
Abstract
Advanced tissue-engineered respiratory models are essential for studying drug or cosmetic toxicity, infection biology and xenobiotic metabolism. Here, we investigated a polyamide 6 (PA6)-based electrospun stromal scaffold as a substitute for porcine-derived small intestinal submucosa (SIS) to build human airway mucosa tissue models at the air-liquid interface. We demonstrate that the porous PA6 scaffold supports extracellular matrix production by human nasal fibroblasts and facilitates the complete differentiation of respiratory epithelial cells to the mucociliary phenotype. These models reduce reliance on animal-derived materials, improve reproducibility, and minimize potential interference from animal-derived antigens and pathogens. Both PA6- and SIS-based models promote fibroblast migration, epithelial differentiation, and the expression of key xenobiotic metabolizing enzymes. They exhibit comparable epithelial barrier integrity and susceptibility to influenza A virus infections. These findings establish PA6 scaffolds as a suitable, animal-free alternative to the SIS to build human airway mucosa tissue models.
Impact Statement
To minimize the usage of animal-components to build tissue-engineered models of the human respiratory mucosa, we replaced the porcine-derived small intestinal submucosa (SIS) by a polyamide 6 (PA6)-based electrospun scaffold. The PA6 scaffold supports extracellular matrix production by human nasal fibroblasts and facilitates the differentiation of respiratory epithelial cells to the mucociliary phenotype. SIS- and PA6-based respiratory tissue models exhibit comparable epithelial barrier integrity, abundance of xenobiotic metabolizing enzymes, and susceptibility to influenza A virus infections. These findings establish PA6 scaffolds as a suitable, animal-free alternative to SIS for generating human airway mucosa tissue models.
Introduction
During the COVID-19 pandemic, animals and cell culture models were used to identify therapeutic drugs for disease treatment. It became evident that antiviral effects of direct-acting compounds, such as hydroxychloroquine, were dependent on the model that was used: Whereas in animal and human cell line cultures, hydroxychloroquine was described to have antiviral activity regarding severe acute respiratory syndrome coronavirus type 2, this substance did not affect viral infection either in primary respiratory human air-liquid interface (ALI) cultures or in nonhuman primates.1–3 Clinical studies demonstrated that hydroxychloroquine did not affect COVID-19 progression.4,5 Thus, in this case, differentiated human primary respiratory tissue models were more predictable as cell line cultures and were in line with data obtained in human patients. Although it cannot be concluded that human primary respiratory tissue models are accurate and predictive in all preclinical studies, they are, together with precision-cut lung slices, regarded as more suitable and in vivo-like than cell line-based cultures to address respiratory research questions in biomedicine. 6 Moreover, three-dimensional (3D) engineered tissue models often outperform animal tests and were validated as alternatives for toxicity screening assays in drugs and cosmetic products, thus, reducing the need for animals.7–9 If scientifically valid options are available, animal testing is no longer mandatory for the Food and Drug Administration.10–12
Respiratory tissue models including ALI cultures, organoids, and precision-cut lung slices, have been applied by many research groups to replicate the human airways in vitro. Major and minor airway epithelial cell populations, such as basal, secretory, ciliated, brush cells, and ionocytes were identified in human respiratory ALI systems, highlighting their similarity to the respiratory epithelium in vivo, whereby culture conditions have to be considered.13,14 Respiratory tissue models replicate key epithelial features, such as barrier integrity, the mucociliary phenotype, and pathogen susceptibility, which are essential for studying interactions with external environmental factors. Moreover, they imply xenobiotic metabolizing enzymes (XMEs), which are essential for the biotransformation of inhaled compounds and chemosensory tissue protection.
To study respiratory infections or xenobiotic metabolism, transwell-based ALI models have been used commonly.15–17 To build more complex tissue models representing the epithelial layer and the adjacent fibroblast-loaded connective tissue, appropriate 3D scaffolds are needed. Several 3D engineered mucosal tissue models are based on scaffolds consisting of animal-derived components, such as hydrogels or porcine small intestinal submucosa (SIS).18–21 A disadvantage here is that such models contain animal proteins in varying composition, which make them less human-like and do not support advancing the research field of nonanimal methods. To reduce animal-derived compounds and associated variation, we investigated novel electrospun synthetic polyamide 6 (PA6) fiber scaffolds as a replacement for the SIS. PA6 is biocompatible, suitable for electrospinning, and it was shown previously that highly porous PA6 scaffolds can be used to build human skin equivalents. 22 Based on this knowledge, we assessed the suitability of the PA6 scaffolds for building nasal mucosa tissue models by comparing their tissue morphology, cell type-specific marker and XME abundance, barrier properties, and susceptibility to influenza A virus with SIS-based scaffolds.
