
Abstract
Background:
Alternative splicing, one of the sources of protein diversity, is often disturbed in cancer. Type 1 iodothyronine deiodinase (DIO1) catalyzes deiodination of thyroxine generating triiodothyronine, an important regulator of cell proliferation and differentiation. The expression of DIO1 is disturbed in different types of cancer. The aim of the study was to analyze the alternative splicing of DIO1 and its possible disturbance in renal cancer.
Methods:
Using real-time PCR, we analyzed 19 tissue samples (T) of renal cancer and 19 matched control samples (C) of the opposite pole of the kidney, not infiltrated by tumor, and 6 control samples (N) (nonneoplastic kidney abnormalities).
Results:
Cloning of DIO1 mRNA isoforms revealed 11 different transcripts, among them 7 new splice variants, not previously reported. The expression of all variants of DIO1 was dramatically (>90%) and significantly (p ≤ 0.0003) lowered in samples T compared to control samples C. The ratio of mRNA isoforms encoding DIO1 protein variants possessing or lacking the active center was lowered in samples T compared with control samples C, suggesting disturbed alternative splicing of DIO1. The expression of mRNA of splicing factors SF2/ASF (splicing factor-2/alternative-splicing factor) and hnRNPA1 (heterogeneous ribonucleoprotein A1), regulating 5′-splice site selection, was significantly but not proportionally lowered in samples T compared to samples C. The mRNA ratio of splicing factors SF2/ASF and hnRNPA1 correlated with the ratio of mRNA isoforms encoding DIO1 protein variants possessing or lacking the active center in controls C but not in samples T.
Conclusions:
Our results show that the expression and alternative splicing of DIO1 mRNA is disturbed in renal cancer, possibly due to changes in expression of splicing factors SF2/ASF and hnRNPA1.
Introduction
DIO1 activity has been detected in liver, kidney, pituitary, and thyroid (2). Previous reports have shown that the expression of DIO1 is disturbed in different types of cancer. DIO1 mRNA and activity were shown to be decreased in papillary thyroid carcinoma (7 –10), and increased in follicular adenoma and follicular thyroid carcinoma (7). DIO1 expression has been proposed as a differentiation marker of cancer cells (11,12).
The primary transcript of the DIO1 gene undergoes alternative splicing in liver, leading to several mRNA variants (13). It has been suggested that proteins encoded by alternatively spliced DIO1 mRNAs could contribute to DIO1 activity in pig kidney (14).
Alternative splicing is one of the mechanisms regulating gene expression in eukaryotes and significantly increases the diversity of encoded proteins. Numerous reports have shown that the process of alternative splicing is often disturbed in cancer, resulting in improperly spliced transcript variants (15). The selection of splice sites is strictly regulated by elements of the spliceosome; for instance, the alternative usage of 5′ splice sites is controlled by the ratio of splicing factors SF2/ASF (splicing factor-2/alternative-splicing factor) and hnRNPA1 (heterogeneous ribonucleoprotein A1) (16).
Clear cell renal cell carcinoma (ccRCC) is the most common subtype of renal cancer, representing 75–82% of primary malignancies of the kidney (17,18). The molecular basis of ccRCC is not entirely understood; however, several molecular markers have been proposed (17). It has also been reported that alternative splicing of several genes is disturbed in ccRCC (19 –22). In a previous work we showed that the expression of DIO1 mRNA and activity are lowered in ccRCC (23), but the alternative splicing of DIO1 in cancer has not been studied yet. Here, we show that alternative splicing of DIO1 primary transcript in kidney tissues results in different transcript variants. We also show that the expression and alternative splicing of DIO1 pre-mRNA is disturbed in ccRCC tissues leading to imbalanced expression of transcript variants. Since we also observed disturbed expression of splicing factors SF2/ASF and hnRNPA1, it seems likely that this may be the reason for the aberrant DIO1 splicing.
