
Abstract
Decellularization does not completely remove the matrix-bound α-Gal epitopes in porcine acellular dermal matrix (pADM), and the presence of residual α-Gal epitopes could elicit adverse immunological reactions and cause potential early failure of xenografts. The present study had evaluated the effectiveness of decellularization and α-galactosidase treatment to eliminate the matrix-bound α-Gal epitopes in pADM, as well as the effect of tissue form (intact pADM vs. microparticle). Decellularization eliminated ∼80% of α-Gal epitopes in porcine dermis, and pADM retained ∼20% of the matrix-bound α-Gal epitopes. While Aspergillus α-galactosidase and Coffea α-galactosidase both hydrolyzed the terminal alpha-galactosyl moiety from oligosaccharides, only Coffea α-galactosidase was effective in eliminating the matrix-bound α-Gal epitopes in intact pADM. Aspergillus α-galactosidase did not work for intact pADM, even at an enzyme activity more than an order of magnitude higher than that of Coffea α-galactosidase used. The different efficacy between Aspergillus α-galactosidase and Coffea α-galactosidase was associated to the accessibility to the matrix-bound α-Gal epitopes in intact pADM. When intact pADM was micronized into fine microparticles, Aspergillus α-galactosidase and Coffea α-galactosidase eliminated the matrix-bound α-Gal epitopes equally well. Thus, the tissue form had significant influence on the efficacy of enzymic cleavage. The findings of the study offer valuable insight for enzyme selection and process development for efficient α-Gal antigen reduction in xenogeneic grafts or tissue scaffolds.
Impact Statement
Decellularization does not eliminate the matrix-bounded xeno-reactive α-Gal epitopes, and immunological responses are potential causes for early failure of clinically implanted xenografts. The present study investigated the effectiveness of different α-galactosidases and the effect of tissue forms in eliminating the α-Gal epitopes in porcine acellular dermal matrix. The finding of the study offers valuable insight for the xenogeneic antigen reduction in processing xenogeneic tissue grafts or scaffolds.
Introduction
The α-Gal (Gal α1,3-Galβ1-4GlcNAc-R) epitope, a carbohydrate antigen that appeared early in mammalian evolution, represents a major barrier in the animal-to-man organ and tissue xenotransplantation. Nonprimate mammals such as pigs synthesize the α-Gal epitope in large quantity via the glycosylation enzyme α−1,3-galactosyltransferase.1,2 Because humans lack α-Gal epitopes and produce the natural anti α-Gal antibody which bind specifically to α-Gal epitopes,3,4 the transplantation of porcine tissues and organs into humans could result in immunological reactions due to the anti-α-Gal binding to xenograft’s α-Gal epitopes. For implanted bioprosthetic heart valves and grafted xeno-skin, the early failure and/or