Methods
Donor information
Primary nasal epithelial cells and fibroblasts were obtained from patients undergoing surgery at the University Hospital Würzburg (Table 1). Informed consent was obtained beforehand. The institutional ethics committee on human research of the University of Würzburg approved this study (vote 179/17).
Anonymized Donor Information
f, female; m, male.
Electrospinning of highly porous PA6 scaffolds
Electrospinning was performed as previously described. 22 Briefly, a 12% (m/v) solution of PA6 (18110, Sigma-Aldrich, Steinheim, Germany) in 1,1,1,3,3,3-hexafluoro-2-propanol (105228, Sigma-Aldrich) was spun on a coaxial electro spinner. The needle tip was positioned 15 cm from the rolling receiver, and electrospinning was performed at 10 kV onto a rotating collector (surface speed of 0.9 m s−1) covered with aluminum foil. Every 3 min, the receiver was sprayed with ethanol and sprinkled with the porogen NaCl, having a particle diameter between 40 and 56 µm. This process was repeated 30 times. Afterwards, the scaffold, including aluminum foil, was removed and stored at room temperature.
Cell isolation and culture
Cell culture was carried out under standard conditions (37°C, 5% CO2). Primary nasal epithelial cells and fibroblasts were isolated from biopsies as previously described.18,23 Epithelial cells were cultured in airway epithelial cell growth medium (AECG) (PB-C-MH350-0099, PeloBiotech, Planegg, Germany). Fibroblasts were grown in Dulbecco’s Modified Eagle Medium (DMEM) (61965–026, Thermo Fisher Scientific, Waltham, USA) supplemented with 10% fetal calf serum (P150508, PAN biotech, Aidenbach, Germany) up to 90% confluency.
PA6 scaffold preparation and fibroblast seeding
PA6 scaffolds were cut into squares (2.5 × 2.5 cm), submerged in water and mounted on cylindrical cell crowns in pairs. The mounted scaffolds were washed overnight in deionized water, autoclaved, submerged in H2O, and transferred to phosphate-buffered saline (PBS). The cell crowns were transferred to a 6-well plate filled with 3 mL PBS per well. The PBS was replaced by DMEM supplemented with 100 µM ascorbic acid 2-phosphate (A8960, Sigma-Aldrich) for 2 h at 37°C. A total of 50,000 fibroblasts were seeded in 500 µL DMEM supplemented with 100 µM ascorbic acid 2-phosphate. The fibroblasts were cultured for 14 days before epithelial cells were seeded. For the analysis of collagen production, fibroblast-loaded PA6 scaffolds were cultured for 3, 7, 14, and 21 days in DMEM supplemented with 500 µM ascorbic acid 2-phosphate before fixation and analysis.
SIS preparation and fibroblast seeding
Porcine jejunum was obtained as described previously. 23 In brief, explanted jejunum of heparin-treated pigs was washed, cut into segments, decellularized using sodium deoxycholate (Sigma-Aldrich), treated with DNase I (Roche, Basel, Switzerland), and γ-sterilized using over 25 kGy (BBF steriXpert, Rommelshausen, Germany). The SIS was then cut open, the serosa was removed, and it was cut into squares (2.5 × 2.5 cm). The squares were mounted on cylindrical cell crowns, transferred to PBS, and incubated in DMEM for 2 h at 37°C. A total of 50,000 fibroblasts were seeded in 500 µL DMEM and cultured for one day before epithelial cell seeding. Animal research was performed according to German law and institutional guidelines approved by the District of Unterfranken Ethics Committee, Würzburg, Germany (approval number 55.2-2532-2-256). The animals received proper attention and human care in compliance with the Guide for Care and Use of Laboratory Animals published by the National Institute of Health (NIH publication no. 85e23, revised 1996) and as approved by the Institutional Board of Animal Protection. Organ explantation complied with the German Animal Protection Law (§4 Abs.3), and the institute’s animal protection officer regularly informed the responsible authorities.
