
Abstract
Hypothermia has been linked to beneficial neurologic outcomes in different clinical situations and its therapeutic value is considered important. For example, in asphyctic neonates and in patients with out-of-hospital cardiac arrest (with ventricular fibrillation as the initial cardiac rhythm), rapid installation of hypothermia has been reported to add substantial therapeutic benefits over nonthermal standard treatments. Yet, in other groups of patients in which the application of therapeutic hypothermia may be applied with clinical benefits, the optimization of therapy remains less straightforward, as the body possesses vigorous defense mechanisms to protect it from inducing hypothermia, that is, especially in conscious patients and/or in those in which the hypothalamus remains intact, such as stroke patients or patients who suffer a myocardial infarction or spinal cord injury. This overview summarizes the body's primary reactions to hypothermia and the defense mechanisms available or evoked. Then, clinically applicable ways to overcome these forceful cold defenses of the body are described to ensure both an optimal induction process for therapeutic hypothermia and maximal subjective comfort for these conscious patients.
Introduction
Thermosensation
Temperature information is detected within the skin by specialized sensory cells known as thermoreceptors. These cutaneous temperature sensors consist of highly specialized ion channels with characteristic temperature thresholds (specialized warm and cold receptors) that change their gating properties accordingly (Romanovsky, 2007). The signals from the peripheral thermosensors are then conveyed centrally by A∂ and C-fibers via thermoafferent pathways (Schepers and Ringkamp, 2010). In addition to these cutaneous cold- and warm-sensitive neurons, thermoreceptors are also present within central body (core) structures. Such deep-body sensors can be found in the esophagus, stomach, large intra-abdominal veins, and/or other organs. Thermal information from these various sites is gathered and partially integrated at multiple levels before reaching the hypothalamus, the central thermoregulatory center in mammals. It should be noted that thermoreceptors within the hypothalamus (and other parts of the brain), the spinal cord, deep thoracic tissues, deep abdominal tissues, and the skin surface each contribute roughly 20% of the total thermal input to the central regulatory system (Simon, 1974; Jessen and Feistkorn, 1984; Cheng et al., 1995). This physiologically weighted average reflects the thermoregulatory importance of these different tissues in the overall thermoregulatory process. However, it should also be noted that with regard to the skin surface, thermoreception, like other sensory modalities, has an unequal distribution throughout the body, prioritizing thermal information especially from the facial areas, the hands, and the feet over other regions. Although thermoreception in the cortex is not exactly represented as seen in the Penfield's homunculus (Penfield and Boldrey, 1937) for somatosensation (Fig. 1), there are similarities (Chatt and Kenshalo, 1977; Berman et al., 1998). Thus, knowledge of this characteristic (i.e., disproportionate central emphasis of certain body parts) from a clinical thermoregulatory perspective can be specifically employed to mislead (fool) the thermoregulatory system by providing overwhelming temperature information at these specific sites, as will be later discussed.
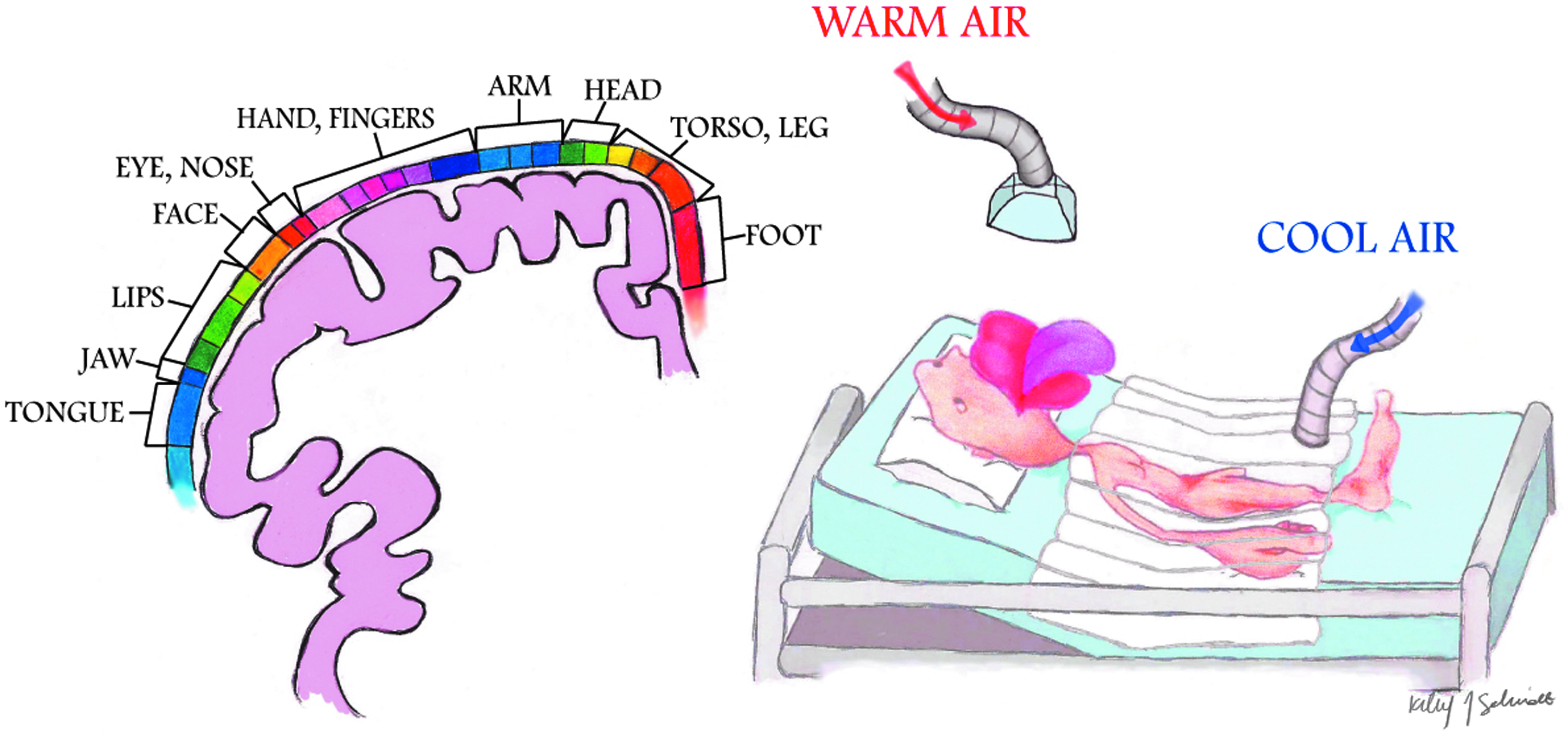
Penfield's homunculus for somatosensation; parts of the body with the highest tactile acuity are overrepresented on the cortex (illustration by Kiley Schmidt). Similarly, thermoreception has an unequal distribution throughout the body, especially with overrepresentation of the facial area. Chatt and Kenshalo (1977) found a similar cortical organization of temperature (both warm and cool stimuli) in humans to that of touch. Further, there is nonproportional contribution of thermal input into the control centers into the hypothalamus for shiver (see Mekjavic and Eiken, 1985; Iaizzo et al., 1999; Sweney et al., 2001), for example, you can suppress whole-body shiver effector mechanisms by focal warming of the face, hands, or feet.
