
Abstract
Therapeutic hypothermia (TH) mitigates damage in ischemic stroke models. However, safer and easier TH methods (e.g., pharmacological) are needed to circumvent physical cooling complications. This study evaluated systemic and pharmacologically induced TH using the adenosine A1 receptor agonist, N6-cyclohexyladenosine (CHA), with control groups in male Sprague–Dawley rats. CHA was administered intraperitoneally 10 minutes following a 2-hour intraluminal middle cerebral artery occlusion. We used a 1.5 mg/kg induction dose, followed by three 1.0 mg/kg doses every 6 hours for a total of 4 doses, causing 20–24 hours of hypothermia. Animals assigned to physical hypothermia and CHA-hypothermia had similar induction rates and nadir temperatures, but forced cooling lasted ∼6 hours longer compared with CHA-treated animals. The divergence is likely attributable to individual differences in CHA metabolism, which led to varied durations at nadir, whereas physical hypothermia was better regulated. Physical hypothermia significantly reduced infarction (primary endpoint) on day 7 (mean reduction of 36.8 mm3 or 39% reduction; p = 0.021 vs. normothermic animals; Cohen's d = 0.75), whereas CHA-induced hypothermia did not (p = 0.33). Similarly, physical cooling improved neurological function (physical hypothermia median = 0, physical normothermia median = 2; p = 0.008) and CHA-induced cooling did not (p > 0.99). Our findings demonstrate that forced cooling was neuroprotective compared with controls, but prolonged CHA-induced cooling was not neuroprotective.
Introduction
Therapeutic hypothermia (TH) is a widely studied treatment for ischemic brain injuries, with demonstrated efficacy in cardiac arrest and neonatal hypoxic–ischemic encephalopathy (Lemyre and Chau, 2018; Myat et al., 2018). Regarding focal ischemic stroke, preclinical meta-analyses have shown improved outcomes in TH-treated animals compared with controls (Dumitrascu et al., 2016). On average, meta-analyses show ∼40% to 45% lesion volume reductions and ∼30% to 45% improved neurobehavioral scores in animals treated with TH.
Despite relatively extensive research, clinical translation efforts have faced challenges related to both the safety and efficacy of TH in ischemic stroke patients: TH induction methods are time-consuming (Clifton et al., 2001; Schwab et al., 1998), and TH has undesirable side effects (e.g., shivering, pneumonia; Geurts et al., 2014; Kuczynski et al., 2020).
To overcome some of the aforementioned problems, research has identified alternative protocols (e.g., local or pharmacological TH methods) and cotherapies (e.g., antishivering protocols). Novel pharmacological TH methods have been employed with hopes of producing TH more easily (Sun et al., 2019). Ideally, pharmacological agents should reduce body temperature without triggering reactive thermogenic responses (Jain et al., 2018).
Pharmacological hypothermia may be advantageous for its simplicity compared with most physical cooling methods. However, thorough testing at preclinical and clinical levels is necessary to understand the safety, efficacy, and side effects of such approaches.
Increased extracellular adenosine levels occur during and following ischemia, which is part of an endogenous neuroprotective response (Melani et al., 2014). N6-cyclohexyladenosine (CHA) is an A1 adenosine receptor (A1AR) agonist that reduces thermogenesis and induces a hibernation-like state in animals (Jinka et al., 2015). A1ARs are located on neurons and glial cells in the cortex, hippocampus, and cerebellum (Sheth et al., 2014).
CHA exerts torpor-like hypothermia through agonism of A1AR in hypothalamic preoptic area nuclei (Hrvatin et al., 2020). Additionally, A1ARs couple to inhibitory G-proteins, reduce glutamate release, and decrease neuronal excitability (Rivera-Oliver and Díaz-Ríos, 2014; Stone et al., 2009).
CHA potentially can have “dual” neuroprotective effects: through hypothermia and by directly reduced neuronal excitability. Outside of the hypothalamus, activation of A1ARs reduces the excitability of cortical neurons independently of temperature (e.g., Rudolphi et al., 1992; Williams-Karnesky and Stenzel-Poore, 2009). As far as we are aware, there are no published reports on whether CHA lessens injury after focal cerebral ischemia.
However, similar studies have been conducted with other adenosine A1 and A3 receptor agonists such as cyclopentyladenosine and N6-R-phenylisopropyladenosine, where they often, but not always (Heron et al., 1994; Roussel et al., 1991), demonstrate protective effects in focal stroke models (Rudolphi et al., 1992). More recently, a study showed that the adenosine A1 and A3 receptor agonist, AST-004, reduces brain injury without reducing core temperature in a nonhuman primate model of middle cerebral artery occlusion (MCAO) (Liston et al., 2022).
