derived MSCs. 21 The effect of GDF5-induced MSCs on tendon repair in vivo has not been demonstrated. Considering that GDF5 provides positive effects on tendon repair outcomes and, for tendon neoformation (regeneration of tendons) and tenogenic differentiation ability in human MSCs, the use of GDF5-induced MSCs (tenogenic differentiation) for tendon repair is, therefore, worth exploring. This study aimed to determine the effects of GDF5-induced rabbit MSCs (rbMSCs) on infraspinatus tendon healing in a New Zealand white rabbit model.
Materials and Methods
In vitro and in vivo models
The use of in vitro and in vivo animal experimental protocols was approved by the Institutional Animal Care and Use Committee, Faculty of Medicine, University of Malaya (PM/24/06/2008/TKZ).
rbMSC isolation
Rabbit bone marrow mononuclear cells were isolated as previously described. 31 Briefly, the rabbits were euthanized by an overdose (≥50 mg per kg body weight) intravenous injection of pentobarbital sodium (Nebutal ® Sodium Solution, 50 mg/mL; Boehringer Ingelheim, Germany). Then, all the long bones were harvested in the animal operation theater and transferred to the cell culture laboratory for bone marrow procurement under sterile conditions in a biosafety cabinet. Then, the bone marrow specimens were used for mononuclear cell isolation and characterized based on the International Society for Cellular Therapy criteria as previously described. 32
Rabbit tenocyte isolation
Native rabbit tenocytes (rbTenocytes) were isolated and cultured from Achilles tendons harvested from New Zealand white rabbits.3,21,33 These cells were used for comparisons in the subsequent experiments.
GDF5-induced tenogenic differentiation in vitro in rbMSCs
The rbMSC primary cultures (at P3, n = 3) were seeded in standard 6-well culture plates at a density of 2 × 104 cells per well and cultured in an MSC culture medium with or without recombinant GDF5 supplement (100 ng/mL), similar to that in the previous study. 21 GDF5-induced rbMSCs were referred to as tenogenic MSC (TMSC). After 4 days, the cells were trypsinized and collected for total RNA extraction and real-time quantitative reverse transcription-polymerase chain reaction (qRT-PCR) analysis of candidate tenogenic marker genes, scleraxis (SCX) and type-I collagen (specifically alpha-1 collagen type-I,COL1A1). In addition, rbMSCs were also seeded on coverslips and cultured with or without GDF5 (100 ng/mL) for immunofluorescence staining analysis for candidate tenogenic marker expressions, namely SCX, TNC, TNMD, and COL-1, on day 4 postinduction with GDF5. The spent culture medium from GDF5-induced rbMSCs culture was used for the SircolTM soluble total collagen assay (Biocolor, Ireland). The primary tenocyte (rbTenocytes) cultures (n = 3) were seeded at a similar density to that of rbMSCs and used as a positive control for comparison in the total collagen assay and immunostaining analysis. The rbTenocyte cells were not supplemented with GDF5. Staining without a primary antibody was performed to serve as a negative control.
In vivo experiment
A total of 18 healthy adult New Zealand white rabbits (2.87 ± 0.53 kg and age of ∼6 months) were randomly assigned into 6 groups (n = 3 per group). The rabbits were anesthetized by an intramuscular injection of ketamine (100 mg/mL; 300 mg/kg body weight; Ilium, Australia) and Xylazil-20 (20 mg/mL; 3 mg/kg body weight; Ilium). During the induction phase of anesthesia and throughout the course of the surgical procedure, the eyes of the rabbits were protected with an ophthalmologic ointment application (Ilium Opticin, Australia) to prevent damage or dryness to the cornea. The area to perform surgery was disinfected with povidone-iodine topical solution 10% w/v (Unidon Solution, India) and locally anesthetized with 3 mL of 20 mg/mL of lignocaine hydrochloride (Ilium) via subcutaneous injection. A 3 cm incision was made on the shoulder, and the infraspinatus tendon was exposed. The infraspinatus tendons were inspected to ensure there was no preexisting lesion in the tendon or peritendinous tissues. An identical, full-thickness, window defect (about one-third of the size of the tendon; 2 mm [width] × 5 mm [length]) was dissected via a ventral longitudinal incision in the central part of each infraspinatus tendon bilaterally. The proximal and distal ends of the window defect were marked with colored (black or blue) Premilene® 3–0 nonabsorbable polypropylene monofilament sutures (B. Braun, Germany) to help identify the wound site at the time of sacrifice and dissection (Fig. 1A-1D). A silicone sheet was used to maintain the defect’s patency. After 3 weeks, the silicon sheet was removed (Fig. 1), and the defects were repaired with either suture alone (Group 1) or fibrin glue (FG) alone (Group 2), or a combination of both (Group 3). In Groups 4–6, in addition to suture and FG, defects were treated with 1–2 × 106 cells, either tenocytes (Group 4), rbMSCs (Group 5), or tenogenic differentiated rbMSCs or TMSCs (Group 6; Fig. 1E, 1F). Incision closure was completed with a simple suture using an Ethicon 4-0-coated vicryl undyed braided absorbable suture (Johnson & Johnson International, Belgium). Both tenocytes and rbMSCs used for transplantation were allogeneic cells derived from New Zealand white rabbits. Cells were mixed within the FG upon transplantation.