Thermal Homeostasis or Thermoregulation
Under normal conditions, body temperature is tightly regulated (homeostasis), for example, even tighter than blood pressure or heart rate (Sessler, 2008). Similar to many other physiologic control systems governed by the central nervous system, the thermoregulatory center utilizes negative feedback mechanisms to minimize perturbations from preset “normal” values (Fig. 2). Thereby, the afferent thermal inputs from the skin surface, neuraxis, and deep tissues are integrated and compared with those within central (core) structures, that is, primarily the hypothalamus. The importance of the central structures for these homeostatic responses can be illustrated by observing the profoundly disturbed thermoregulation responses within brain-dead individuals or stroke victims, in whom the hypothalamic integrity becomes greatly affected (Wood et al., 2004). Nevertheless, sites other than those in the hypothalamus have considerable importance for thermoregulatory responses, for example, as within the spinal cord, where thermal information is substantially preprocessed from that being obtained from the periphery (Satinoff, 1978).
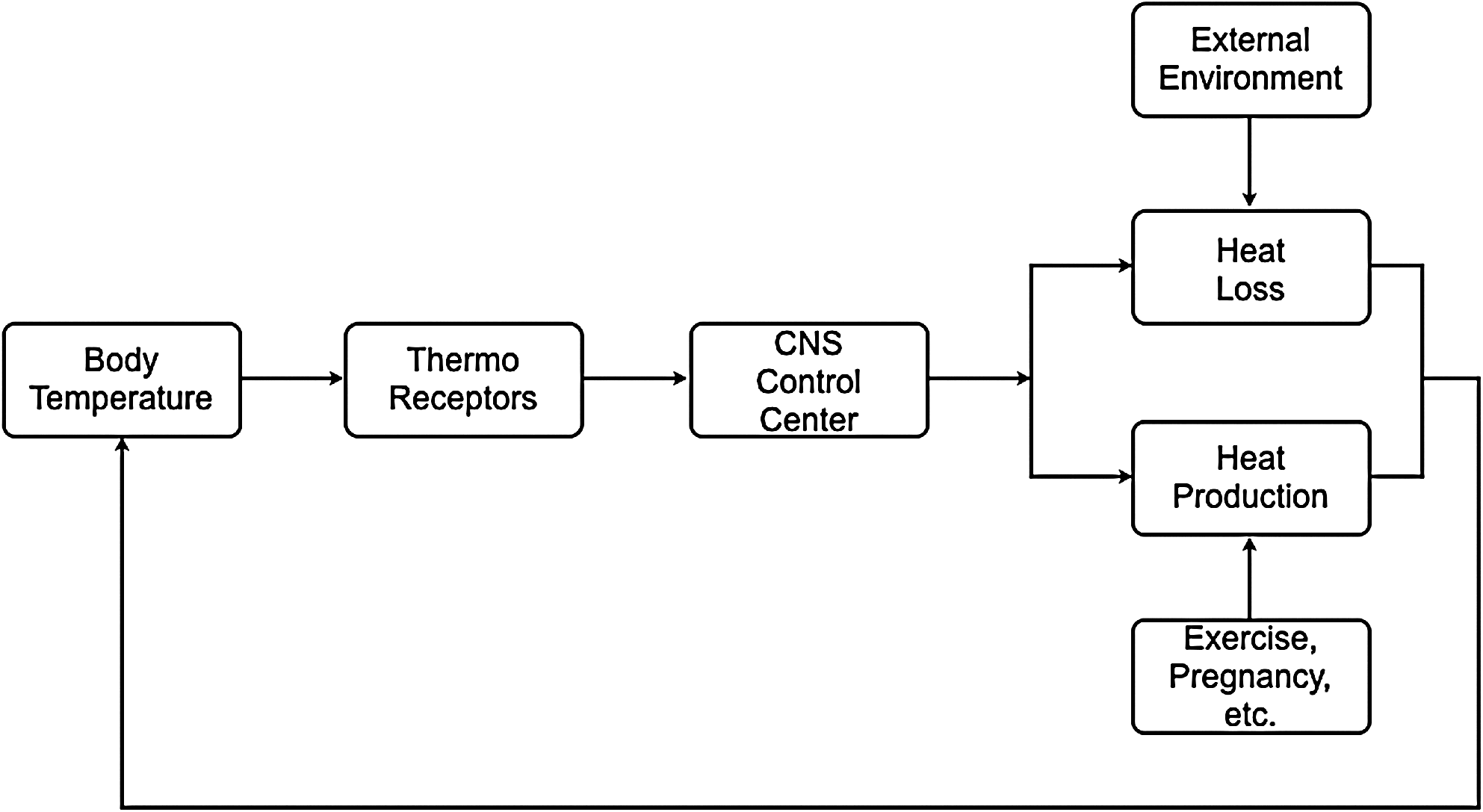
Negative feedback of the thermoregulatory center.
In general, in the case that an integrated thermal input exceeds one of the threshold temperatures for heat and cold, counterregulatory autonomic mechanisms come into effect. Normally, in humans, core temperature is tightly maintained near a set point of 37°C, as virtually every cell in the human body functions best at this “optimal” temperature, with the exception of immune cells, which are known to operate most efficiently in febrile conditions (Kluger et al., 1975; Roberts and Sandberg, 1979; Jampel et al., 1983; Rowsey, 1997), this being associated with reduced pathogen viability at these higher body temperatures (Mackowiak, 1981). In general, during a given period of the day, core temperature is controlled within 0.2°C of this set point. Yet, it is also known that this small, so-called interthreshold range widens as we age; this control is less affected by either gender or body type (somatotypes) (Lopez et al., 1994; Vassilieff et al., 1995). In the case when even a slight deviation in core temperature is detected, this tight interthreshold range is surpassed and effector mechanisms will automatically increase or decrease an individual's metabolic heat production and/or heat loss or gain from the environment (Fig. 3). Although both central body (brain, spinal cord, heart, lungs, and thorax) and peripheral parts (limbs and skin) theoretically share a common “ideal” functional set point of 37°C, it is only the core compartment that is held within the tight (minimal) temperature range, which is readily counteracted by defensive measures. In other words, the temperature in the peripheral compartment, even under normal conditions, can vary greatly from the hypothalamic set point with minimal consequences. This can be readily illustrated during normal conditions, where it is common that the temperature in the peripheral compartment is 2°C–4°C lower than the core temperature. Nevertheless, depending on environmental and metabolic (e.g., hormonal or exercise) circumstances, this difference can vary greatly.
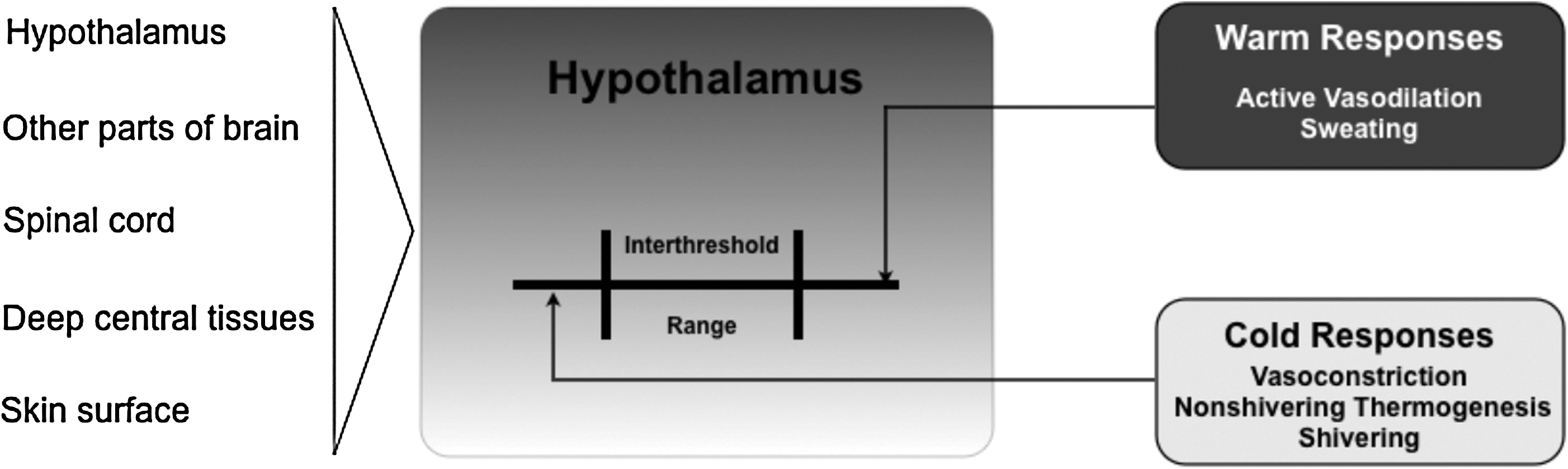
Thermoregulatory system.