Global ischemia studies have shown that CHA reduces CA1 pyramidal cell loss, ameliorates neurological deficits, improves survival, and conversely that acute adenosine A1 receptor antagonism worsens injury (Daval et al., 1989; von Lubitz et al., 1988; Zhou et al., 1994). Surprisingly, many in vivo studies did not control or measure temperature following stroke and CHA administration, making it impossible to discern the extent to which neuroprotective effects were actually mediated by hypothermia.
Ultimately, the ability of CHA to induce rapid hypothermia makes it a desirable candidate as a pharmacological TH agent (e.g., reducing temperature by 1 to 1.75°C/h; Jinka et al., 2015). Given the relatively strong foundation of research demonstrating the efficacy of adenosine receptor agonists in ischemic stroke and the efficacy of CHA following global cerebral ischemia, we hypothesized that CHA administration would lessen injury following focal ischemic stroke.
This study aimed to determine whether CHA was neuroprotective in a rat model of focal ischemic stroke, and as a positive control, we used a systemic TH protocol with proven neuroprotective efficacy in stroke models (Clark et al., 2008). Previous studies using this physical TH protocol have demonstrated up to 50% infarct volume reductions in permanent MCAO models (Yanamoto et al., 2001).
To investigate the role of the temperature-dependent and independent effects of CHA, we constructed a 2 × 2 factorial design that included hypothermic (HYPO) or normothermic (NORMO) animals, with either physical or pharmacological methods (HYPO or NORMO × physical or CHA), following endovascular suture occlusion of the middle cerebral artery. We selected this stroke model because it offers heterogeneity in the degree of injury that is consistent with the Stroke Treatment Academic Industry Roundtable criteria forming the gold standard of translational stroke research (Fisher et al., 2009; Kent and Mandava, 2016).
Given that this is the first use of CHA in the context of focal ischemic stroke, we did not use female animals to maximize internal control and focus on whether potential benefits were first seen in male animals (Dirnagl, 2019), which simplified the design of our study and limited the workload. Moreover, to detect potential sex effects (assumed to be smaller than treatment effects), a far larger sample size would be required to have sufficient statistical power.
Notably, this injury profile is representative of a population of patients who may not all benefit from clinical reperfusion therapies alone and thus are more likely to require supplemental therapies (Campbell et al., 2019; Jickling and Sharp, 2015). Our primary endpoint was infarct volume, and our secondary endpoint was behavioral deficits measured using a neurological deficit scale. We hypothesized that any form of body-wide TH would reduce brain injury, thereby improving behavioral outcomes compared with NORMO controls.
Methods
Ethics statement
Our procedures were developed a priori (but not preregistered) and later approved by the Animal Care and Use Committee of the University of Alberta, and comply with the Canadian Council on Animal Care Guidelines. Our experiments were designed with animal ethics in mind and reported according to the ARRIVE guidelines (Kilkenny et al., 2010).