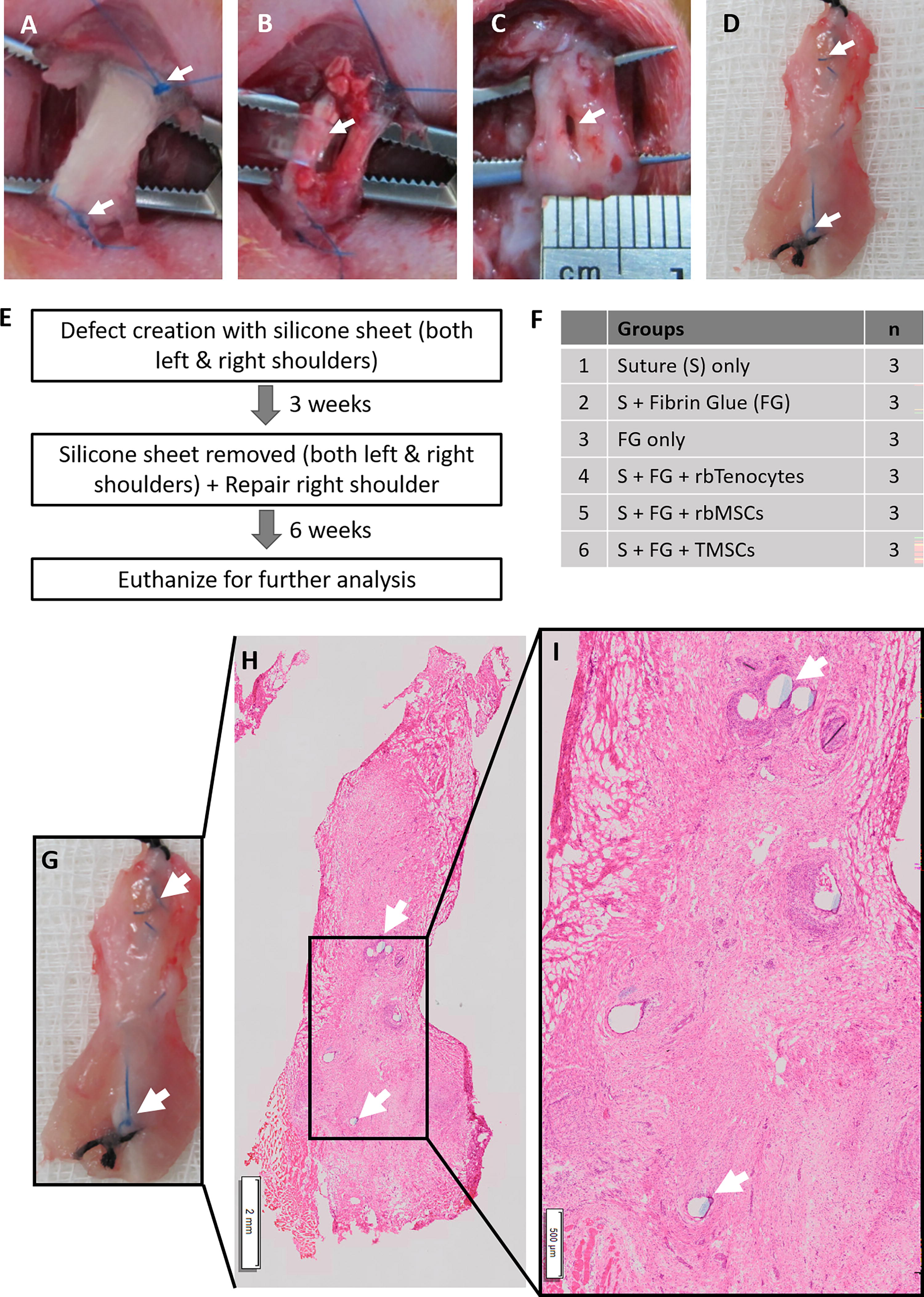
Defect creation and repair in rabbit infraspinatus tendon model.