degeneration of xenografts is commonly attributed to the anti α-Gal response in human patients.5–9 The studies on the sera of patients who underwent pulmonary and/or heart valve replacement with porcine bioprostheses have clearly revealed that the implantation of animal-based bioprosthesis elicited the increased formation of anti-α-Gal antibodies in patients, with IgM in a more rapid transient response and IgG in a delayed longer change.10,11 Similar anti-α-Gal specific IgG immune responses were also reported by other workers after implantation of valve bioprostheses.12,13
The elimination of the α-Gal epitopes in xenografts is of great importance for a successful xenotransplantation, which could overcome the severe shortage of allogenic tissues available for human implantation. One of the strategies is decellularization and enzymic cleavage using α-galactosidase to eliminate the α-Gal epitopes.8,13–16 Decellularization removes immunogenic cell materials, and enzymatic cleavage can further eliminate the extracellular matrix-bound antigens, which improves the biocompatibility of derived xenografts. The approach has been applied to cartilage, 17 dermis,14,18 heart valve,13,19,20 ligament, 16 pericardium,8,15,19–21 tendon, 22 and vein. 23 The α-galactosidase treatment was found to effectively remove the α-Gal epitopes from these animal tissues. In vivo implantation of the xenografts treated with the α-galactosidase (i.e., ligament, pericardium, skin, and tendon) elicited no or minimal anti-Gal activity.8,14,16,22
However, not all α-galactosidases are effective in removing α-Gal epitopes from xenograft tissues under physiological conditions. For example, recombinant human α-galactosidase A does not clear α-Gal epitopes in bovine pericardium under physiological conditions (pH 7–8) and works only under acidic pH conditions (pH 5.0) at the concentration (10 U/mL) that could cause tissue injuries and alter mechanical properties. 15 The Xanthomonas manihotis α-galactosidase removes α-Gal epitopes in decellularized bovine anterior cruciate ligament at an even higher concentration of 100 U/mL, an estimated enzyme cost of US$6300 per piece that is too high to be used for practical production. 16 For comparison, Bacteroides thetaiotaomicron α-galactosidase removes α-Gal epitopes in porcine heart valve under physiological conditions (pH 7.2) at the activity of 0.1 U/mL, which is 100–1,000 times more efficient than human α-galactosidase A. 21 The Coffea arabica α-galactosidase eliminates α-Gal epitopes in porcine skin at the activity of 0.05 U/mL with the cost less than US$7 per 100 cm2, despite of the fact that skin tissue has a very large number of α-Gal epitopes. 18