Group sizes were determined a priori using G-Power (version 3.1). Using previous data from the surgeon performing the MCAO surgery (assuming a standard deviation of ∼30 mm3), we needed at least 17 animals per group to detect a 25–30% reduction (Clark et al., 2008) in infarct volume with 80% statistical power (α = 0.05; two-tailed test).
With the exception of 6 pilot animals, rats were randomly allocated using an online tool at Random Lists website. The surgeon was blinded to the group allocation during the induction of MCAO, and histological analyses were performed in a blinded manner. A timeline, including the key points of the experimental design for this study, is provided in Figure 1A.
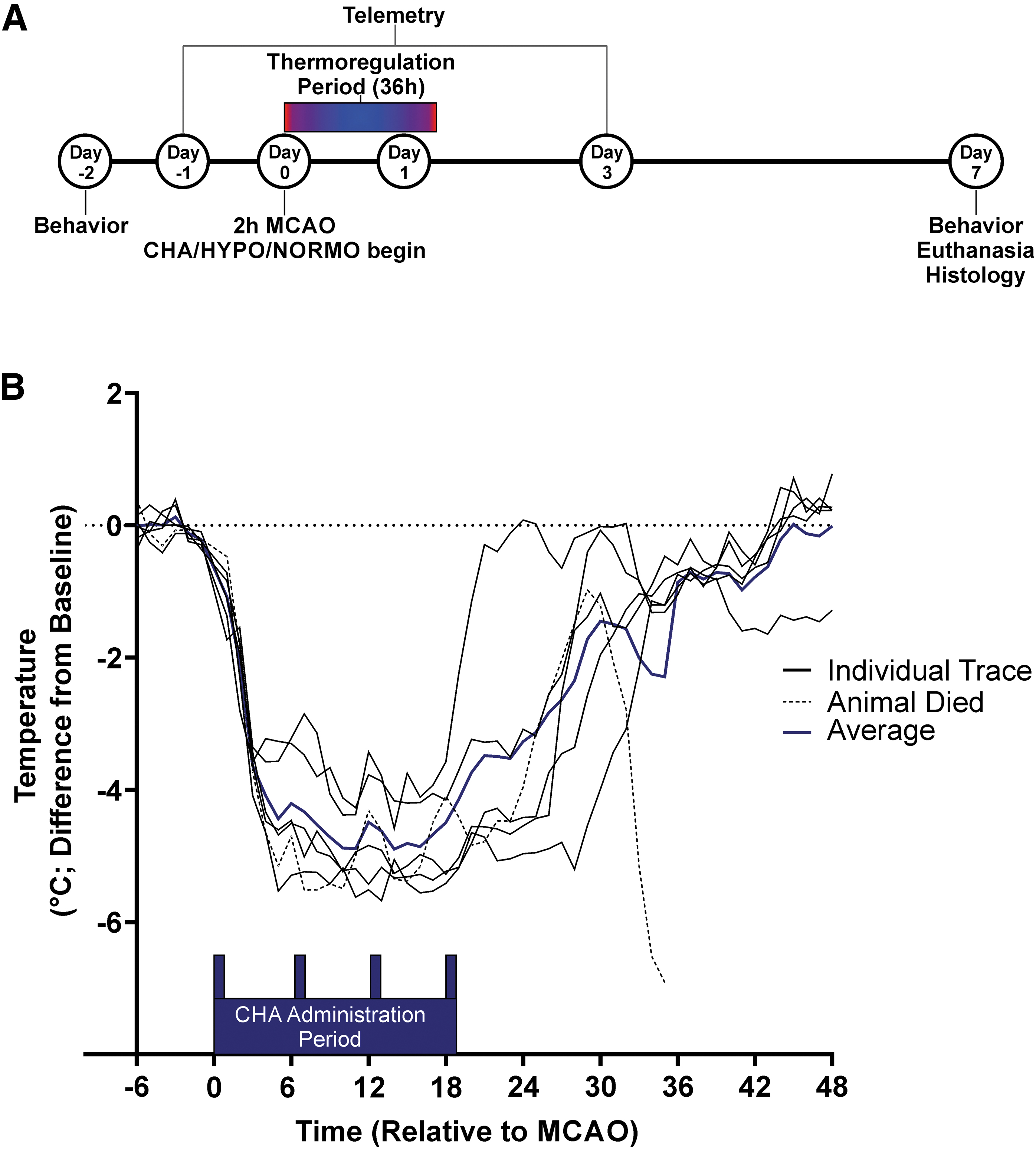
Experimental design and pilot study core temperature data.
Subjects
Ninety-seven male Sprague–Dawley rats (weight 320 ± 30 g; age: 9–11 weeks) were obtained from Charles River Laboratories (Saint-Constant, Quebec, Canada). Animals were housed individually in a temperature- and light-controlled room with 12-hour light–12-hour dark cycles for the duration of the experiment to collect telemetry data. Animals were given food (Purina Rodent Chow) and water ad libitum. Following MCAO, all rats were given wet mashed rodent chow, peanut butter, and shelled sunflower seeds to mitigate weight loss. Animals were checked postoperatively every 6 hours for at least 3 days.
Surgical procedures
All surgical procedures were performed aseptically during the light phase of the cycle. Surgical regions were shaved, washed with ethanol, and coated with Betadine before being draped before incision. During all procedures, rats were anesthetized with 4% isoflurane and maintained at 2% (60% N2O, balance O2). Body temperature was controlled and recorded using a rectal temperature probe, and body temperature was maintained near 37°C.
Postoperative analgesia was achieved following all procedures using locally applied Bupivacaine (0.5 mg/kg subcutaneous [S.C.]; SteriMax, Inc., Oakville, Ontario). Intraoperative pulse oximetry variables were collected using a paw sensor (PhysioSuite; Kent Scientific, Torrington, CT).
Core probe implant
Sterilized and calibrated telemetry probes (Model TA10TA-F40; Data Sciences International, St. Paul, MN) were implanted into the abdominal cavity as before (Liddle et al., 2020). Probes were implanted at least 3 days before MCAO, and baseline data were collected at least 24 hours before MCAO. Data were collected every 30 seconds and averaged every hour. The TA10TA-F40 probes monitor body temperature and relative whole-body movement across the radiofrequency receiver.
Postoperative analgesia was achieved with Bupivacaine (0.5 mg/kg S.C.) and Meloxicam (0.2 mg S.C.; Boehringer Ingelheim Ltd.).
Endovascular suture occlusion model
Following the method of Longa et al. (1989), the neck was shaved and prepared with Betadine. Next, a midline incision was made, and the common, internal, and external carotid arteries were isolated. Following isolation, a silicone rubber-coated 4-0 suture (30 mm length, 2–3 mm coating length; Doccol Corporation, Sharon, MA) was advanced 20–22 mm into the internal carotid artery, through the external carotid artery.