Materials and Methods
Tissue specimens
Tissue samples were obtained with the permission of the Ethics Committee of Human Studies (The Medical Centre of Postgraduate Education) from patients during unilateral nephrectomy performed due to kidney cancer (19 patients) and other nonneoplastic abnormalities (nephrolithiasis, renal cirrhosis, hydronephrosis, and injury—six patients).
The samples were divided into three groups: cancer tissues (19 samples “T”) and two types of control tissues—(a) the opposite pole of the cancer kidney with no signs of tumor infiltration (19 samples “C”) and (b) tissues from kidneys with other, nonneoplastic abnormalities (six samples “N”). Clear cell renal cell cancer was diagnosed upon histological evaluation of the excised tumors according to WHO criteria (24).
RNA isolation
Total RNA was isolated from ∼100 mg deep-frozen tissue with GeneMATRIX Universal RNA Purification Kit (EURx, Gdansk, Poland) according to manufacturer's instructions.
Real-time PCR
The isolated RNA served as a template in reverse transcription reaction, and the obtained cDNA samples were used in semiquantitative real-time PCR analyzes. Reverse transcription was performed with RevertAidTM H Minus First Strand cDNA Synthesis Kit (Fermentas, Vilnius, Lithuania) using oligo-dT primer, according to manufacturer's protocol.
Real-time PCRs were performed with QuantiFast SYBR Green PCR Kit (Qiagen, Hilden, Germany) according to manufacturer's protocol. Primers detecting variants of type 1 deiodinase mRNA were designed on the basis of sequences deposited in GenBank database (variant 1: NM_000792, variant 2: NM_213593, variant 3: NM_001039715, variant 4: NM_001039716). The sequences of the primers are shown in Table 1, and their positions relative to mRNA in Figure 1. Primers detecting SF2/ASF and hnRNPA1 mRNAs were designed on the basis of sequences NM_006924.4 and NM_002136.2, respectively. Real-time PCR conditions were as follows: initial denaturation at 95°C for 5 minutes, followed by 50 cycles (denaturation at 95°C for 10 seconds and combined annealing/extension at 60°C for 30 seconds); then melting curve analysis: 135 cycles (50°C with 0.3°C temperature increase in each cycle). Expression levels were normalized to values obtained for β-actin. The data are given as mean ± standard error (SE) (n = 6 for N, n = 19 for C, and n = 19 for T).