The present study has investigated the effectiveness of two different α-galactosidases, and the mechanism that leads to their different efficacies in eliminating the matrix-bound α-Gal epitopes in porcine acellular dermis matrix (pADM). Our study revealed that the accessibility of the enzyme to α-Gal epitopes and tissue form had significant influence on enzymic treatment. The study offers valuable insight into tissue process and enzyme selection for the efficient α-Gal antigen reduction in xenogeneic grafts or tissue scaffolds.
Method
Decellularization of porcine dermis and micronization
Porcine hides (5–8 mm thick) were leveled by a mechanical splitter to remove 0.5-mm epidermal layer and subcutaneous adipose tissue. The dermis tissue of uniform thickness (0.8–1.0 mm and/or 1.8–2.0 mm) was washed in purified water and surface-sterilized in 0.1% per acetic acid solution. Disinfected dermis was aseptically processed to yield a porcine acellular dermal matrix (pADM) via decellularization in 1.0% Triton ×-100 solution for 20 h. The pADM was washed four to five times with 0.9% sodium chloride solution to remove the detergent before α-galactosidase treatment. In order to investigate the effect of tissue forms (i.e., intact dermal matrix vs. microparticles) on the efficiency of xenogenic antigen reduction, the intact pADM was freeze-dried and then micronized by a cryo-mill with liquid nitrogen to tissue microparticles at different sizes. The size distribution and specific surface area of pADM microparticles were measured in 1% aqueous suspension by dynamic light scattering (Mastersizer 2000, Malvern Panalytical Ltd, Shanghai, China).
α-Galactosidase treatment
Two α-galactosidases were comparatively analyzed as follows: one extracted from green beans of Coffea arabica (Sigma-Aldrich, Inc., Saint Louis, Missouri, USA) and another purified from the fermentation solution of Aspergillus niger (Weikeqi Biological Technology Co., Chengdu, Sichuan, China). The purity of both enzymes was tested greater than 98% using the high-performance liquid chromatography method. Coffea α-galactosidase had a molecular weight of 37 kD with the optimal activity in pH 6.5 at 25°C, whereas Aspergillus α-galactosidase had a molecular weight of 108 kD with the optimal activity in pH 4.2 at 25°C. Due to different properties, the enzymic efficacy was first evaluated with p-nitrophenyl α-
Removal of matrix-bound α-Gal epitopes
The intact pADM and pADM microparticles were rehydrated in 100 mM phosphate buffer (pH 6.5), and α-galactosidase was added into solution to remove the matrix-bound α-Gal epitopes for 36 or 48 h. The ratio of solution volume to tissue weight was 25 mL/g. Enzymic activity tested was at the range of 0–2000 U/L, where one unit of enzymic activity was defined as the hydrolyzation of 1.0 µmole of p-nitrophenyl α-
Detection of matrix-bound α-Gal epitopes