The occluding suture was fastened using a 5-0 silk suture, and the animals were awakened from the surgery. About 115 minutes following MCAO, rats were briefly reanesthetized (<10-minute duration of anesthesia) and the occluding suture was removed 120 minutes post-MCAO, establishing reperfusion. The external carotid artery stump was tied off, and wound clips closed the neck incision.
CHA administration
We dissolved CHA in 8% dimethyl sulfoxide (DMSO; Sigma-Aldrich) in physiological saline. To induce a torpor-like state, we first administered an induction dose of CHA 10 minutes following reperfusion (1.5 mg/kg intraperitoneal [I.P.]). We maintained the torpor-like state for ∼24 hours by giving 3 additional maintenance doses of CHA every 6 hours (1.0 mg/kg I.P.). We selected these doses of CHA based upon previous literature and early dose–response experiments (data not shown) that showed inconsistent hypothermia with lower induction doses (thus indicating a threshold dose needed to achieve torpor; Jinka et al., 2015).
On average, each rat received 0.11 mL (∼0.25 mg/kg) of DMSO and 1.4 mg of CHA across the 4 doses.
Thermoregulation protocols
Cyclohexyladenosine-hypothermia
Because CHA-related hypothermia reduces the body temperature of the animal to ambient (room) temperature, we set a lower limit of 33°C in the CHA-HYPO rats to prevent complications related to deep hypothermia and to maintain TH consistent with current clinical practices (Wan et al., 2014). Servo-controlled, 175-W, infrared heating lamps (∼30 cm above the animal) were used based upon core temperature data for 36 hours post-MCAO (Colbourne et al., 1996).
Hypothermia
We modeled our physical TH protocol from a pilot study of 6 CHA-treated rats allowed to cool to 33°C following the induction and maintenance doses. From this, we identified the physical TH parameters that we applied to our HYPO animals: onset time of 10 minutes post-MCAO, duration of ∼24 hours, induction rate of −0.8°C/h, TH depth of ∼33°C, and rewarming rate of 0.36°C/h. Physical TH was achieved using a servoregulation system that modulates temperature with application of water mist, coupled with an overhead fan, and with the use of overhead, 175-W, infrared heating lamps to avoid getting too cold and to aid in rewarming (Colbourne et al., 1996).
The automated system maintains the temperature within 0.5°C of the target temperature, and our TH protocol was similar to previously published clinical trials (Shankaran et al., 2012; Tahir and Pabaney, 2016). As well, this protocol is similar to our past work demonstrating enduring neuroprotection against global (Colbourne et al., 1999) and focal ischemia (Clark et al., 2009). Thus, this physical cooling protocol was expected to serve as a positive control and also to be a reasonable treatment from which to base the CHA-induced cooling upon.
CHA-NORMO and NORMO treatments
Both CHA-NORMO and NORMO animals were prevented from having body temperatures <36.5°C through the use of infrared heat lamps. We chose to maintain animals at this temperature (∼0.5°C to 1°C lower than baseline) rather than the true NORMO body temperature for rats to prevent unnecessary stress and any potential risk of overheating in these control groups.
We and others have shown that brain temperature corresponds to body temperature in freely moving nonanesthetized rats, although it is slightly lower (within ∼1°C; Chen et al., 1992; DeBow and Colbourne, 2003), and a study showed a similar correspondence during CHA-induced cooling (Laughlin et al., 2018). This very mild level of cooling might also be neuroprotective based upon past studies (e.g., 34°C, an ∼2°C drop from normal values; Colbourne and Corbett, 1995).
Behavioral assessment: neurological deficit score
All animals underwent two 10-minute handling sessions as well as a familiarization session to minimize confounding by stress. Behavioral deficits were assessed 2 days before MCAO (baseline) and on day 7 post-MCAO. The neurological deficit score (NDS) was scored as previously described (MacLellan et al., 2006). Briefly, each rat received a score from 0 (no deficits) to 14 (severe deficits).
Rats were scored on 5 subtests. These included spontaneous circling (graded from 0: no circling, to 3: continuous circling), contralateral hindlimb retraction (0: immediate retraction when displaced, 3: no retraction), bilateral forepaw grasp (0: normal grasp of a bar, 3: rat unable to grasp bar), beam walking ability (0: readily crosses a beam, 3: rat unable to cross or stay on a beam for more than 10 seconds), and contralateral forelimb flexion (0: uniform forelimb extension, 2: shoulder adduction and wrist flexion).
Infarct volume quantification
On day 7 post-MCAO, rats were overdosed with sodium pentobarbital (100 mg/kg I.P.; Bimeda-MTC, Lavaltrie, Quebec) and transcardially perfused with 0.9% saline and 10% neutral buffered formalin. Tissue was sectioned at 100 μm, thionin stained, and analyzed using ImageJ (Version 1.51; NIH).