Tissue model generation
To build full-thickness airway mucosa tissue models using PA6 and SIS scaffolds, the medium of the fibroblast-loaded scaffolds was switched to a mixed medium (DMEM:AECG 1:1) and 250,000 nasal epithelial cells were seeded in 500 µL AECG on the apical side. After one day of submerged culture, the models were airlifted and incubated until the mucociliary phenotype developed. Cells from the same donor in the same passage were used to build PA6- and SIS-based airway mucosa tissue models.
Collagen quantification
To quantify the collagen concentration in fibroblast-loaded PA6 scaffolds 3, 7, 14, and 21 days after cell seeding, the samples were hydrolyzed with 6 M HCl and processed using a Total Collagen Assay (QuickZyme Biosciences, Leiden, The Netherlands) according to the manufacturer’s instructions. The samples were measured at 570 nm in a microplate reader (Infinite 200 PRO NanoQuant Microplate Reader, Tecan, Männedorf, Switzerland).
Histological analysis
For histological analysis, tissue models were fixed in Roticlear® Histofix (P087, Carl Roth, Karlsruhe, Germany), embedded in paraffin (6642.6, Carl Roth), and sectioned to a thickness of 5 µm using a microtome (RM2255, Leica, Wetzlar, Germany). The samples were deparaffinized in xylene (9713.3, Carl Roth, Karlsruhe, Germany) and rehydrated in ethanol with decreasing concentrations. Picro-Sirius red and hematoxylin/eosin (HE) staining were performed following standard protocols. For immunofluorescent staining, samples were boiled at pH 6 for 20 min to retrieve antigens obscured by paraffin embedding. Unspecific antibody bindings were blocked using 5% bovine serum albumin in PBS containing 0.01% Triton X-100 for 1 h. The samples were incubated with primary antibodies (Table 2) overnight at 4°C. The samples were washed in PBS with 1% Tween 20 (PBST) and incubated with secondary Alexa Fluor™ 647 conjugated donkey anti-mouse (dilution 1:400, A31571, Thermo Fisher Scientific) and Alexa Fluor™ 488 conjugated donkey anti-rabbit (dilution 1:400, A21206, Thermo Fisher Scientific) antibodies for 1 h, washed with PBST, and covered with coverslips using Fluoromount-G™ with 4′,6-diamidino-2-phenylindole (DAPI, 00–4959-52, ThermoFisher). Images were captured with a Keyence fluorescence microscope (BZ-X810, Keyence, Neu-Isenburg, Germany).
List of Primary Antibodies
AKR, Aldo-keto-reductase; ALDH, Aldehyde dehydrogenase; CK, Cytokeratin; DCXR, Dicarbonyl/L-xylulose reductase; GST, Glutathione transferase; MUC, Mucin.
FITC-Dextran permeability assay
PA6- and SIS-based airway mucosa tissue models were incubated apically with 500 µL PBS at 37°C for 15 min. The apical supernatant was removed, and models were transferred to a new 6-well plate. Fluorescein isothiocyanate (FITC)-dextran (4 kDa, 46944, Sigma-Aldrich) was dissolved at 0.25 mg/mL in a preheated (37°C) mixed medium. The solution was sterile-filtered (0.22 µm), and 1 mL was applied apically to assess epithelial barrier properties. After 30 min, the models were transferred to new plates and washed with PBS. The remaining basal supernatant was transferred to black 96-well plates, and the fluorescence was measured using a microplate reader (absorption: 490 nm, emission: 525 nm). Unseeded scaffolds were used as a reference.
Viral infection
To investigate model-specific differences in influenza A virus (IAV) replication, SIS and PA6 models were built in quadruplicates. Two SIS and PA6 models were designated for RNA extraction, while two were intended for immunofluorescent staining. The models were washed and infected with 200,000 infectious units of IAV for 4 h, followed by three washes with PBS. Intracellular RNA was purified using the E.Z.N.A. Total RNA Kit I (R6834, Omega Bio-Tek, Norcross, USA). For viral genome quantification, the LightCycler® Multiplex RNA Virus Master kit (LightCycler® Multiplex RNA Virus Master Version 09, 07083173001, Roche, Basel, Switzerland), the LightMix® Modular Influenza kit (53-0101-96, TIB MOLBIOL, Berlin, Germany), and a LightCycler® 480 II (Roche) were used. Viral genomes were quantified with the cycler-specific software version 1.5.