The effect of decellularization and α-galactosidase treatment on α-Gal epitope removal was evaluated both qualitatively and quantitatively. For the qualitative detection, immunohistological chemistry (IHC) analysis was performed on cryo-sections by staining with the biotinylated-GS-IB4 lectin (Sigma-Aldrich, Inc., Saint Louis, Missouri, USA) (20 mg/mL) before stain color development in diaminobenzidine solution.18,25 Untreated porcine dermis and human dermis were used as positive control and negative control, respectively. To compare the effectiveness of α-galactosidase treatment, IHC slides were evaluated by three trained persons independently. The staining intensity was given by referencing the untreated pADM positive control (100% staining) and the human dermis negative control (0% staining). For the quantitative detection of residual α-Gal epitopes in decellularized and α-galactosidase-treated pADM samples, the inhibition ELISA assay with M86 monoclonal antibody was used, 26 and the protocol was based on the original Galili’s method.18,27 Tests were done with the α-Gal antigen quantitative detection kit per the manufacturer’s instruction (Beijing Sanyao Science and Technology Co., Beijing, China). Before tests, intact pADM samples were ground in liquid nitrogen to 25–100 μm microparticles.
Experiment
The pH difference for the optimal enzyme activity
Coffea α-galactosidase and Aspergillus α-galactosidase had different optimal pH for enzyme activity (Fig. 1). Coffea α-galactosidase had the optimal pH 6.5, whereas Aspergillus α-galactosidase had the optimal pH 4.2. For the reason that both enzymes worked well in the close to neutral pH 6.5, the effects of α-galactosidases and tissue forms on the removal of matrix-bound α-Gal epitopes were investigated in the pH 6.5 phosphate buffer.
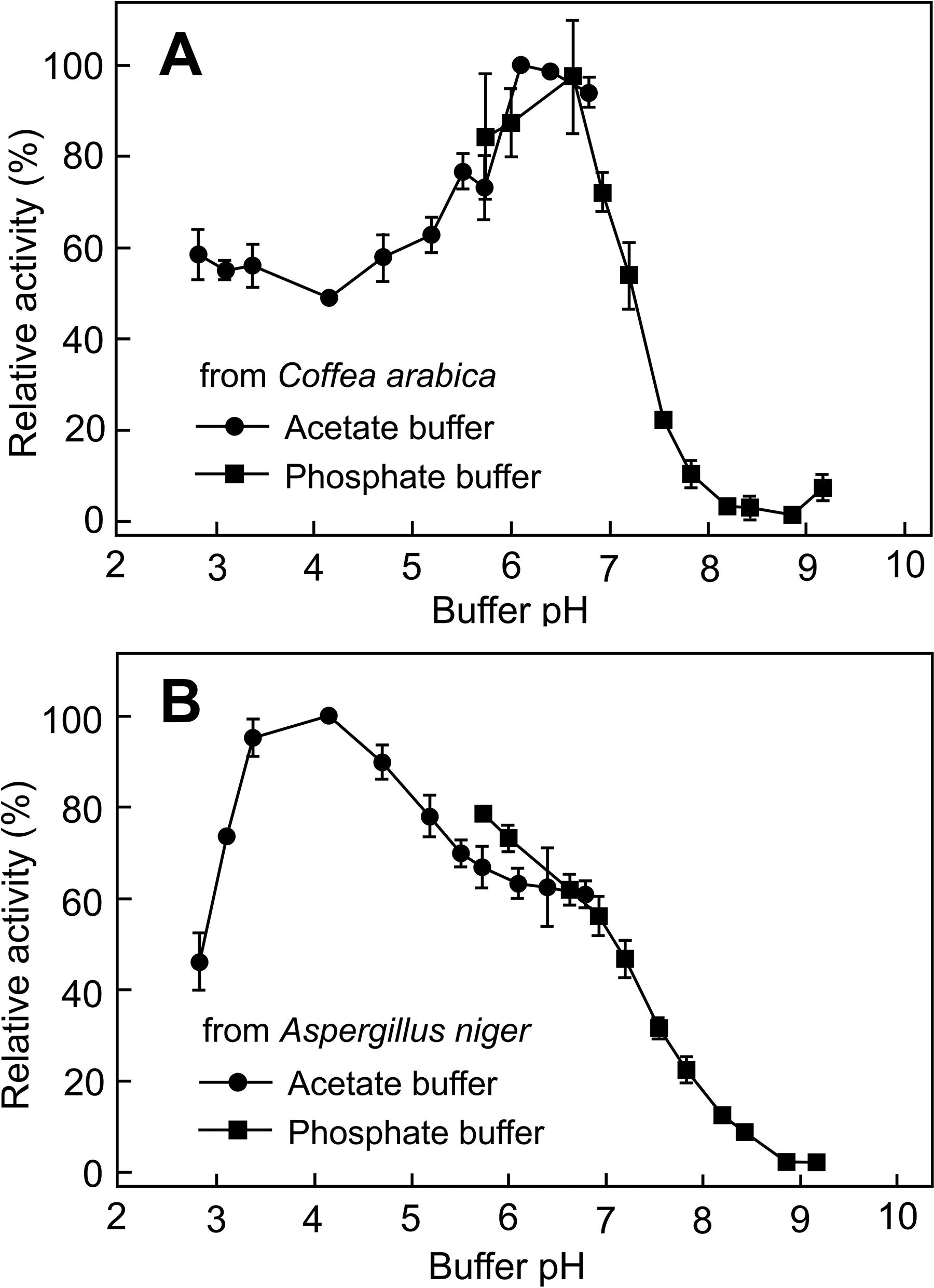
The effect of pH on the α-galactosidase activity.
Matrix-bound α-Gal epitopes in pADM
The total number of α-Gal epitopes in the unprocessed porcine dermis was determined to be 0.95 × 1015 ± 0.17 × 1015 counts/mg (n = 3 animals). Decellularization reduced the number of α-Gal epitopes in pADM to 0.19 × 1015 ± 0.04 × 1015 counts/mg, indicating the removal of α-Gal epitopes by 80.3 ± 4.4% (Fig. 2). The remaining α-Gal epitopes in pADM could not be further reduced via additional decellularization washes, even if pADM was micronized before additional extraction or washes. IHC analysis with the biotinylated-GS-IB4 lectin showed more or less even staining over the entire extracellular matrix of the pADM cross-section (Fig. 3A), suggesting that the remaining α-Gal epitopes after decellularization were matrix-bound. The content of residual DNA in pADM was determined to be ∼1 ng/mg tissue and represented 99.6% removal, also demonstrating that the remaining 19.7% α-Gal epitopes in pADM was not resulted from incomplete decellularization.
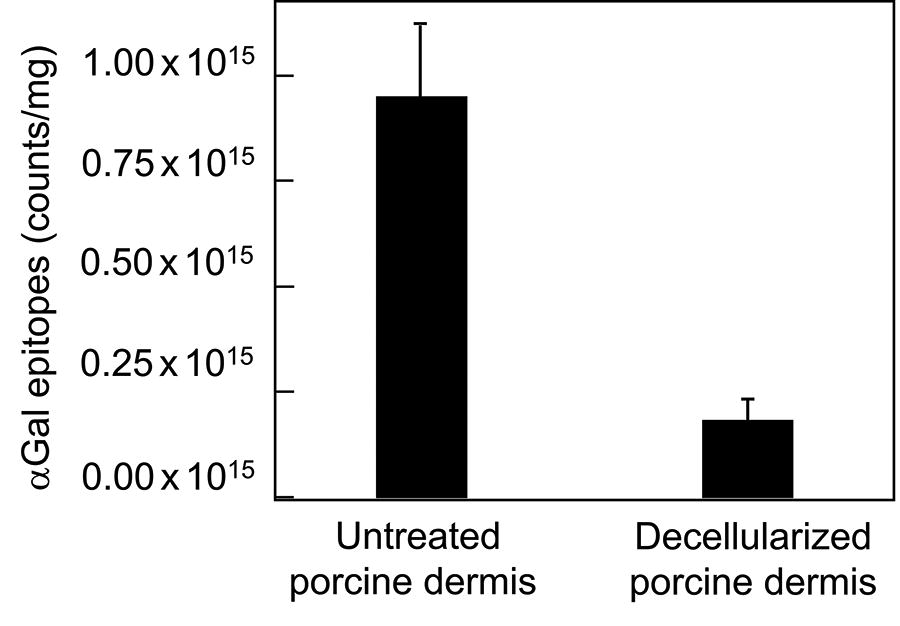
The number of α-Gal epitopes in the untreated porcine dermis and porcine acellular dermal matrix (pADM). Data were mean ± standard deviation (n = 3 animals).