Postoperative medication included Meloxicam injection (5 mg/mL; 0.3 mg per kg body weight; Meloxicash) and Kombitrim 240 injection (sulfamethoxazole 200 mg/mL + trimethoprim 40 mg/mL; 30 mg per kg body weight; Kela, Belgium) for 3 consecutive days at a 24-h interval to minimize discomfort during the postoperative period. The activity of the animals was not restricted. The animals were fed an ordinary diet. Mobilization and walking patterns were noted daily. After 6 weeks postoperatively, the rabbits were euthanized by an overdose intravenous injection (≥50 mg per kg body weight) of pentobarbital sodium (Nebutal Sodium Solution; 50 mg/mL; Boehringer), and the infraspinatus tendons of both shoulders were harvested for histological analysis. The specimens consisted of the entire infraspinatus tendon, beginning 5 mm proximal and ending 5 mm distal to the original defect. The defect in the infraspinatus tendon was identified by locating the marker sutures (Fig. 1D). Part of the infraspinatus muscle that is attached to the distal end of the infraspinatus tendons was collected when the tendons were harvested. The specimens were snapped frozen and stored at −80°C, before being used for cryosectioning. The tissues were sectioned in the frontal longitudinal sections (5 µm thick), including scar tissue if present. Tissue sections were stained with hematoxylin and eosin (H&E), Van Gieson (VG), and immunofluorescence (for SCX, TNC, TNMD, and COL-1) staining. The H&E staining only provides information about the pattern, shape, and structure of cells in the tendons. The VG staining allowed the observation of nuclei (blue), collagen (red), and other components (yellow; e.g., cytoplasm, muscle, fibrin, and red blood cells) at the same time. The collagen structure observed in the H&E-stained tendon tissue mounts was confirmed by the VG stain.
For gene expression analysis, 15 ten-micron-cryosections were collected in a 1.5 mL microcentrifuge tube and stored at −80°C before being further processed for total RNA extraction. Relative gene expression analyses were conducted to compare the mRNA levels of two candidate tenogenic marker genes (COL1A1 and SCX) in Groups 4–6 as well as the untreated tendon. Immunofluorescence staining of candidate tenogenic markers (COL-1, SCX, TNC, and TNMD) was also conducted.
Histology scoring
The original defect sites were localized using the marker sutures placed during surgery (Fig. 1G-1I). Stained sections were scanned and analyzed using Panoramic Viewer 1.14 (3DHistech). Histological evaluations were performed on H&E and VG-stained slides. The parameters used for histological scoring were that developed by Soslowsky and Watkins.34,35 An increase in Watkins’s scores indicates improved tendon healing, 34 whereas, for the Soslowsky’s scores, an increase in value suggests inferior healing. 35 The median values were used and compared using nonparametric tests due to the relatively small sample size used in this study. The Kruskal–Wallis test, which is equivalent to the analysis of variance, and the Mann–Whitney U test, which is equivalent to the Student’s t-test, were employed using SPSS software (SPSS). A p-value <0.05 was considered significant.
Relative gene expression analysis
Only SCX and COL1A1 gene expression levels were measured. This was due to the limited amount of total RNA that could be extracted from the minimal amount of tendon tissue (from cryosections), and that tendon also has very low cellularity compared with other tissues. The qRT-PCR analysis was conducted using a transcriptor high-fidelity cDNA synthesis kit (Roche Diagnostics GmbH, Germany) to reverse-transcribe the total RNA into cDNA and with iQ SYBR® Green Supermix (Bio-Rad Laboratories, Inc., USA) as well as the Bio-Rad CFX96™ real-time detection system (Bio-Rad Laboratories, Inc.) to perform the qRT-PCR analysis as previously described, 21 using the primers listed in Supplementary Table S1. The gene expression data were presented as normalized fold expression relative to the reference gene Glyceraldehyde 3-phosphate dehydrogenase.