Statistics
Statistical significance was calculated using a Kruskal–Wallis test and a Student–Newman–Keuls test for the collagen quantification and the FITC-dextran permeability assay. Statistics were done using a student’s paired t-test for influenza A infections. Tests were considered significant for p < 0.05.
Results
Biologization of the PA6 scaffold
To replace the porcine-derived SIS, we evaluated the suitability of a highly porous PA6 scaffold to support fibroblast migration, ECM formation, epithelial cell attachment and differentiation. Fibroblasts were seeded on a PA6 scaffold and cultured for 3, 7, 14, and 21 days. Within 21 days, fibroblasts showed progressive ECM development on the PA6 scaffold, as visualized by Picro-Sirius red staining. Fourteen and 21 days after fibroblast seeding, the qualitative results appeared similar (Fig. 1A). A quantitative hydroxyproline assay confirmed the substantial increase in collagen levels at day 14 (58.7 ± 37.1 µg/mL) and day 21 (96.2 ± 47.4 µg/mL), significantly higher than at day 3 (1.6 ± 2.8 µg/mL) (Fig. 1B). These findings established day 14 as the optimal time point for epithelial cell seeding.
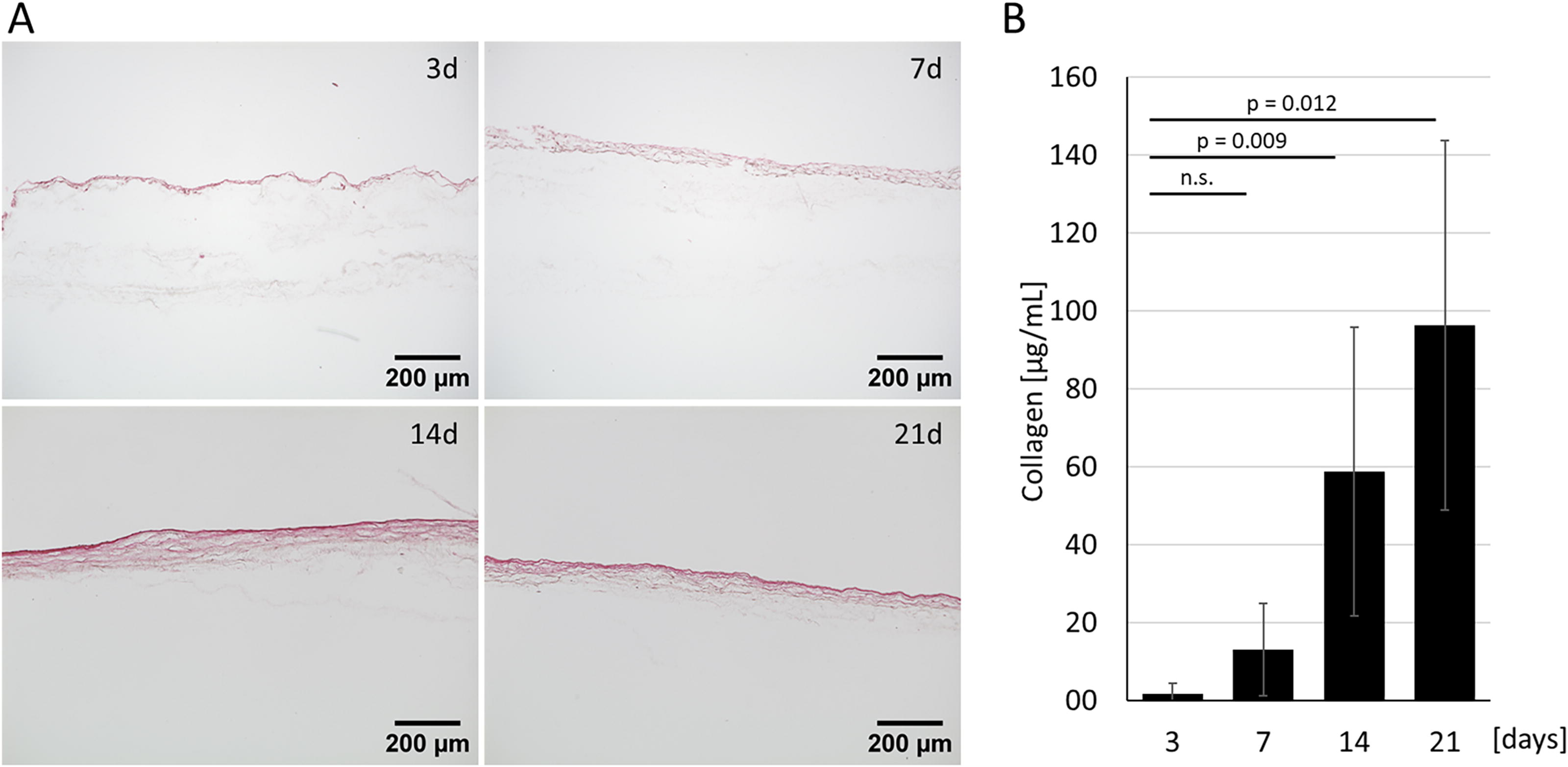
Human nasal fibroblasts biologize synthetic polyamide 6 (PA6) scaffolds.