Normal Thermoregulatory Defense Mechanisms
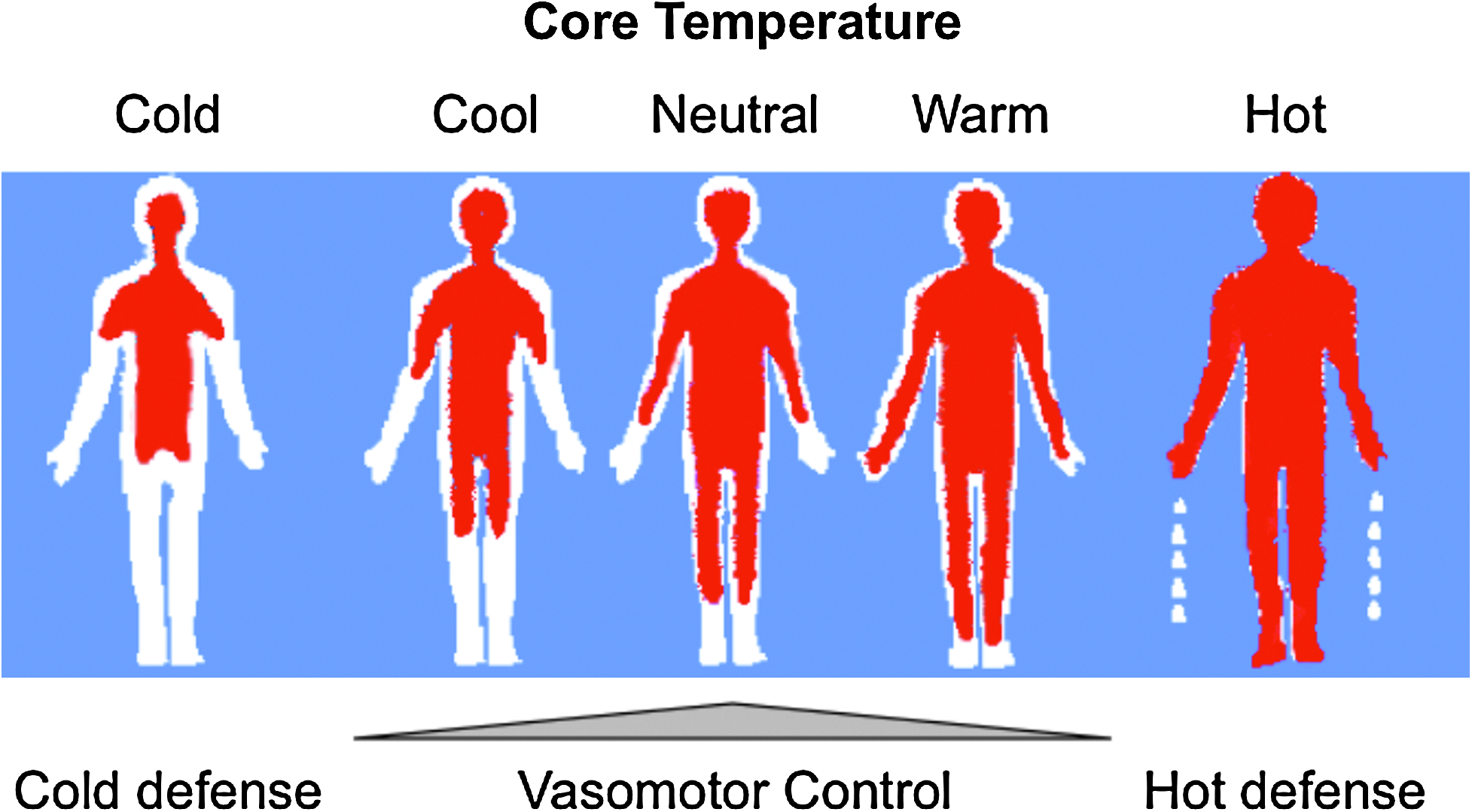
Effector mechanisms can roughly be divided into behavioral and hypothalamically controlled autonomic responses. The behavioral component of the response (e.g., protection by clothes, shelter, shade, or air conditioning) is most important relative to humans. Nevertheless, it should be noted that as one becomes a patient, these controls are taken over by one's healthcare provider. Concerning the effector response governed by the autonomic nervous system (hypothalamus), precapillary vasodilation and sweating are active in the thermoregulatory response to heat, whereas arteriovenous shunting or induced vasoconstriction and shivering are primarily activated in response to cold (Fig. 4). In general, with a considered normal set point of 37°C, cutaneous vasoconstriction is initiated at a core temperature of around 36.5°C and significant shivering at around 35.5°C. The thermoregulatory arteriovenous shunts are located primarily in acral regions, that is, the fingers and toes (Hales et al., 1985). It should be noted that although most blood vessels constrict in response to regional hypothermia, these arteriovenous shunts are relatively resistant to local temperature changes and seem to be mainly controlled by central mechanisms (Sessler, 2008). More specifically, these specialized thermoregulatory vessels are under α-adrenergic control; at typical ambient temperatures, a tonic sympathetic outflow maintains these shunt vessels in a relatively closed state. This thermoregulatory mechanism is metabolically efficient, as it requires a minimal amount of energy and is therefore most frequently in use during normal conditions (see low threshold level for activation; Fig. 5). In infants, nonshivering thermogenesis follows as the next defense layer (elicited effector response) that is evoked to protect one from cold. Interestingly, thermogenesis via shivering is totally absent in the newborn and not fully effective until several years of age (Brooke et al., 1973). In contrast, in an adult, nonshivering thermogenesis has a relatively minor or marginal role (Jessen, 1980). In other words, skeletal muscle activation in the form of both involuntary tensing and shivering are of greater thermogenesis importance; these are the next effector responses to protect one from becoming hypothermic, that is, subsequent to initiated behavioral responses and vasoconstriction. By definition, shivering is involuntary, oscillatory muscular activity that can greatly increase one's metabolic heat production to counteract imminent hypothermia (Iampietro et al., 1960). It should be noted that within either the hypothermic individual or the hypothermic patient recovering from anesthesia, the elicitation of forceful shivering can increase metabolic heat production several fold. However, over a longer time period (hours), it is considered that only doubling the metabolic heat production can be maintained (Horvath et al., 1956; Giesbrecht et al., 1994). Moreover, one needs to consider that from the perspective of maintaining one's core temperature, the overall gain in core heat is less than expected; much of the heat produced by this increase in metabolic rate is generated in the large muscle groups of the extremities and thereby lost to the environment, instead of being retained to the core. Further, any increase in muscle activity leads to an increase in peripheral perfusion, thus counteracting the body's effort to minimize core-to-peripheral heat transfer. So, in general terms, one can consider that the shivering response is metabolically inefficient and can be viewed as a last resort response against impending hypothermia, that is, in cases wherein behavioral defenses and vasoconstriction are not available or have failed.