The removal of matrix-bound α-Gal epitopes in porcine acellular dermal matrix (intact pADM) by Coffea or Aspergillus α-galactosidases.
Removal of matrix-bound α-Gal epitopes by α-galactosidases
Intact pADM samples at two different thickness ranges were used to investigate the effect of α-galactosidases on matrix-bound α-Gal epitopes. While both Aspergillus and Coffea α-galactosidases hydrolyzed the terminal alpha-galactosyl moiety from oligosaccharides (Fig. 1), they differed significantly in eliminating the matrix-bound α-Gal epitopes in the pH 6.5 phosphate buffer. Coffea α-galactosidase significantly reduced the number of matrix-bound α-Gal epitopes at 50 U/L, but the Aspergillus α-galactosidase had no effect even with more enzyme being added to 2000 U/L (Fig. 3). The qualitative detection by GS-IB4 lectin binding showed the reduced staining intensity between unprocessed porcine dermis and Coffea α-galactosidase-treated pADM (Fig. 3A, D), but the similar staining intensity between unprocessed porcine dermis and Aspergillus α-galactosidase-treated pADM (Fig. 3A, C). Aspergillus α-galactosidase also did not reduce the number of matrix-bound α-Gal epitopes in other pH and buffer conditions even when its enzymic activity was increased to 5000 U/L (Fig. 4). The number of remaining matrix-bound α-Gal epitopes after the Aspergillus α-galactosidase treatment was not significantly different from the untreated pADM material.

The number of matrix-bound α-Gal epitopes in porcine acellular dermal matrix (intact pADM) after the Aspergillus α-galactosidase (5,000 U/L) treatment in different pH and buffers for 48 h. The acetate buffer was used for pH in 3.1–5.5, whereas the phosphate buffer was used for pH in 6.8–9.2. Data were mean ± standard deviation (n = 3).
The effect of pADM micronization
To test whether the inability of Aspergillus α-galactosidase was due to the high density of intact pADM that resulted in the inaccessibility of α-Gal epitopes by the Aspergillus α-galactosidase, the intact pADM was ground in liquid nitrogen into microparticles before the α-galactosidase treatment. As pADM was ground into the increasingly smaller microparticles, the number of the exposed α-Gal epitopes increased steadily until the sizes of microparticles was reduced to less than 100 µm (Fig. 5A). Micronization greatly increased the specific surface area of pADM (Fig. 5B), exposing the matrix-bound α-Gal epitopes to be readily accessible by α-galactosidase molecules. In micronized pADM, both Coffea α-galactosidase and Aspergillus α-galactosidase were equally effective in reducing the number of matrix-bound α-Gal epitopes (Fig. 5C).

The removal of matrix-bound α-Gal epitopes in micronized porcine acellular dermal matrix (micronized pADM) by Coffea or Aspergillus α-galactosidases in pH 6.5 and phosphate buffer.