Immunofluorescence staining
For fluorescence imaging, cells were seeded on coverslips or tissue sections (8 µm thick) mounted on the silanized slides (Dako, Denmark). These were then fixed with ice-cold acetone for 5 min. The specimens were rinsed twice with stain buffer (BD Pharmingen™, USA) before hybridization with primary antibodies (unconjugated mouse monoclonal or goat polyclonal antibodies; Supplementary Table S2) at 4°C overnight in a humid chamber. After overnight hybridization, the specimens were washed twice with stain buffer before proceeding to stain with fluorescence-conjugated secondary antibodies (fluorescein isothiocyanate-conjugated antimouse immunoglobulin G [IgG] and Texas red-conjugated antigoat IgG) and counterstained with nucleus stain (H33342) for 30 min at room temperature. Then, the specimens were washed twice with stain buffer before being mounted with fluoroGel mounting medium (GeneTex, Inc., USA). Fluorescence images were examined and captured either using a Nikon-Eclipse-Ti-U microscope with NIS-Element AR software (Nikon, Japan) for samples with double staining or a TCS SPII confocal laser scanning microscope (CLSM; Leica Microsystems, GmbH, Germany) with LAS AF Lite software (Leica Microsystems) for samples with triple staining. For CLSM imaging, fluorescence images were captured with sequential scanning to avoid fluorescence signal cross-talk, and line averaging (eight lines) was used to enhance the quality of the image.
Atomic force microscopic imaging
Tissue samples were processed and imaged in the air using a BioScope Catalyst™ BioAFM (Bruker Corporation, USA) in peak-force quantitative nanomechanics (QNM) imaging mode with large amplitude. All imaging was conducted in the air with a Scanasyst-Air probe (Bruker probes, nominal tip radius 2 nm, force constant 0.4 N/m). Following a 100 µm × 100 µm image captured, a rectangular region of interest with a dimension of 10 µm × 10 µm was selected along the collagen fibers, which spanned consistent topographical features (i.e., collagen fibrils).
Statistical analysis
At least three independent experiments were performed. All data were analyzed using SPSS software (version 25). The nonparametric test (Kruskal–Wallis and Mann–Whitney U tests) was used to determine significant differences between the groups, where a p value < 0.05 is considered a significant difference. The nonparametric test was used as the sample size for this study is small (n = 3). For the total collagen content analysis for in vitro experiments, the data were presented as the mean ± standard error of the mean.
For Watkins’s score and Soslowsky’s score, the data were presented as box plots with a median ± interquartile range (Supplementary Table S3).
Results
Isolation and characterization of rbMSCs in vitro