We quantified infarction volume as follows: tissue loss = volume of healthy tissue in the unaffected hemisphere – volume of healthy tissue in the affected hemisphere (MacLellan et al., 2006); tissue loss in the unaffected hemisphere = volume of the uninjured hemisphere – ventricular volume; and tissue loss in the affected hemisphere = volume of the injured hemisphere – (ventricular volume + infarct volumes). This method accounts for atrophy and ventriculomegaly from injury.
Statistical analysis
Data were analyzed and visualized using GraphPad Prism 8 (GraphPad Software Inc.; San Diego, CA). Assumptions of analysis of variance (ANOVA) were checked visually and formally analyzed using Levene's test and Shapiro–Wilk test. When assumptions were met, data were analyzed using parametric techniques. Ordinal data were analyzed using nonparametric techniques (e.g., Mann–Whitney U test).
Data are expressed either as means ±95% confidence intervals (95% CIs) or medians with interquartile ranges. An α-value threshold at 0.05 represented statistical significance.
Results
Intraoperative variables
Intraoperative temperature and pulse oximetry data are tabulated in Table 1. There were no significant differences in intraoperative temperature or oxygen saturation. However, small (<10 beats per minute) differences in heart rate were detected by one-way ANOVA: CHA-HYPO animals had significantly faster heart rates than CHA-NORMO (p = 0.024) and HYPO rats (p = 0.017), and NORMO rats had significantly faster heart rates than CHA-NORMO (p = 0.0001) and HYPO rats (p < 0.0001).
Intraoperative Physiological Variables
Intraoperative temperature and O2 saturation did not differ between groups.
Significantly different from CHA-HYPO and NORMO.
BPM, beats per minute; CHA, cyclohexyladenosine; HYPO, hypothermic; NORMO, normothermic; SD, standard deviation.
Pilot experiment
In our pilot CHA-HYPO animals, one animal died apparently as a result of subarachnoid hemorrhage, leaving us with five animals.
Temperature and activity data were collected from the pilot CHA-HYPO rats following MCAO (Fig. 1B). These temperature data were used to develop the physical TH protocol.
Mortality and exclusions
We determined a priori that animals would be excluded if they did not reach their experimental endpoint (e.g., due to mortality or premature euthanasia) or if there were intraoperative or treatment-related complications (e.g., incomplete treatment due to computer power failure). A total of 22 animals were excluded in our study: 17 animals died (2 of these 17 died before treatment, 11 had evidence of subarachnoid hemorrhage, and 4 died without evidence of hemorrhage), 2 were prematurely euthanized (excessive body weight loss and failure to thrive), 2 were excluded due to surgical error, and 1 was excluded because of technical difficulties.
Of the animals that died, 6 were in the CHA-HYPO group, 6 were in the CHA-NORMO group, 1 was in the HYPO group, and 4 were in the NORMO group. Mortality differences among groups were not significant following a chi-squared test (p = 0.18). After exclusions, the group sizes were as follows: 16 in the CHA-HYPO group, 15 in the CHA-NORMO group, 19 in the HYPO group, and 19 in the NORMO group.
Poststroke weight changes
To evaluate potential CHA-related side effects, we assessed postoperative recovery of body weight at day 7 post-MCAO. Rats had similar body weights on day 7 post-MCAO, demonstrating similar recovery of weight loss (one-way ANOVA p = 0.65; data not shown).
Postoperative temperature
Two-way ANOVA of postoperative temperature data revealed that time, treatment, and interaction factors were significant (p < 0.0001 for all; Fig. 2A). Thus, simple effects analyses were conducted to further explore temperature differences between select groups. When we compared CHA-HYPO treatment with physical hypothermia, CHA-treated animals were significantly cooler at 2 and 3 hours post-MCAO, but between 4 and 24 hours, there were no differences between groups.

Core temperature and relative activity data collected using abdominal telemetry probes. Lines represent means for each group.
During the rewarming period, CHA animals were warmer than HYPO animals until animals returned to baseline (p ≤ 0.03 for all comparisons 25–40 hours post-MCAO; range of differences = 0.9–2.9°C). CHA-NORMO rats were significantly colder than NORMO animals until 35 hours post-MCAO, thus CHA-NORMO animals were mildly HYPO (∼1°C) for ∼1.5 days post-MCAO.
Postoperative activity
Two-way ANOVA of postoperative activity showed that time, treatment, and interaction factors were significant (p < 0.001 for all; Fig. 2B). In the first 24 hours, simple effects analyses showed that HYPO animals had significantly less activity than NORMO animals at 6 hours until 12 hours, then again at 14, 18, 20, 21, 23, and 24 hours post-MCAO. Between 24 and 48 hours, HYPO rats were significantly less active at 29–33 hours, then again at 35 and 36 hours post-MCAO.