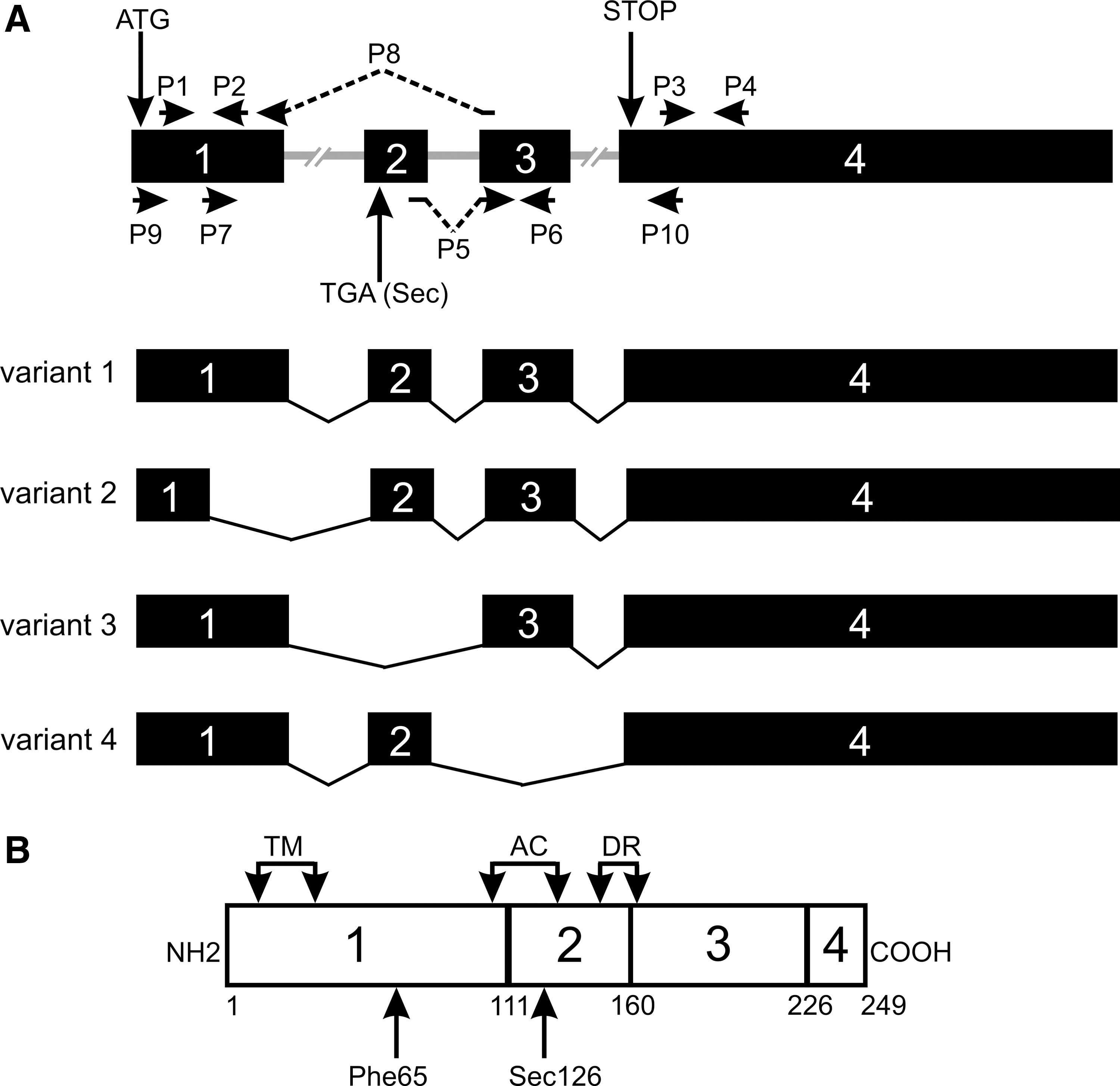
The gene, transcripts, and protein product for DIO1 according to GeneID: 1733 in Entrez Gene database. (
F, forward primer; R, reverse primer; DIO1, type 1 iodothyronine deiodinase.
Cloning of DIO1 splicing variants
Splicing variants of DIO1 were amplified from cDNA with a two-step PCR. The first reaction was performed with primers P9 and P4 (for sequences, see Table 1) under reaction conditions: initial denaturation at 95°C for 10 minutes, followed by 5 cycles (95°C for 20 seconds, 62°C for 20 seconds, and 72°C for 1 minute; the annealing temperature was decreased 0.5°C every cycle), then 40 cycles (95°C for 20 seconds, 57°C for 20 seconds, and 72°C for 1 minute), and final elongation at 61°C for 30 minutes. The PCR products were purified with GeneMATRIX PCR/DNA Clean-Up Purification Kit (EURx) and served as a template in the second round of PCR with primers P9 and P10 under reaction conditions: initial denaturation at 95°C for 10 minutes, followed by 45 cycles: (95°C for 15 seconds, 59°C for 30 seconds, and 72°C for 45 seconds), and final elongation at 61°C for 30 minutes. The PCRs were performed with Perpetual OptiTaq DNA Polymerase HOT START (EURx). The products of PCR were electrophoresed in 1% agarose gel stained with ethidium bromide. The products of the second round of PCR were purified with GeneMATRIX PCR/DNA Clean-Up Purification Kit (EURx), cloned into pGEM-T vector (Promega, Madison, WI), and transformed into Escherichia coli Fusion-Blue Competent Cells (Clontech, Mountain View, CA). Clones were checked with colony-PCR performed with primers SP6 and T7 specific for the pGEM-T vector, Perpetual OptiTaq DNA Polymerase HOT START (EURx), and under the following conditions: initial denaturation at 95°C for 10 minutes, followed by 40 cycles (denaturation at 95°C for 20 seconds, annealing at 55°C for 20 seconds, and extension at 72°C for 1 minute), and final elongation at 61°C for 30 minutes. The products of PCR were analyzed by agarose gel electrophoresis, and positive clones were used for isolation of plasmids with standard alkaline lysis procedure. Plasmids with cloned splicing variants were sequenced with Big-Dye terminator kit (Applied Biosystems, Foster City, CA) and primers SP6 and T7 on an ABI3100 machine.