Core-to-periphery heat distribution in different environmental conditions.
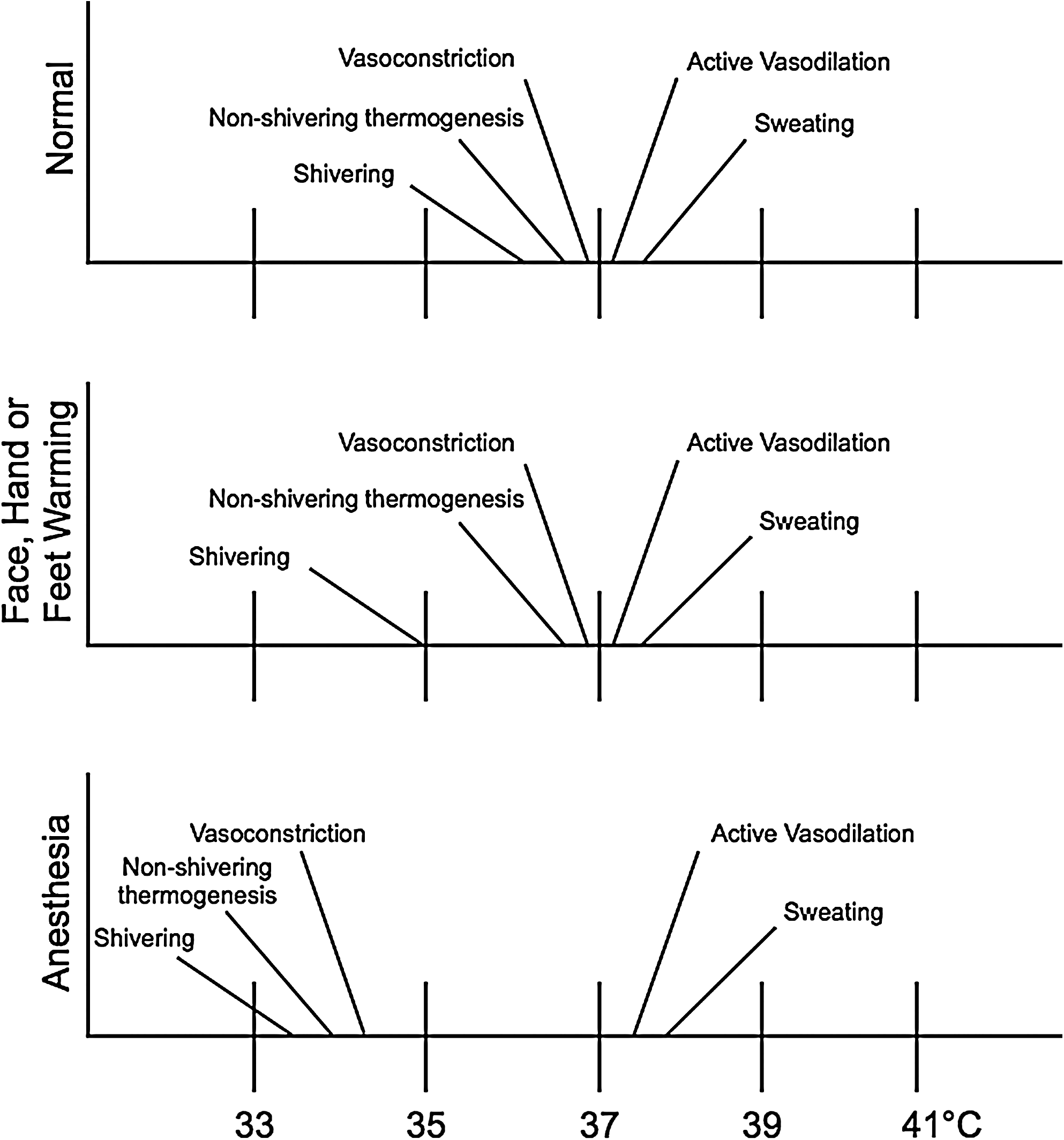
The autonomic thermoregulatory response thresholds in normal conditions, during warming of the face/hands/feet, or under general anesthesia.
Adverse Consequences of Thermoregulatory Defenses
During the active cooling of unanesthetized subjects, the autonomic thermoregulatory defense measures of the body can be vigorous. In such cases, the induced cutaneous vasoconstriction and involuntary motor activity (muscle tensing and shiver) will greatly reduce the rate at which an individual will become hypothermic or even completely avert any core temperature reductions. Although elderly subjects rarely shiver because advanced age impairs thermoregulatory responses (Frank et al., 1995; Vassilieff et al., 1995), the induction of therapeutic hypothermia can nevertheless often turn out to be difficult. Further, one has to be well aware of the detrimental effects associated with hypothermia relative to protective benefits (e.g., neural, cardiac, and other tissues). For example, cold-induced shivering increases the metabolic rate and thereby substantially increases one's cardiovascular work and oxygen consumption (Polderman and Herold, 2009). Frank et al. (1997a) observed an increased incidence of perioperative morbid cardiac events (i.e., unstable angina/ischemia, cardiac arrest, or myocardial infarction) and ventricular tachycardia in hypothermic patients (35.4°C±0.1°C), compared with normothermic patients (36.7°C±0.1°C). Earlier studies have suggested adverse effects of cold stress on the heart (Rose, 1966; Bainton et al., 1977; Lassvik and Areskog, 1979). However, Frank et al. (1997a) were able to show such a fatal interrelation in a prospective, randomized fashion. Interestingly, both in the aforementioned study and in an earlier study from the same group (Frank et al., 1993), shivering was not correlated to cardiovascular complications. This finding shifted the focus to the presence of a second basic thermoregulatory response in hypothermic patients, the adrenergic response with associated systemic vasoconstriction and hemodynamic changes. One can observe, during cooling, dramatic increases of heart rates as well as blood pressure levels (Polderman, 2009). Even at already mild hypothermia (0.5°C–1.2°C below normal core temperature), systemic vasoconstriction and increase in arterial blood pressure occurs (with up to a 700% increase in circulating norepinephrine levels) (Frank et al., 1995, 1997b). With these data, it seems prudent, even imperative in certain clinical situations (e.g., stroke patients with associated comorbidities), to dampen and minimize the adverse side effects of therapeutic hypothermia associated with the adrenergic (and shivering) response. Finally, the shivering reaction itself is unpleasant for conscious patients and should be avoided on the basis of discomfort alone. As will be noted later, these aforementioned side effects (with their undesirable consequences) have led to the common practice of strict temperature management throughout the hospital setting in the past two decades.