When we compared the activity of CHA-HYPO rats and NORMO rats, CHA-HYPO rats had significantly less activity (p ≤ 0.05) at 6–10, 12, 14–16, 19–23, 28, 30–33, and 35–36 hours post-MCAO. There were no significant differences in activity between CHA-HYPO and CHA-NORMO rats in the first 48 hours post-MCAO. In general, both CHA treatments and HYPO treatment appeared to reduce animal activity, compared with NORMO animals.
HYPO, but not CHA-HYPO, animals showed reduced histological tissue loss
A planned comparison between HYPO and NORMO groups demonstrated that HYPO animals had a substantial 39% reduction in tissue loss (mean difference = 36.76 mm3; Cohen's d = 0.75 (95% CI of Cohen's d: 0.07–1.43); Fig. 3A) compared with NORMO controls (p = 0.026). Planned contrasts between CHA-HYPO and CHA-NORMO animals did not demonstrate a significant difference (p = 0.33), nor did an unplanned comparison between CHA-HYPO and NORMO animals yield a significant difference (p = 0.82).
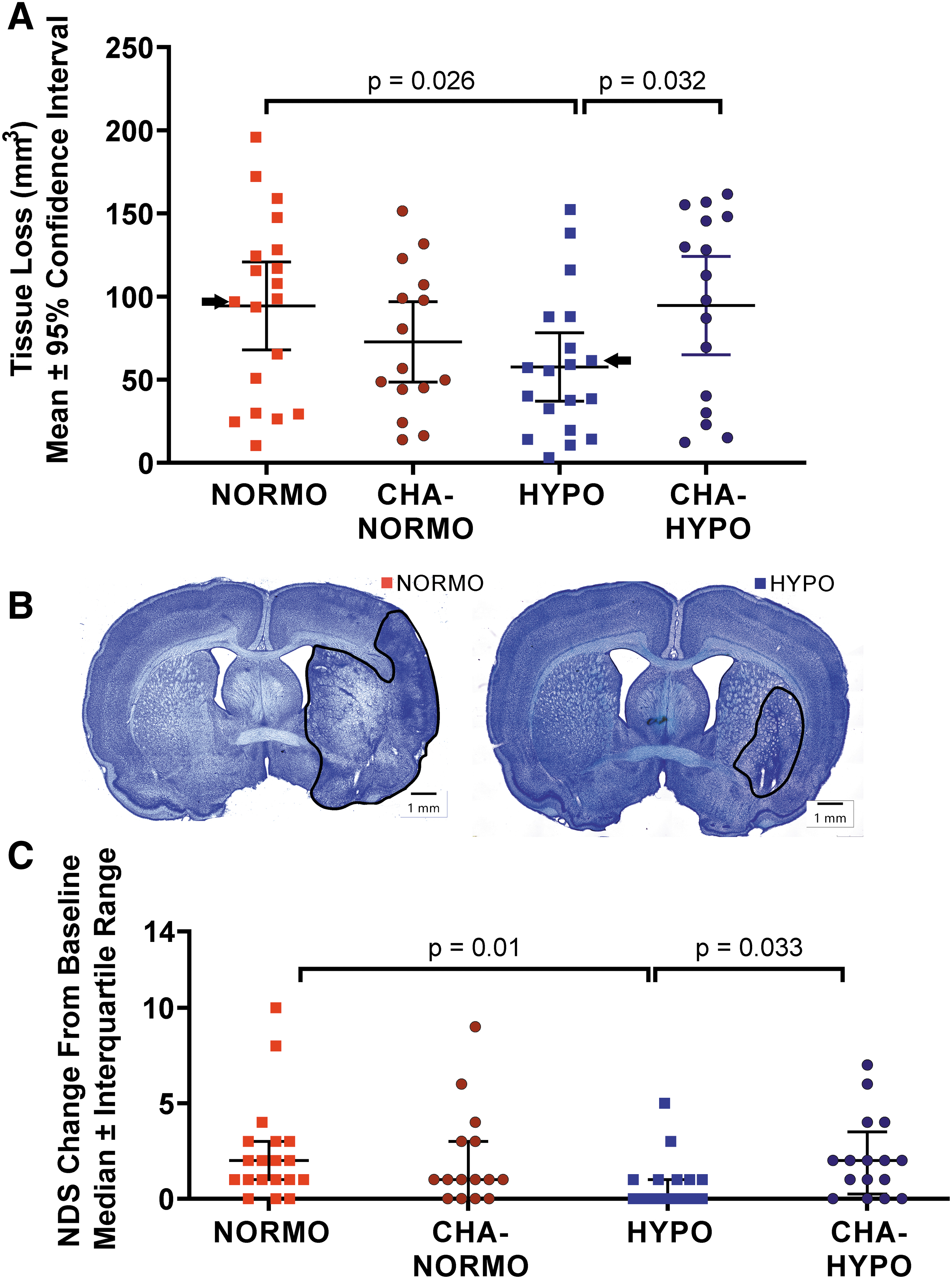
Histological and behavioral endpoints collected on day 7 post-MCAO.