The plastic adherent enriched rbMSC was cultured and expanded in vitro (Fig. 2A). These cells were confirmed for their rbMSC characteristics by CD marker expression analysis and trilineage differentiation assay, as previously reported. 32 The GDF5-induced rbMSCs (Fig. 2B) were spindle-shaped, similar to that of the rbTenocyte culture (Fig. 2C).
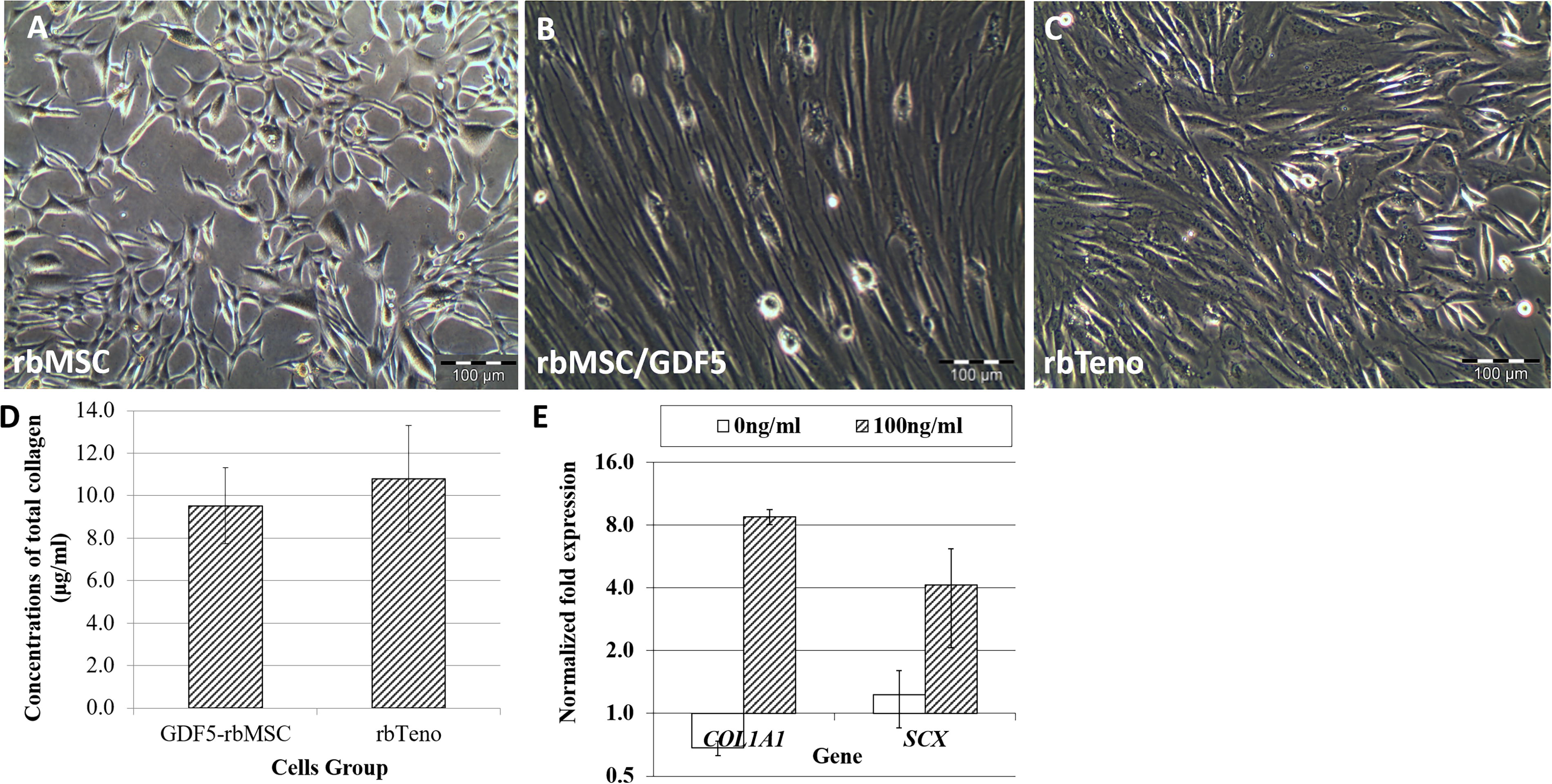
Morphology appearance of rabbit primary mesenchymal stromal cells (rbMSC) culture, growth differentiation factor 5 (GDF5)-induced rbMSCs (rbMSC/GDF5; TMSC), and rabbit tenocyte (rbTenocyte) (n = 3).
Total collagen expression levels and candidate tenogenic marker expressions in vitro
The total collagen expression levels in GDF5-induced rbMSCs were similar to that of rbTenocytes (Fig. 2D), suggesting that the GDF5-induced rbMSCs underwent tenogenic differentiation with a tenocyte phenotype. In gene expression analysis, the GDF5-induced rbMSCs also showed a significant (p < 0.05) upregulation in COL1A1 and SCX mRNA expression levels (8.76 ± 0.71 folds and 4.12 ± 2.05 folds, respectively; Fig. 2E). The results of immunofluorescence staining revealed that COL-1, TNMD, TNC, and SCX were expressed in the GDF5-induced rbMSCs (Fig. 3). These observations suggest that GDF5 could induce tenogenic phenotypic expression in rbMSCs.

Immunofluorescence staining of (
Tendon defects transplanted with TMSCs showed enhanced tendon healing in vivo
The animals showed restricted movement up to 3 days after the defects were created. The peritendinous fibrous reaction was noticeable during the second surgery at 3 weeks after the initial defect creation. At the time of harvest at 6 weeks postsurgical repair, animals showed an equal range of motion of the shoulders bilaterally. There was no detectable difference in the activity level of the animals in each group. Macroscopically, there was no gross evidence of infection at the surgical sites (Fig. 4A). In the non cell-based-treated groups (suture only, FG only, and combination of suture and FG), the tendon defect area was filled with pale fibrous scar tissue similar to that of the contralateral untreated infraspinatus tendons (Fig. 4A). Progressive tendon healing was prominent in the TMSC (or GDF5-induced rbMSC) group, where noticeable tendon callus (neo-tendon) formation was observed (Fig. 4B). Early tendon callus formation suggests that the use of TMSCs stimulates neo-tendon formation. We observed no evidence of cartilage formation or ectopic bone formation in any of the specimens. In addition, the histological analysis (H&E and VG) was carefully evaluated for the presence of cartilage or ectopic bone formation. We found no cartilage formation or ectopic bone formation at 6 weeks after tendon repair.

The histological appearance at 6 weeks postsurgical repair for infraspinatus tendon defects treated with suture alone (S), fibrin glue alone (FG), and a combination of suture and fibrin glue (S+FG), compared with that treated with suture and fibrin glue in addition of cells (S + F+ cells), rbTenocytes, rbMSCs, or TMSC, and untreated (control without repair) (n = 3).