SIS- and PA6-based respiratory models comparably express characteristic differentiation markers and XMEs
To assess the PA6 scaffold as a replacement for SIS, we built tissue models of human respiratory mucosa and analyzed marker expression using immunofluorescent staining. Fibroblasts and epithelial cells from the same donor in the same passage were used to build PA6- and SIS-based airway mucosa tissue models for comparative analysis. In both model types, epithelial cells differentiated to the mucociliary phenotype similarly. The models showed cytokeratin 5- (CK5) positive basal cells, acetylated tubulin-positive ciliated cells, and mucin 5AC- and 5B-producing cells along with the membrane-bound mucin 1 (Figs. 2 and 3). The presence of vimentin throughout the scaffold shows the localization of the fibroblasts. Additionally, the XMEs aldo-keto-reductase family 1 member B10, glutathione transferase A1, glutathione transferase P1, dicarbonyl/L-xylulose reductase, aldehyde dehydrogenase 1A1, and aldehyde dehydrogenase 3A1 were present, suggesting the metabolic activity of the epithelium. Thus, PA6-based models successfully replicated the mucociliary phenotype and the presence of selected XMEs found in SIS-based models (Fig. 3).

Immunofluorescent staining shows epithelial cell differentiation in small intestinal submucosa (SIS)-based respiratory tissue models. Airway epithelial marker include acetylated tubulin, cytokeratin 5, mucins 1, 5AC, 5B, and xenobiotic metabolizing enzymes Aldo-keto-reductase (AKR) 1B10, Glutathione transferase (GST) A1, Dicarbonyl/L-xylulose reductase (DCXR), GSTP1, Aldehyde dehydrogenase (ALDH) 1A1, and ALDH3A1.

Immunofluorescent staining shows epithelial cell differentiation in polyamide 6 (PA6) -based respiratory tissue models. Airway epithelial marker include acetylated tubulin, cytokeratin 5, mucins 1, 5AC, 5B, and xenobiotic metabolizing enzymes Aldo-keto-reductase (AKR) 1B10, Glutathione transferase (GST) A1, Dicarbonyl/L-xylulose reductase (DCXR), GSTP1, Aldehyde dehydrogenase (ALDH) 1A1, and ALDH3A1.
SIS- and PA6-based respiratory models have comparable barrier properties and are suitable for IAV infection studies
The FITC-dextran permeability assay is a well-accepted and suitable method to measure the barrier integrity of airway ALI models.18,24,25 Thus, the epithelial barrier integrity was assessed by investigating the permeability of 4 kDa FITC-dextran from the apical to the basolateral compartment (Fig. 4A). Scaffolds of both type that were loaded with fibroblasts only, showed a sharp drop in permeability by 12.5 ± 3.1-fold (SIS) and 13.7 ± 3.1-fold (PA6) compared to unseeded scaffolds. Adding epithelial cells and letting them differentiate at the ALI (full model) decreased the permeability 303.5 ± 136.3 (SIS) and 315.2 ± 162.3 (PA6) fold compared to unseeded scaffolds. PA6-based models achieved barrier properties equivalent to SIS-based models, demonstrating their functional comparability. HE staining underlines the intact epithelial morphology and shows that the epithelial layer is directly anchored to the underlying biologized scaffold (Fig. 4B).