In a post hoc comparison of all 31 CHA-treated animals compared with 19 NORMO animals, the difference in tissue loss remained statistically nonsignificant (p = 0.25). Moreover, HYPO animals had significantly less tissue loss than CHA-HYPO animals (p = 0.032). Thus, physical cooling was neuroprotective, but CHA treatment was not, regardless of cooling depth. Our stroke model caused injury from frontal brain regions (e.g., 3 mm anterior to bregma) to the hippocampus (e.g., 5.5 mm posterior to bregma).
Subcortical injury mostly occurred in the striatum, whereas cortical injury extended anterior and posterior to the striatum. Representative histology for average animals in the HYPO and NORMO groups is shown in Figure 3B.
HYPO, but not CHA-HYPO, animals showed improved behavioral outcomes
MCAO significantly worsened NDS at 7 days poststroke (baseline median = 0, day 7 median = 1; p < 0.0001; Fig. 3C). Among groups, a significant effect was detected (p = 0.01) owing to a significant difference between the HYPO and NORMO groups (HYPO group median = 0; NORMO group median = 2; p = 0.01). HYPO rats also performed better than CHA-HYPO rats (p = 0.033). There were no baseline differences (p = 0.78), and no other post-MCAO behavioral comparisons were significant (p > 0.99).
A post hoc comparison of all CHA-treated rats and NORMO animals revealed no significant differences between groups (p = 0.98). Thus, CHA did not affect behavioral outcomes, whereas physical cooling was beneficial.
Discussion
We expected to find clear evidence of neuroprotection in this study with CHA, but only physical cooling was neuroprotective. CHA did not significantly reduce post-MCAO injury or impairment, even though comparable cooling protocols, both systemic and locally applied, were clearly effective in past studies (Dumitrascu et al., 2016) and in the current study.
Notably, our physical TH paradigm was closely matched to the CHA-HYPO temperature profile in terms of induction rate, depth, duration, and rewarming rate. As a final point, we used large sample sizes that were determined a priori to overcome known variability in this stroke model and had 99.9% post hoc power to detect the difference in infarction volume between NORMO and HYPO animals.
We developed our physical TH protocol using data from a pilot study of 6 CHA-HYPO animals. Unexpectedly, the CHA-HYPO rats emerged from hypothermia ∼6 hours sooner than the HYPO group. Thus, the CHA-HYPO rats had 24 hours of hypothermia compared with 30 hours for the HYPO animals. This is likely due to individual differences in CHA metabolism as the individual CHA-HYPO temperature traces had more variation in time to target and depth of hypothermia compared with HYPO animals, which had more strict temperature control on our servoregulated system.
Although this heterogeneity is similar to clinical studies, it may have introduced additional variability into an already variable stroke model (Lyden et al., 2019). During the thermoregulation period, the core body temperature of animals in the CHA-NORMO group was ∼1°C lower than animals in the NORMO group. We maintained the CHA-NORMO animals at 36.5°C (as opposed to 37–37.5°C at baseline) to prevent stress and possible overheating during the temperature regulation process (Plahta et al., 2004).
Our study showed that a 1.5 mg/kg loading dose of CHA and 3 additional 1.0 mg/kg maintenance doses were unable to provide neuroprotective benefit following 2 hours of MCAO. This was surprising, especially because previous studies by our laboratory and others have shown that 21–24 hours of postischemic systemic TH (32–33°C) is sufficient to induce neuroprotection, even in permanent occlusion models (Clark et al., 2008; Yanamoto et al., 1996), and this neuroprotective efficacy was repeated in the current study.
Our behavioral data echoed our histological findings, in that only HYPO animals benefitted following MCAO. Moreover, many previous studies have demonstrated that CHA is efficacious in the context of global cerebral ischemic insults (Daval et al., 1989; von Lubitz et al., 1988). Reasons for the failure of CHA to induce neuroprotection are unclear, but may simply be the result of chance (estimates of treatment efficacy vary wildly among studies with low statistical power; Button et al., 2013), or it may be related to previous observations of side effects and/or individual differences in responsiveness to CHA, thereby potentially reducing drug efficacy in some animals (Jinka et al., 2015; Laughlin et al., 2018).
Indeed, some groups have attempted to counteract the cardiac side effects associated with systemic administration by giving an A1AR antagonist that does not cross the blood–brain barrier (Jinka et al., 2015). Additional studies may be required to better understand side effects associated with CHA and the factors that may reduce the efficacy of CHA as a treatment for focal ischemic stroke.
Moreover, although many studies have demonstrated neuroprotective efficacy with A1AR agonists following cerebral ischemic insults, there is evidence demonstrating mixed or null effects with respect to behavioral and histological outcomes following the acute administration of A1AR agonists in treatment of focal and global cerebral ischemic insults (Heron et al., 1994; Roussel et al., 1991; von Lubitz et al., 1988).