The microscopic appearance of a normal tendon is an organized matrix of collagen fibers bundled parallel to each other and follows an arrangement parallel to the tendon’s longitudinal axis (Fig. 4C). The tendon cells that reside in the normal tendons are usually spindle-shaped, with an elongated nucleus aligned parallel to the bundles of collagen fibers. In this study, untreated (Control-no replair) tendons showed disorganized collagen fibers and increased cellularity. The cells appeared randomly aligned with a plump or rounded morphology (Fig. 4C). Histological analysis revealed that tendon defects treated with different methods showed different stages of healing, that is, early, moderate, or mature, at 6 weeks postsurgery. In non cell-based-treated groups, collagen fibers from repaired sites appeared highly disorganized, having cells with plump morphologies and rounded nuclei, as well as evidence of blood vessels. This indicated that these tendons were at the moderate healing stage as compared with the early healing stage in the untreated tendons (Fig. 4C). In cell-based-treated groups, the fibers appeared more organized and were arranged parallel to the long axis of the tendons. The TMSC group demonstrated mature tendon healing, as evidenced by the presence of cells that have elongated nuclei aligned parallel to the collagen fibers. In addition, the collagen fibers in the TMSC group were more organized (i.e., collagen fibers aligned into bundles) than in the other groups (Fig. 4C). In short, the healed tendon in the TMSC group had a close resemblance to that of normal tendon tissue (Fig. 4C) at the endpoint (week 6).
In normal tendons, VG-stained sections show mature collagen fibers, as evidenced by the deep red color. In the present study, there appears to be reduced intensity in all samples tested, demonstrating the presence of immature collagen fibers at tendon repair sites (Fig. 4D). This observation was not unexpected due to the new collagen fiber formation at the tendon repair site. In addition, VG staining also revealed an abundance of blood vessels in the non cell-based-treated groups as well as the untreated tendons.
Both Watkins’s (p = 0.008) and Soslowsky’s (p = 0.008) scores (Fig. 4E and 4F; Supplementary Table S3) showed significant differences between the treatment groups. Groups treated with cells were significantly superior to groups not treated with cells in both scoring systems. Among the cell-based-treated groups, significant differences were observed in the rbTenocyte-treated group compared with either the rbMSC- or TMSC-treated groups in the Watkins’s scores.
To further compare the cell-based-treated groups, gene expression analyses, immunofluorescence staining, and atomic force microscopy (AFM) imaging were conducted in the cell-based-treated groups, compared with that of a normal tendon (without defect) and the untreated (tendon defect without repair or no repair). This is to show the improvement in tendon healing observed in the cell-based-treated groups compared with that of a tendon in the normal condition and with a defect without treatment (no repair group). The qRT-PCR analysis showed a significant (p < 0.05) upregulation in the relative gene expression levels in the TMSC group for COL1A1 (a 0.45 ± 0.02-fold) and SCX (a 3.20 ± 0.28-fold) compared with the rbTenocyte group (a 0.11 ± 0.01-fold and a 0.38 ± 0.08-fold for COL1A1 and SCX, respectively) and compared with the rbMSC group (a 0.02 ± 0.01-fold and a 0.30 ± 0.06-fold for COL1A1 and SCX, respectively).
Immunofluorescence staining for candidate tenogenic markers (COL-1, SCX, TNC, and TNMD) revealed the tissue distribution of each protein on normal and repaired tendons (Fig. 5). The stained nucleus demonstrated similar morphologies between the TMSC group and that of normal tendons. These nuclei were observed mainly between the collagen bundles, which were stained intensely with COL-1. The COL-1 bundles were aligned parallel to each other in both TMSC and normal tendons. SCX, a transcription factor used as a candidate tenogenic marker, 36 was detected more intensely in the intracellular region of the tenocyte-like cells in both the normal and TMSC groups, and dimly expressed in the tenocyte and rbMSC groups (Fig. 5). The TNC expression was detected throughout the tendon specimens in normal tendons as well as the TMSC group; however, localization patterns were found in the untreated, tenocyte, and rbMSC groups. The TNMD was only observed more intensely in the intracellular region of the tenocyte-like cells in the normal tendon and TMSC groups. The expression pattern of TNMD in TMSC was comparable to that of normal tendons. To further evidence the improvement in collagen fibril orientation in the cell-based-treated groups, AFM imaging was conducted. The AFM images showed an improved collagen fibril orientation in the rbMSC and TMSC groups compared with that of the control group (Fig. 6). Although both rbMSC and TMSC groups showed collagen fibrils in an organized parallel direction, the collagen fibrils in the rbMSC group were smaller in diameter compared with that of the TMSC and normal tendon. It is worth noting that no mineralized fibril bundle was observed in the tendon repair site (in all groups), indicating that there was no ectopic bone formation.