To prove the suitability of PA6-based human airway mucosa tissue models for viral studies, we infected models based on both scaffold types with IAV and compared the viral gene expression. Immunofluorescent staining revealed similar infection rates in the epithelial layer independent of the scaffold used (Fig. 5A). These results were confirmed by quantification of viral RNA expression, revealing similar genome numbers confirming comparable gene expression in PA6- and SIS-based models (Fig. 5B).
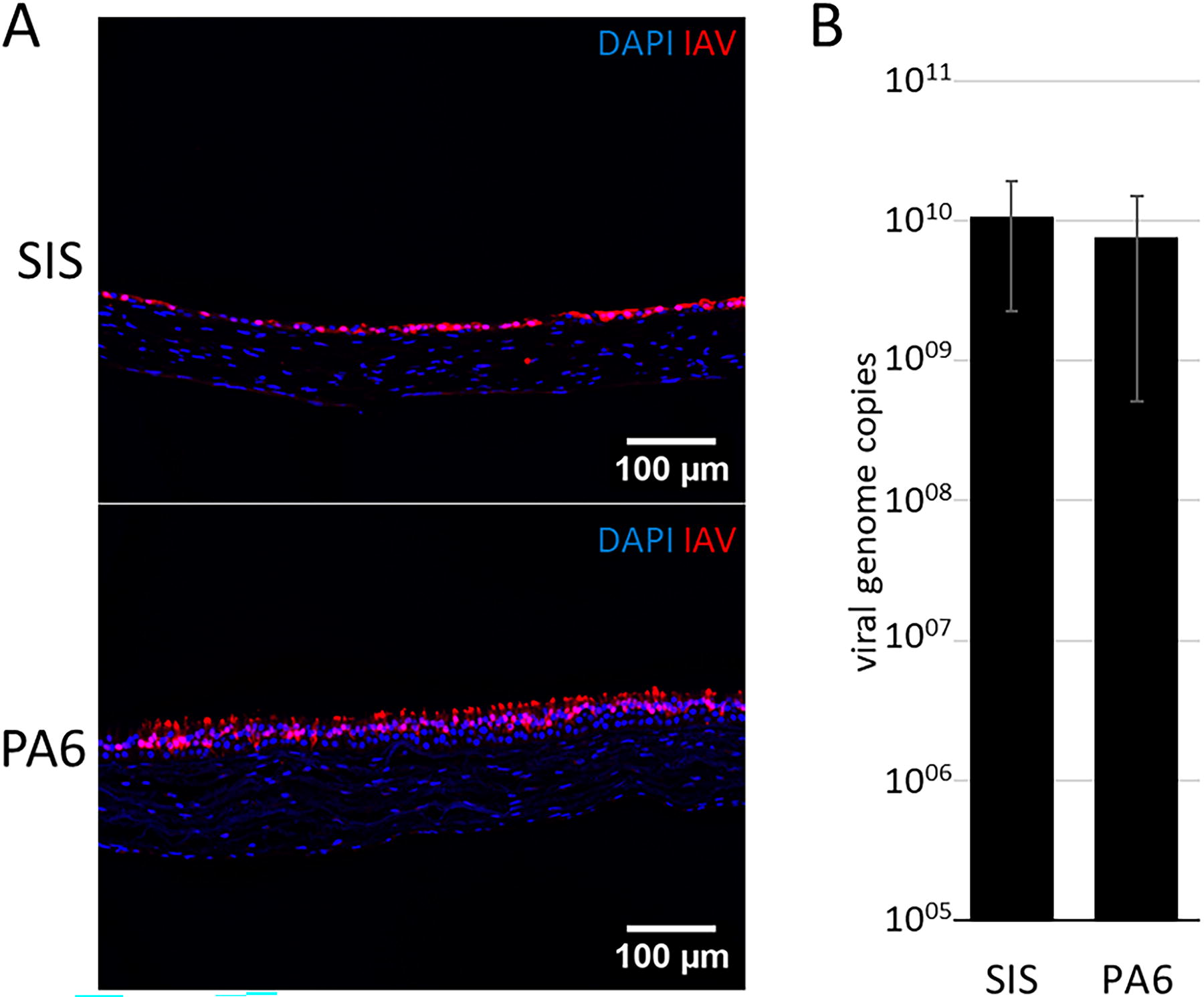
PA6- and SIS-based airway mucosa tissue models are equally susceptible to IAV.