Discussion
The use of α-galactosidase to remove the matrix-bound α-Gal epitopes in xenografts was reported for more than two decades,8,13–23 but the method has not been commonly utilized in the process of manufacturing regenerative tissue products despite of its many advantages over acidic, alkaline, and/or cross-linking treatments. The technical issues involve the selection of suitable enzymes for various xenograft materials and the high cost of commercially available enzymes. Animal tissues used to make implantable medical devices vary in density, form, shape, and structure, including artery, bone, cartilage, dermis, fascia, heart valve, ligament, pericardium, tendon, vein, and others. Although all α-galactosidases can hydrolyze the terminal alpha-galactosyl moiety from soluble oligosaccharides, glycolipids, and glycoproteins, in practice one α-galactosidase that is effective in eliminating the matrix-bound α-Gal epitopes in a certain tissue may not necessarily work in another tissue. Different α-galactosidases have different sizes, molecular structures, selective substrates, and optimal working conditions, which may not be compatible with the tissue processing conditions for a particular medical device. In the present study, we discovered another important factor, the accessibility of α-galactosidase to the matrix-bound α-Gal epitopes in xenografts, as demonstrated by the effect of tissue forms (intact pADM vs. microparticle) (Figs. 3 and 5). This finding points out a rational way to develop the cost-effective process method for efficient α-Gal epitope removal via the selection of suitable α-galactosidase according to the xenograft’s tissue structures (e.g., localization of α-Gal epitopes, matrix porosity, enzyme size, molecular structure, and diffusibility within tissue matrix).
Presently there are several commercially available α-galactosidases, with the genes originated from organisms Aspergillus niger (35–159 kD), Coffea arabica (35–72 kD), Bacteroides fragilis (55–106 kD), Bacteroides thetaiotaomicron (48–83 kD), Homo sapiens (human) (23–51 kD), and Xanthomonas manihotis (∼69 kD). The size and structure of these enzymes vary greatly. The present study showed that the small Coffea arabica α-galactosidase (37 kD) could eliminate the matrix-bound α-Gal epitopes in highly dense intact pADM, whereas the large Aspergillus niger α-galactosidase (108 kD) had no effect without micronization to expose the matrix-bound α-Gal epitopes. Coffea arabica α-galactosidase was also reported to be effective in removing α-Gal epitopes from anterior cruciate ligament, aortic valve, and pericardium.16,19 A search on the Uniprot database showed the existence of smaller α-galactosidases (20–25 kD), which would become potentially better and more cost-effective enzyme choice.
In conclusion, decellularization of xenografts does not remove the matrix-bound xeno-reactive α-Gal epitopes. The present study showed the effectiveness of different α-galactosidases and the effect of tissue forms in removing the α-Gal epitopes. The results of the current investigations could be implemented for experimental, preclinical, and/or clinical research focused on pig-to-human transplantation of cutaneous/subcutaneous xenograft-based bioprostheses intended for tissue engineering as well as reconstructive and regenerative medicine of dermo-integumentary system in patients afflicted with skin diseases, lesions, injuries, burns or postoperative losses.28–32 Our newly completed clinical study with 145 burn patients has indeed demonstrated that the α-Gal epitope-reduced porcine dermal matrix adhered to the wound bed well, recellularized, revascularized, and partially incorporated for patient skin regeneration (unpublished data).
Footnotes
Acknowledgment
The authors thank Beijing Ruijian Gaoke Biotechnology Co., Ltd. (Beijing, China) for providing tissue materials, enzymes, and other reagents for the study.
Authors’ Contributions
W.Q.S. designed the study and obtained research funding. Y.-Y.Z., Y.-T.T., and S.-L.H. performed experiments. Y.-Y.Z. and W.Q.S. participated in data analysis, interpretation, and writing of the article.
Disclaimer
The authors declare that Beijing Ruijian Gaoke Biotechnology Co., Ltd. (Beijing, China) provided tissue materials, enzymes, and other reagents for the study, but the company did not interfere with the analysis and/or interpretation of the data.
Disclosure Statement
Y.Z. and W.Q.S. were the graduate student and the graduate advisor (professor), respectively, of University of Shanghai for Science and Technology. Y.T. and S.H. were employees of Beijing Ruijian Gaoke Biotechnology Co., Ltd. (Beijing, China). Publishing this article did not lead to any financial benefit to any authors.
Funding Information
The authors acknowledge funding from the Municipal Commission of Science and Technology (Beijing, China) under the grant number Z181100002218001.