Confocal images of COL-1, SCX, TNC, and TNMD on normal, untreated, and cell-based-treated (G4, G5, and G6) groups (n = 3), at 6 weeks postsurgical repair (field of view within the repaired site). Immunofluorescence staining was performed among the cell-based-treated group to see the improvement in tendon healing compared with that of a normal tendon and a tendon defect left without treatments. For immunofluorescence staining, the infraspinatus tendons were collected for cryosectioning. The tendon tissue sections (8 µm thick) mounted on the silanized slides were used for immunofluorescence staining. The COL-1 and SCX were more intensely observed in normal and G6 (A). The TNMD showed a wavy pattern on both normal and G6 (B) (left to right panels: H33342, COL-1 [or TNC], SCX [or TNMD], and merged images. Sb: 50 µm).

Tapping mode atomic force microscopy (AFM) topography (height) images of tendon collagen fibrils, obtained from normal, untreated, and cell-based-treated (G4, G5, and G6) groups (n = 3), at 6 weeks postsurgical repair. AFM imaging was performed among the cell-based-treated group to see the improvement in tendon healing, in terms of the tendon collagen fibers orientation, compared with that of a normal tendon and a tendon defect left without treatments. For AFM imaging, the tendon tissue sections (8 µm thick) mounted on the glass slides were fixed with 4% glutaraldehyde and processed with the hexamethyldisilazane method to dry the samples. The images showed the collagen fibril organization in the tendon defect area. The normal tendon appeared to have collagen fibrils organized in parallel orientation to the tendon’s long axis. The collagen fibrils in the untreated and G4 were in random orientation. Both G5 and G6 showed tendon fibrils arranged in a parallel orientation similar to that of the normal tendon.
Discussion
The present study demonstrated that GDF5 induces rbMSCs to differentiate into the tenogenic lineage. This deduction was based on the following findings: (1) total collagen expression levels in GDF5-induced rbMSC were similar to that of rbTenocytes; (2) mRNA expression levels of COL1A1 and Scx were significantly upregulated in rbMSCs supplemented with GDF5; and (2) GDF5-induced rbMSCs expressed tenogenic phenotypic markers, that is, COL-1, SCX, TNC, and TNMD. When transplanted in vivo, the tendons repaired with TMSCs showed better tendon healing as compared with rbTenocytes and rbMSCs. This deduction was based on the following findings: (1) early tendon callus formation in TMSC group when observed macroscopically; (2) tendon healing in TMSC group showed that cells resembled tenocyte-like phenotype, that is, cells with spindle-shaped and elongated nucleus, which were aligned parallel to tendon’s longitudinal axis; (3) collagen fibers in TMSC group were in organized patterns as compared with the randomly aligned collagen fibers in non cell-based-treated groups as well as untreated tendons; (4) both Watkins’s and Soslowsky’s scores demonstrated tendons from the TMSC group showed better repair quality as compared with the non cell-based-treated groups (Supplementary Fig. S1 and Supplementary Table S3); (5) gene expression analysis showed that the tendon of the TMSC group expressed significantly higher COL1A1 and SCX compared with the tenocyte and rbMSC groups; and (6) candidate tenogenic marker was expressed in the TMSC group with the tissue distribution comparable to that observed in the normal tendon; and (7) improved collagen fibril orientation in the rbMSC and TMSC groups compared with that of the control group, from the AFM topography imaging.
In monolayer culture, our previous study demonstrated that 100 ng/mL of GDF5 produced an upregulation of tenogenic marker genes with a downregulation of nontenogenic, that is, osteogenic and chondrogenic gene markers, in human MSCs. 21 To determine the effects of GDF5-induced rbMSC or TMSC in tendon healing, an evaluation of the tenogenic differentiation effect of GDF5 on rbMSCs must be conducted before the in vivo study commences.
In this study, the tendon–tendon repair model was used, whereas previous studies utilized the tendon–bone junction repair models. Similar tendon callus formation that was observed in the TMSC group had been reported in rat tendon–bone junction repaired with the suture at 6 weeks postrepair 37 and in tendon–tendon healing in rat Achilles tendon after 1 week of platelet concentrate injection. 38 Factors that can contribute to tendon callus formation at the repair site are the presence of paratenon 39 and intact periosteum. 37 In this study, since there was no paratenon or periosteum involved, the paracrine effect from the transplanted TMSC could be the factor that contributes to tendon callus formation. A recent study also reports the important role of bursae in tendon regeneration. 40
Transplanted TMSC can participate in tendon repair in two possible manners: (1) by proliferation to the appropriate number of cells and subsequent differentiation into mature tenocytes for tendon healing and (2) by mediating the behavior of cells involved in the repair process through paracrine signaling.41,42 The transplanted TMSC may secrete trophic factors that are highly stimulatory to tendon ECM production, that is, TGFβ 43 and bone morphogenetic protein 2 (BMP2), 44 which can play a role in regulating differentiation and healing kinetics. 45 However, the interaction between TMSC paracrine signaling and ECM cues and how they affect progenitor cell differentiation at the healing tendon remains to be elucidated.