Modulation of Thermoregulation
Physical means
Invasive focalized cooling of the hypothalamus will immediately initiate profound central, whole-body shivering (“Frank” shiver) (Hammel et al., 1960). On the other hand, reducing skin temperature from 33°C to 30°C also initiates and maintains shivering, despite a brain temperature of 38°C (Lim, 1960). Consistent with these findings, humans placed in a 10°C environmental chamber for a period of 15–40 minutes experienced intense shiver despite the fact that their core temperatures stayed the same or even slightly increased (Iaizzo et al., 1983; Pozos and Iaizzo, 1991). It is interesting to note that applying radiant facial warming has been reported to greatly attenuate shiver, even when the original stimulus is intense and persistent (Mekjavic and Eiken, 1985). In further experiments from our group, both focal hand and facial warming were able to suppress shivering in mildly hypothermic patients (Fig. 6) (Iaizzo et al., 1999; Sweney et al., 2001). This warming facilitated and allowed the induction of mild hypothermia in healthy young individuals (this group represents the worst case scenario, as they shiver rigorously) without any pharmacologic intervention. Importantly, in these experiments, with the exception of focal warming (hands and face, respectively), the individuals were treated with cooling via conduction (water mattresses) and convective air (whole-body coverlets) (Fig. 7). Most of the skin surface was cooled, nevertheless shivering was avoided and suppressed efficiently, and core body temperature was decreased. This is also illustrated in Figure 8, wherein infrared thermographs show the time-wise changes of whole-body cutaneous temperatures of a subject during a noninvasive therapeutic cooling session. Before application of facial warming (t=25 minutes), shiver was evident with a concomitant increase in cutaneous temperature due to increased peripheral tissue perfusion. After initiation of facial warming, shivering ceased clinically and cutaneous body temperature became uniformly low. Thus, when applying this treatment to a stroke victim, for whom cooling of the head/brain is targeted, it is important to note that scalp temperatures remained cool even when facial warming was applied. Again to reiterate, the blunted or attenuated shivering response importantly facilitated the induction of mild hypothermia by external devices (cooling mattress and coverlet) without any pharmacologic intervention. It should be noted that both these normal subjects and most patients who suffer a stroke will have fully functioning thermoregulatory responses.
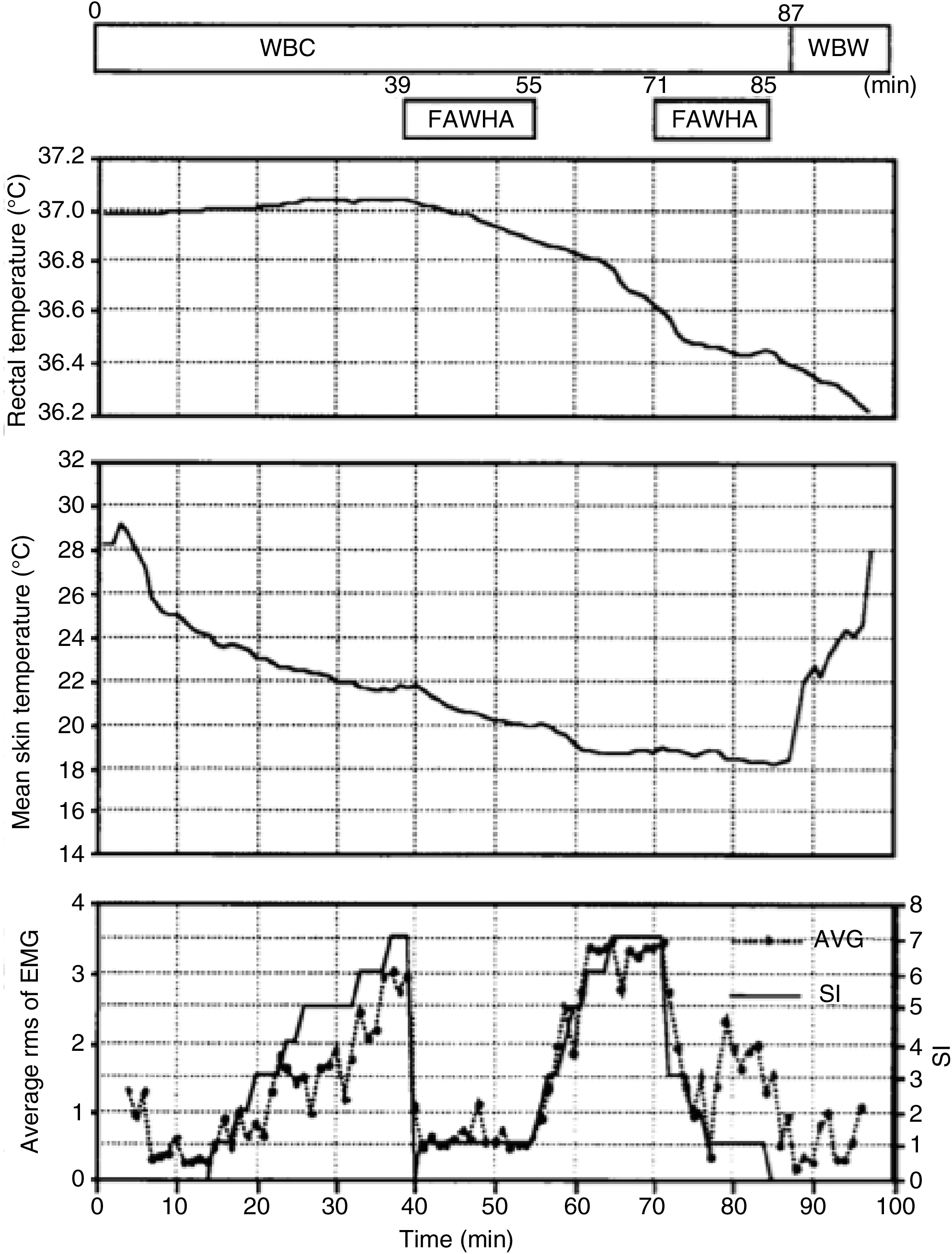
Temporal changes in rectal temperature, mean skin temperature, and involuntary muscular activity in a volunteer subjected to external whole-body cooling, facial application of warm air, and whole-body warming. The intensity of involuntary motor activity was assessed by measuring the relative changes in overall electromyogram activity and a subjective index of shiver (SI; analog scale from 0 to 10) (Iaizzo et al., 1999). AVG, average; rms, root mean squares; WBC, whole-body cooling; FAWHA, facial application of warm air; WBW, whole-body warming; EMG, electromyogram.