Overall, administering CHA appeared to be safe following MCAO: infarct sizes were not larger in either CHA-treated group, nor were there behavioral abnormalities or weight recovery differences at 1 week post-MCAO in either group. However, CHA on its own was unable to provide neuroprotection despite relatively ideal treatment onset and treatment parameters that were hypothesized to be neuroprotective based upon previous literature. An alternative consideration is that physical cooling may convey additional neuroprotective effects (Jackson and Kochanek, 2019), which we have remarked upon in our past work comparing systemic and local cooling (Clark et al., 2009).
Conclusions, Limitations, and Future Studies
Pharmacologically induced hypothermia using the adenosine A1 receptor agonist, CHA, was ineffective in treating a 2-hour focal ischemic stroke in rats, while physically induced cooling with similar parameters was actually beneficial. The comparison between NORMO and HYPO (and CHA-HYPO compared with CHA-NORMO) animals may be useful in future data syntheses (e.g., meta-analyses) as HYPO dosing issues remain unsolved.
As no single study can investigate the multitude of factors that may influence treatment responsiveness, future investigations may investigate whether deeper or more prolonged hypothermia induced by CHA is neuroprotective, potentially identifying more effective protocols. These studies should compare the mechanisms that are engaged by hypothermia, induced by physical or pharmacological means, and that led to differential histological and behavioral outcomes herein.
Notably, although it is possible (and likely) that both CHA treatments and physical hypothermia impacted cerebral blood flow in our study (Blood et al., 2003; Kurisu and Yenari, 2018; McBean et al., 1989), we did not assess cerebral blood flow and whether differences in blood flow were associated with efficacy differences in our study. Cerebral blood flow would be difficult to measure without introducing confounding effects into our experimental design (e.g., from anesthesia).
Moreover, blood flow data would be difficult to interpret given the reduction in metabolism that follows induction of torpor and hypothermia treatments as well as other temporally changing effects (e.g., shivering and reduction in general movement activity). On the latter, we showed overall reduced behavioral activity in CHA-HYPO, CHA-NORMO, and HYPO groups (Fig. 2B), and thus it is conceivable that this would correlate with reduced brain activity and therefore reduced cerebral blood flow.
Future studies should also take advantage of additional MCAO models (e.g., distal or thromboembolic models) as no single stroke model perfectly mimics the patient condition. Furthermore, potential effects of CHA on the various protective and pathophysiological mechanisms in stroke, particularly in the penumbra (Walther et al., 2022), should be considered. As future investigations are undertaken (i.e., with CHA or other hypothermia-inducing agents), additional factors should be investigated (e.g., impact of sex, age, and comorbidities; Dirnagl, 2019; Lapchak et al., 2013).
Finally, future studies may investigate whether CHA can be used synergistically with physical hypothermia methods as the induction phase of targeted temperature management is often difficult and time-consuming (Sessler, 2009). Perhaps the hypothermia process could be facilitated through pharmacological reduction of thermogenesis using A1ARs in tandem with physical cooling methods.
Footnotes
Acknowledgment
The authors would like to thank Xiuqing Ding for surgical contributions to this project.
Authors' Contributions
F.C. was involved in conceptualization, methodology, and writing—original draft preparation. L.J.L. and T.F.C.K. were involved in data collection and formal analysis of results, methodology, and writing—original draft preparation. Y.-G.H. was involved in conceptualization, methodology, data collection and formal analysis, and writing—reviewing and editing. A.M.B. was involved in conceptualization, methodology, funding acquisition, and writing—reviewing and editing. P.M. was involved in conceptualization, funding acquisition, and writing—reviewing and editing.
Author Disclosure Statement
A.M.B. is senior medical science advisor and cofounder of Brainomix, a company that develops electronic ASPECTS (e-ASPECTS). The other authors declare no competing conflicts of interest.
Funding Information
This project was funded, in part, by the Berlin Institute of Health at Charité–Universitätsmedizin Berlin (to A.M.B.) and the Leducq Foundation (to A.M.B. and P.M.). P.M. is funded by grants from the Else Kröner-Fresenius-Stiftung (2019_A34), the Volkswagen Stiftung (9A866), and the Einstein Foundation Berlin (EJF-2020-602, EVF-2021-619, and EC3R). P.M. is Einstein Junior Fellow and A.M.B. is Einstein Visiting Fellow, both funded by the Einstein Foundation Berlin. Y.-G.H. is funded by the U.K. Medical Research Council. F.C. is supported by the Canadian Institutes of Health Research (Grant number 166087). L.L. and T.F.C.K. are supported by the Canadian Graduate Studies–Doctoral Award from the Canadian Institutes of Health Research.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.