In line with the 3R (replace, reuse, and reduce) principles for animal in vivo studies, this study maximized the outcome measures that we may have from the total of three animals in each group. Nevertheless, there are several limitations identified within the present study that future studies should avoid. First, tissue recovery was only performed at a single time point, that is, 6 weeks. Previous studies involving tendon repair have reported a longer recovery time of 8–12 weeks, 35 whereas others conducted their study with more than one recovery time point. 46 Despite not having either, the present study was able to observe distinct apparent changes in tendons treated with different methods, which indicates that the time points used in this study were sufficient to observe the effects of TMSC in tendon healing.
Second, like all other previous studies, the question as to whether cells transplanted at tendon defect sites are directly responsible for the repair or if it was because of the factors released or other possibilities could not be answered in the present study. The use of fluorescence and bioluminescence imaging, that is, green fluorescence protein (GFP)-transfected MSCs for in vivo cell tracking or the use of a reporter gene system consisting of enhanced GFP for molecular imaging, would be of advantage. The observed tendon repair in the TMSC group (i.e., tendon callus formation, collagen fibers in oriented organization, and candidate tenogenic marker expression), which was not detected in all the other groups (treated and untreated control), suggested that these observations were due to the implantation of TMSC at the tendon defect area. Nonetheless, the GFP-labeled MSCs, especially those with lentiviral-based transduction (for producing a comparatively stable and longer-term GFP expression), have their limitations; especially, those using HIV-based vectors 47 may induce an unwanted immune response in vivo 48 as well as insertion mutagenesis in the animal. 49 Lastly, the use of other functional assessments and quantitative outcome measures such as biomechanical testing would allow objective measurements for regaining function postrepair, which is not possible via histology or biochemistry analysis. The data presented in this study shall be interpreted with caution without the biomechanical testing data. Future studies shall include measures that would overcome these limitations, with a larger sample size for better power of the study.
Conclusions
The present study demonstrates that GDF5-induced rbMSCs show tenogenic differentiation potential. When applied to the damaged tendon, TMSCs (GDF5-induced rbMSC) produced enhanced tendon healing compared with conventional suture repair.
Footnotes
Acknowledgments
The authors thank the University of Malaya for a PhD thesis scholarship for the first author; Dr. Haryanti Azura Bt Mohammad Wali, as a veterinarian, from the Animal Experimental Unit, Faculty of Medicine, University Malaya, for her help in postoperative care and pain management for the animals; Ms. Hak Savatey and Ms. Sahrinanah Mappiare, Medical Laboratory Technologist, for their assistance in animal handling; and The Advanced Imaging Laboratory, Faculty of Medicine, University Malaya, for the AFM imaging services provided.
Authors’ Contributions
S.-L.T.: Formal analysis (lead), methodology (lead), conceptualization (lead), writing—original draft (lead), formal analysis (lead), and writing—reviewing and editing (lead); C.-K.C.: Formal analysis (equal), original draft (supporting), and writing—reviewing and editing (equal); TSA.: conceptualization (supporting), writing—original draft (supporting), and writing—review and editing (equal); S.-H.T.: Formal analysis (equal), original draft (supporting), and writing—reviewing and editing (equal); W.-M.N.: Formal analysis (equal), original draft (supporting), and writing—reviewing and editing (equal); L.S.: Conceptualization (supporting), writing—original draft (supporting), and writing—reviewing and editing (equal); T.K.: Conceptualization (lead), writing—original draft (supporting), and writing—reviewing and editing (equal).
Institutional Review Board Approval
The use of in vitro and in vivo animal experimental protocols was approved by the Institutional Animal Care and Use Committee, Faculty of Medicine, University Malaya (PM/24/06/2008/TKZ).
Disclosure Statement
No competing financial interests exist. All authors declared that they and their immediate families and any research foundation with which they are affiliated did not receive any financial payments or other benefits from any commercial entity related to the subject of this article.
Funding Information
This work was supported by the High Impact Research-Ministry of Higher Education (HIR-MOHE) research grant initiative (UM.C/625/1/HIR/MOHE/MED/04, E000003-20001), eScience Grant (12-02-03-2017), and University of Malaya postgraduate student grant (PS167/2008C, PS360/2009B, and PS219/2010A).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.