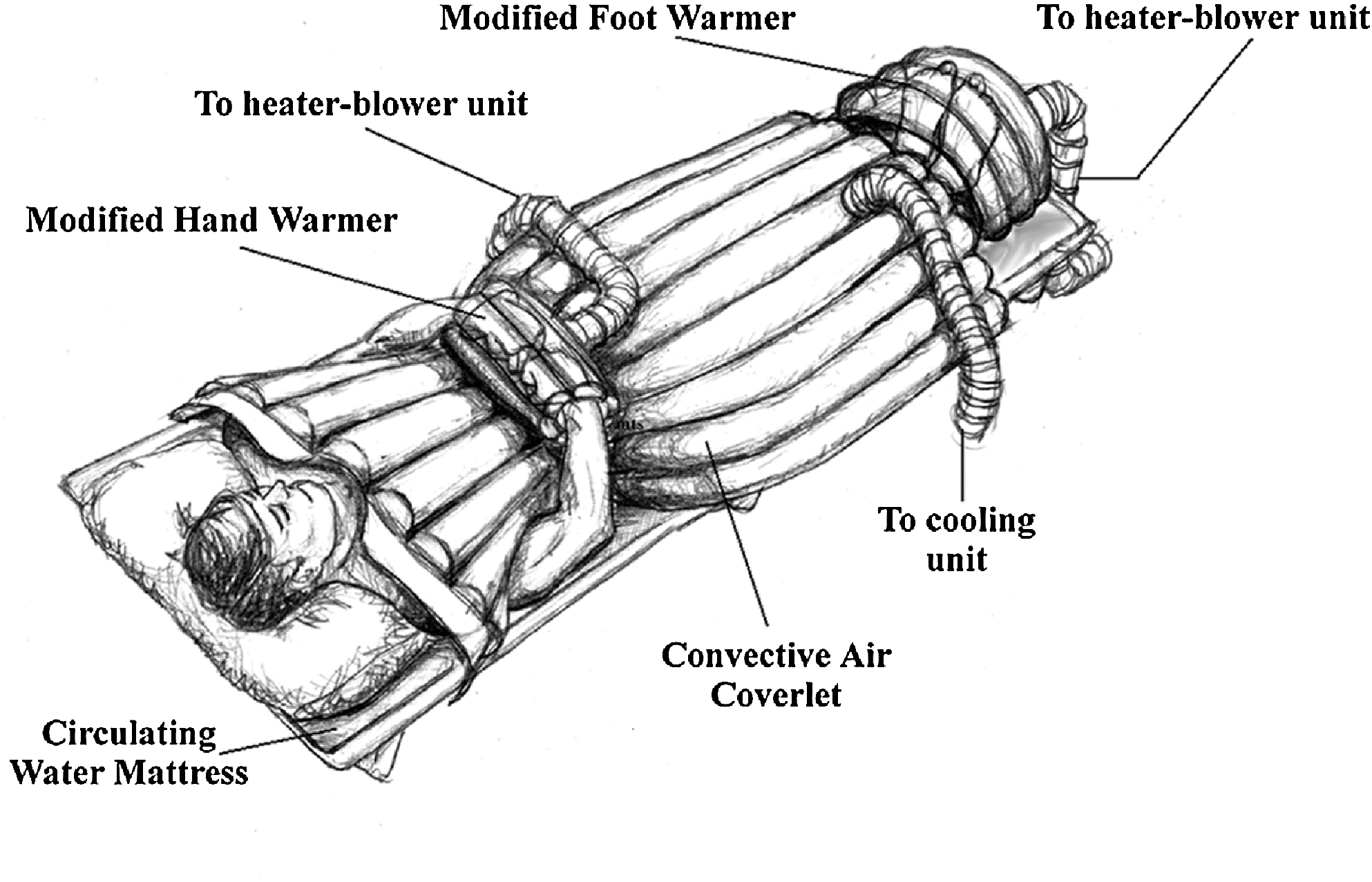
Hand and feet warming apparatus combined with external cooling [adapted illustration from Sweney et al. (2001).]
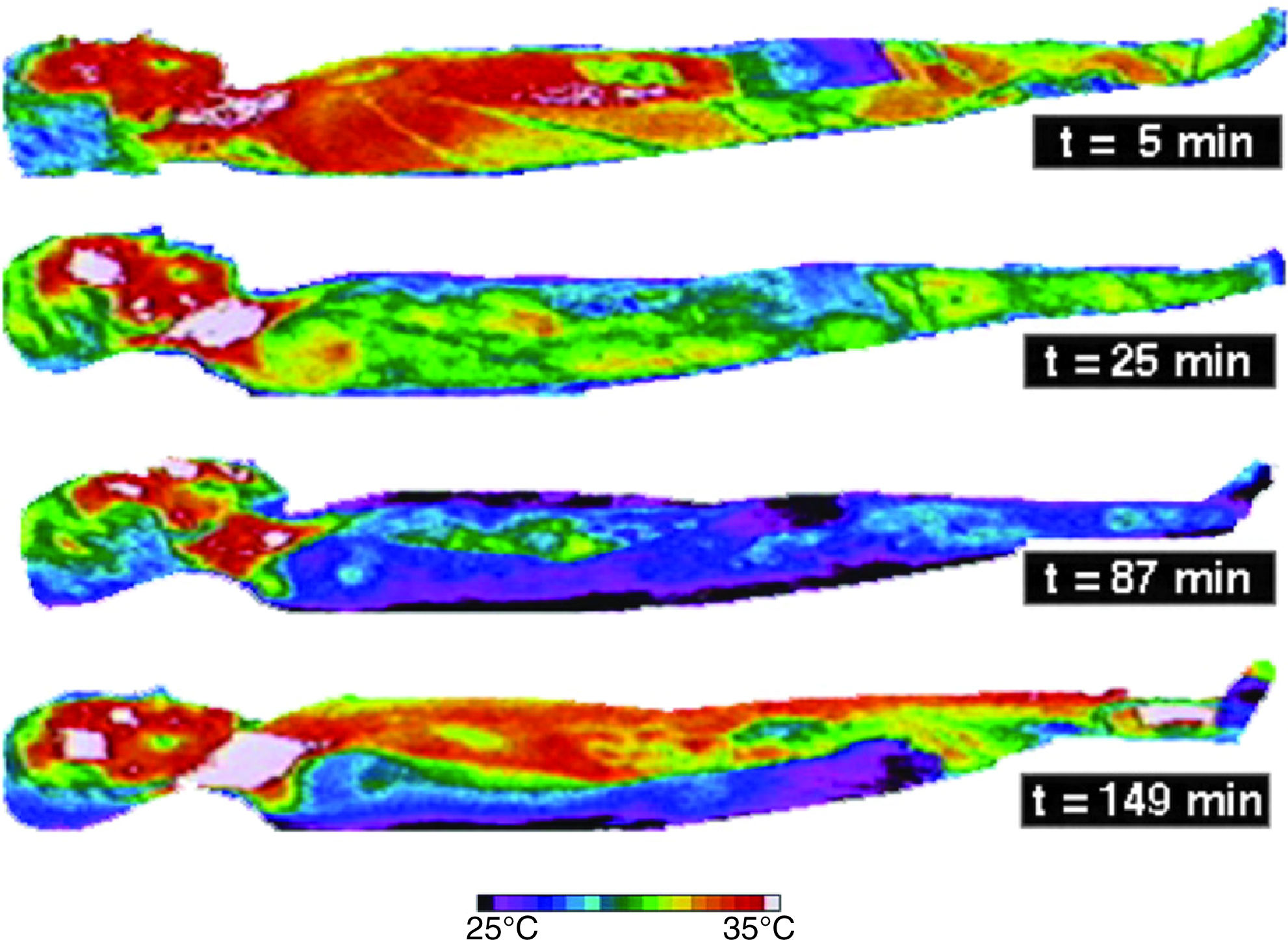
Infrared thermographs showing time-wise changes of whole-body cutaneous temperatures during external cooling. t=5 minutes, early stage prior to shiver; t=25 minutes, during cooling with evident shiver; t=87 minutes, after initiation of facial warming, at which time the shiver was suppressed; t=149 minutes, during whole-body rewarming with a convective air coverlet (Iaizzo et al., 1999).
As noted earlier, thermoreceptors have an unequal distribution throughout the body. Importantly, thermal information from the facial area, the hands, and feet are centrally overrepresented (Fig. 1). This integrated receptor input may help explain why focal facial or acral (hand and/or foot) warming is efficient in decreasing the whole-body shiver response. Concomitantly and perhaps equally important, all the subjects had significantly higher comfort scores during the facial or hand warming, compared with the periods when warming was turned off (Fig. 6). In an unpublished pilot study by our laboratory, we observed that the application of facial warming to patients who were mildly hypothermic in the postanesthetic recovery area was also associated with reduced rates of self-administered analgesic agents. Moreover, in a recent study by Badjatia et al. (2009), full-body surface counterwarming provided efficient control of shivering and was able to improve the metabolic profile of patients during therapeutic cooling, further underscoring the substantial potential and efficiency of these physical means in counteracting the thermoregulatory defenses of the body.