Discussion
This study demonstrates the suitability of highly porous PA6 scaffolds to build complex human airway mucosa tissue models. It eliminates the necessity of animal-derived matrices while maintaining direct contact between the epithelium and the fibroblast-loaded connective tissue. Comparative analysis with models built using an SIS scaffold revealed similar tissue morphology, cell differentiation, epithelial barrier integrity, XME abundance, and IAV susceptibility.
PA6 is known for its biocompatibility, high availability, and suitability for electrospinning,26,27 which makes it a suitable polymer for scaffold generation. Furthermore, the stability against harsh solvents in tissue analysis and characterization, like paraffin embedding, brings the main advantage compared to other biocompatible polymers like polyesters, polystyrene, and polycarbonate. The synthesis approach, the ultrastructure, and the mechanical properties of highly porous PA6 scaffolds have been assessed in a previous study. 22 It is notable that the pure PA6 scaffold, which was not chemically modified with cell-adhesive sites, such as RGD peptides, 28 allowed nasal fibroblast migration, homogenous fibroblast distribution, proliferation, and ECM synthesis. In accordance with previous data on human dermal fibroblasts, 22 we assume that the defined porosity achieved by the repeating-spinning process in combination with adsorbed proteins from the cell culture medium are deciding parameters that enable fibroblasts to biologize the PA6 scaffold sufficiently. If electrospun PA6 scaffolds are too dense, fibroblasts will remain on the scaffold’s surface. 22 It can be assumed that ECM molecules and factors secreted by the fibroblasts, such as hepatocyte growth factor, support epithelial cell attachment and differentiation at the ALI, similar to SIS-based tissue models. 29 Even though the collagen concentration was higher 21 days after fibroblast seeding, 14 days of scaffold biologization was sufficient to facilitate epithelial cell attachment and differentiation. A shorter cultivation time is staff-, time-, and material-saving, which is why we chose day 14 as the optimal time point for epithelial cell seeding.
The usage of PA6 provides significant advantages over SIS through its synthetic nature. Instead of having a porcine extracellular matrix (ECM), the PA6 scaffold supports the synthesis of a human donor-specific ECM significantly contributed to by human nasal fibroblasts. This process supports epithelial cell attachment and differentiation while preventing porcine-derived contamination.22,30,31 The absence of animal-derived contaminants improves the reproducibility of the built models and reduces cross-species interference.32–35 Both scaffolds incorporate connective tissue that allows cell migration and interaction across the entire model.17,34 The direct contact of the epithelial cells to the connective tissue enhances model complexity, enabling further cell-cell communication, the formation of a physiological basal lamina, and the integration of other cell types, such as immune cells. As nanofibers can influence immune cell differentiation, these effects must be evaluated before conducting experimental studies.36–38 While our tissue model still relies on some animal-derived components, such as fetal calf serum or antibodies for immunofluorescent staining, ongoing efforts aim to replace them. In sum, PA6-based scaffolds are a highly suitable alternative to SIS, significantly advancing animal-free research, for example, in research on respiratory infection and nasal xenobiotic metabolism.
Authors’ Contributions
N.P., E.L., and H.O. performed the experiments. N.P., J.B., and M.S. wrote the article. A.F., A.S., and S.H. obtained informed consent, took care of patient data management, obtained human tissue specimens and edited the article. T.W. contributed to the synthetic scaffold. J.M.H., J.B., and M.S. designed the study. N.P., J.B., and M.S. discussed the data. M.S., S.H., and J.M.H. acquired the funding. All the authors consent to being accountable for the content of the work and approve the submitted version.
Footnotes
Disclosure Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding Information
This work was supported by the German Research Foundation (DFG, grant numbers HA 7315/4-1, STE 2442/6-1), and RTG 2157 “3D Tissue Models for Studying Microbial Infections by Human Pathogens”), and by the French National Research Agency (ANR) (grant number ANR-18-CE92-0018-01).