Pharmacological intervention
Anesthetic drugs
Humans are “homeotherms” capable of maintaining their body temperatures within a very narrow range. Yet, the administration of many general anesthetics alters this ability and transforms humans into “poikilotherms,” wherein core temperature will follow environmental temperature. General anesthetic agents have profound effects on the central nervous system and notably on central thermoregulation. Hence, under the influence of such agents, the interthreshold range increases by a factor of 10 or more during general anesthesia (Fig. 5); core temperature change is not counteracted by thermoregulatory defenses (Sessler, 2009a). More specifically, during such anesthesia, the sweating threshold increases only marginally, while the thresholds for both vasoconstriction and shivering are substantially decreased. This can be also observed at higher doses of anesthetics; in other words, larger shifts in interthreshold ranges occur at higher doses/potencies. For example, propofol (as well as opioids and sedatives) decreases both the vasoconstriction and shivering thresholds linearly (Sessler, 2009b). Yet, it has been reported that for volatile anesthetics the responses can be a bit more nonlinear, with an even larger, potentiated effect at higher dosages (Annadata et al., 1995; Kurz et al., 1995a; Matsukawa et al., 1995; Talke et al., 1997). The threshold levels for the two major cold defenses (vasoconstriction and shivering) are synchronously reduced, that is, the difference between the two threshold levels stays constant at 1°C. Further, most general anesthetic agents have a direct vasodilatatory action, hence increasing skin blood flow (with the exception of the vasoconstrictor ketamine) (Janssen et al., 1991). This, in turn, leads to direct core-to-periphery heat transfer and thereby rapid (within minutes) and significant drops in core body temperatures (Fig. 9). This altered heat distribution is a major cause and concern for perioperative hypothermia. At the same time, however, one can take advantage of this redistribution of heat by using anesthetics to induce therapeutic hypothermia more efficiently (Sessler, 2000). Indeed, Matsukawa et al. (1995) found that core-to-peripheral redistribution of heat accounted for 81% of the total decrease in core temperature after the first hour of anesthesia; this remained the dominant cause of hypothermia even in the following hours.
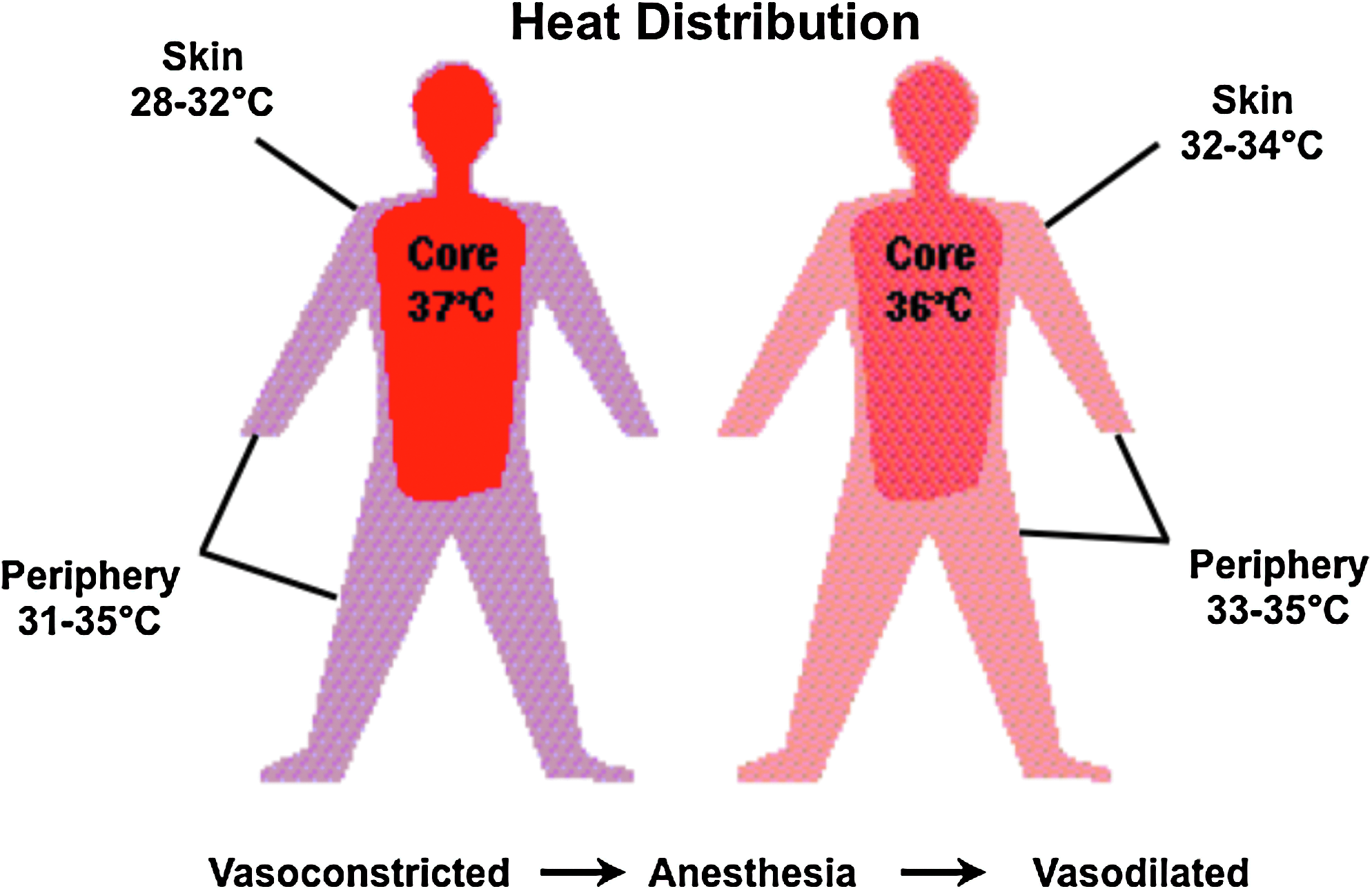
Core-to-periphery redistribution of body heat under general anesthesia.
Muscle relaxants
Muscle relaxants have no effect on the threshold levels of the various thermoregulatory defense mechanisms. Yet, muscle relaxants, through their peripheral action at the neuromuscular junction, will prevent all skeletal muscle tensing–shivering responses. In contrast, the associated central thermoregulatory changes and responses are not modulated, that is, the undesirable side effects of tachycardia and hypertension are not mitigated. It should also be noted that muscle relaxants can be only used in conjunction with anesthesia or sedation. Moreover, their prolonged use results in the functional denervation of skeletal muscles and has been linked to myopathy in severely ill patients. Therefore, their primary use today is limited and minimized within the intensive care setting (Hund, 1999). However, a recent systematic review on anesthesia and analgesia during hypothermia after cardiac arrest has shown that neuromuscular blocking drugs were routinely used to prevent shivering in 54 of 68 intensive care units (ICUs) and to treat shivering in 8 ICUs; note that the use of neuromuscular blocking agents was discouraged in only one ICU unit (Chamorro et al., 2010).
Opioids
Opioids are often administered to patients to minimize shiver as an effect response. For example, meperidine was frequently used for the treatment of perioperative shiver because of its beneficial and characteristic therapeutic profile (Burks et al., 1980). The administration of opioids inhibits the motor (shiver) thermal response at twice the rate as the vasoconstrictive response, at any given dose (Leslie et al., 2004). The antishivering action of meperidine is already present at blood levels that do not lead to any observable respiratory depression or sedation, for example, a relatively small dose (25 mg, iv) of meperidine decreases the shivering threshold by 2°C (Alfonsi et al., 1998). Alfentanyl, a pure μ-receptor agonist, is another therapeutic option for shiver control. This agent provides for a dose-dependent reduction in shivering threshold (Kurz et al., 1995a); however, it takes relatively high drug doses to induce substantial effects. Thus, because of the risk of respiratory depression at high doses, the sole use of alfentanyl for shiver control is commonly discouraged. Chamorro et al. (2010) have shown, in their recent systematic review on comatose cardiac arrest patients treated with therapeutic hypothermia, that fentanyl was the most used analgesic agent (at doses between 0.5 and 10 μg/kg/h), followed by morphine. However, it should be noted that in their treatment algorithm for employing therapeutic hypothermia, these authors propose the use of remifentanil, mainly because of its pharmacokinetic considerations.
Sedatives (nonopioids)
Sedatives (nonopioids) have been shown to have some effect on vasoconstriction and shivering thresholds; for example, midazolam, a benzodiazepine, is commonly used in this manner (as a sedative agent in 57% of the ICUs in a recent systematic review on patients treated with therapeutic hypothermia, with doses between 5 mg/h and 0.3 mg/kg/h) (Chamorro et al., 2010). However, even with the use of high doses of such agents for sedation, thermoregulatory thresholds are only reduced by approximately 0.6°C (Kurz et al., 1995b). Further, because of depressed CYP3A4 and CYP3A5 activities during hypothermia, the sedative effects of midazolam may then be unpredictably prolonged in these circumstances (Tortorici et al., 2007). It should be noted that clonidine, a central α-2 receptor agonist, only slightly decreases vasoconstriction and shivering thresholds as well, despite the administration of substantial dosing (Delaunay et al., 1991). Yet, a slightly stronger effect on the thermoregulatory defense mechanism is seen with the use of a related drug, dexmedetomidine. Nevertheless, the overall degree of thermoregulatory inhibition in relation to the administration of sedation is small. In other words, the use of dexmedetomidine as a single agent for the induction of therapeutic cooling is not effective or practical. Further, buspirone, a serotonin 1A partial agonist, was shown to reduce the shivering threshold to 35.0°C±0.8°C (Mokhtarani et al., 2001). However, this threshold reduction is only minor clinically; thus, buspirone seems not sufficiently potent for use as a single agent to alter thermoregulatory responses.
Drug combinations
Today, we have not identified the use of a single pharmacologic agent that is sufficient to induce a substantial tolerance toward therapeutic hypothermia; hence, the approach of combining different agents has been studied. This combination approach has the important advantage of reducing the known side effects (sedation, bradycardia, and respiratory depression) by utilizing each agent at lower doses. For example, the combination of meperidine and dexmedetomidine has been studied for such applications; this combination therapy was shown to have an additive effect, decreasing the shivering threshold level to about 35°C (Doufas et al., 2003). Another example of additive beneficial effects is the combination of buspirone and dexmedetomidine. The combined administration of these medications reduced the shivering threshold level to about 34°C (Lenhardt et al., 2009). Additionally, a synergistic therapeutic effect was demonstrated for the combined use of buspirone and meperidine; administration at low doses reduced the shivering threshold to 33.4°C±0.7°C. Importantly, because of low drug concentrations, there was no sedation or respiratory depression associated with this treatment approach (Mokhtarani et al., 2001).
Mild Hypothermia and Cerebral Protection
Significant protection against cerebral ischemia may occur by core body temperature reduction of as little as 1°C (Busto et al., 1987, 1989). Further, in a prospective study it was shown that, in acute stroke, an association existed between body temperature and initial stroke severity, infarct size, mortality, and outcome; results were best in those patients who were mildly hypothermic at the time they arrived for treatment (Jorgensen et al., 1996; Reith et al., 1996). Consistent with this report, it was shown that in patients with acute stroke, the occurrence of fever during the first 7 days was associated with a higher risk of death in the first 10 days; it was concluded that patients with a higher temperature had a worse stroke outcome (Azzimondi et al., 1995). Recently, Kollmar et al. (2010) observed that hypothermia (35°C) prevented the increase of perihemorrhagic edema in patients with large spontaneous intracerebral hemorrhage.
Hypothermia-induced cerebral protection was initially considered to be mainly due to reduced metabolism (Michenfelder and Theye, 1970; Steen et al., 1979), that is, the body and brain can tolerate prolonged periods of ischemia simply because of reduced demands for oxygen at these lower temperatures. This theory, however, has been challenged by studies that have found cerebral protective effects at only modest hypothermia (i.e., reductions in brain temperature of 2°C–5°C) (Busto et al., 1987; Natale and D'Alecy, 1989; Leonov et al., 1990; Minamisawa et al., 1990a, 1990b). Using established animal models of complete (Natale and D'Alecy, 1989; Leonov et al., 1990) and near-complete (Busto et al., 1987; Minamisawa et al., 1990a, 1990b) ischemia, these authors observed improved postischemic neurologic function (Natale and D'Alecy, 1989; Leonov et al., 1990) and histopathology (Busto et al., 1987; Leonov et al., 1990; Minamisawa et al., 1990a, 1990b) in the hypothermia-treated animals. Importantly, in some of these studies, improved recovery from ischemia was even observed when hypothermia was instituted during or immediately following the ischemic insult (Busto et al., 1987; Natale and D'Alecy, 1989; Leonov et al., 1990; Minamisawa et al., 1990a). Further, in two of the studies, no reduction of lactate production during ischemia was observed in hypothermia-treated subjects (Busto et al., 1987; Natale and D'Alecy, 1989). These findings indicated that reduction of global cerebral metabolism was not the sole mechanism involved in the beneficial effects of mild hypothermia. Indeed, a myriad of additional mechanisms of (mild) hypothermia have been unveiled since (Lanier et al., 1992), positively mitigating and modulating destructive processes associated with ischemia-reperfusion, for example, excitotoxicity, neuroinflammation, blood–brain barrier disruption, free radical production, apoptosis, and numerous others (for a detailed review on the mechanisms of action of hypothermia, see Polderman, 2009).
To date, there exists strong evidence of important cerebral protection in two clinical scenarios, first, in the case of cardiac arrest victims with ventricular fibrillation as the initial presenting cardiac rhythm (Bernard et al., 2002; Hypothermia after Cardiac Arrest Study Group, 2002) and, second, in the case of asphyctic newborns (Shankaran et al., 2005; Azzopardi et al., 2009), wherein hypothermic treatment proved efficient. Nevertheless, there is much ongoing research on new therapeutic indications for mild hypothermia, including that in conscious patients. If such approaches prove to be clinically beneficial, the outlined approaches described here could also prove helpful in delivering optimal patient care.
Conclusion
Hypothermia, either profound or mild, has been repeatedly shown to be a potent therapeutic option. Although the induction of therapeutic hypothermia in anesthetized, unconscious patients is rather straightforward, this is not the case in conscious, unanesthetized patients. The thermoregulatory defenses (effector responses) in the awake patient population can be vigorous and difficult to overcome. However, there are means to circumvent these thermoregulatory defenses (physiological tricks), and several antishivering protocols have shown good clinical potential. More specifically, if one were to combine such physical means together with a low-dose combination of antishivering drugs, this could be seen as very promising for an efficient clinical induction of therapeutic hypothermia in those patients in whom cerebral protection is required.
Footnotes
Acknowledgments
We would like to thank Monica Mahre for her assistance with manuscript preparation and both Kiley Schmidt and Dr. Matt Sweney for providing illustrations.
Disclosure Statement
The authors declare that no competing financial interests exist.