In silico analysis
The obtained sequences were compared with GenBank reference sequence NC_000001.9–GI:89161185 using BLAST (
Statistical analysis
Normal distribution was tested using the Shapiro-Wilk test, followed by either Wilcoxon signed rank test or paired t-test; p < 0.05 was considered statistically significant. Pearson's coefficient of correlation was used to analyze the correlation between the ratio of DIO1 variants and SF2/ASF:hnRNPA1 mRNAs ratio.
Results
mRNA expression analysis
Four reference sequences of transcript variants of human DIO1 gene have been deposited in Entrez-Gene database: variant 1 (NM_000792.4), variant 2 (NM_213593.2), variant 3 (NM_001039715.1), and variant 4 (NM_001039716.1). The transcripts differ, among others, by the presence or absence of exon 2 encoding the active center of the enzyme and a fragment of exon 1 encoding amino acids responsible for substrate specificity (Fig. 1). The primers for real-time PCR analysis were designed to detect the following groups of transcripts: (A) all four variants (primers P3 and P4 located in exon 4, common to all transcripts); (B) transcripts with the longer version of exon 1 (i.e., variants 1, 3, and 4; primers P1 and P2); (C) variant 3 (lacking exon 2; primers P7 and P8); (D) transcripts with exon 2 and 3 (i.e., variants 1 and 2; primers P5 and P6).
The expression of all four groups of transcripts was detectable in all samples analyzed (Fig. 2). There were no significant differences in the expression of all four transcript groups between control samples C and N. However, the expression of transcripts was dramatically and statistically significantly decreased in samples T when compared with both types of control samples. The level of expression of mRNA groups A, B, C, and D in tumor samples versus control samples C was ∼4%, 9%, 7%, and 1%, respectively (p ≤ 0.0003) (Fig. 2).

Expression of type I deiodinase transcript variants. (
The observed decrease in expression of DIO1 mRNAs in tumor samples was not uniform (i.e., the expression level of each transcript groups was differently altered when compared to control samples). Therefore, as a result of such differences, a change in ratios of transcript variants in tumor samples when compared to relevant ratios in normal tissue must have occurred. Therefore, we calculated the ratio of transcript groups D (variants 1 and 2) and C (splice variant 3) in samples T, C, and N (Fig. 2E). The ratio, calculated as mean of D:C ratios in individual samples, was 0.6 ± 0.2 SE, 1.0 ± 0.1 SE, and 0.3 ± 0.1 SE in N, C, and T samples, respectively.
The imbalanced expression of the two groups of DIO1 transcript variants suggested disturbances in splice site selection. Therefore, we decided to analyze the expression profiles of mRNA of two splicing factors, SF2/ASF and hnRNPA1, known to participate in 5′ splice site selection (Fig. 3). The expression of mRNA for the two factors was similar in both types of control samples. However, in tumor samples we observed a decrease of SF2/ASF mRNA expression to 37 ± 9% SE and hnRNPA1 mRNA to 44 ± 11% SE of the respective levels in the control. The ratio, calculated as mean of SF2/ASF:hnRNPA1 mRNA ratios in individual samples, was 0.7 ± 0.1 SE, 0.8 ± 0.1 SE, and 0.6 ± 0.1 SE in N, C, and T samples, respectively (Fig. 3C).
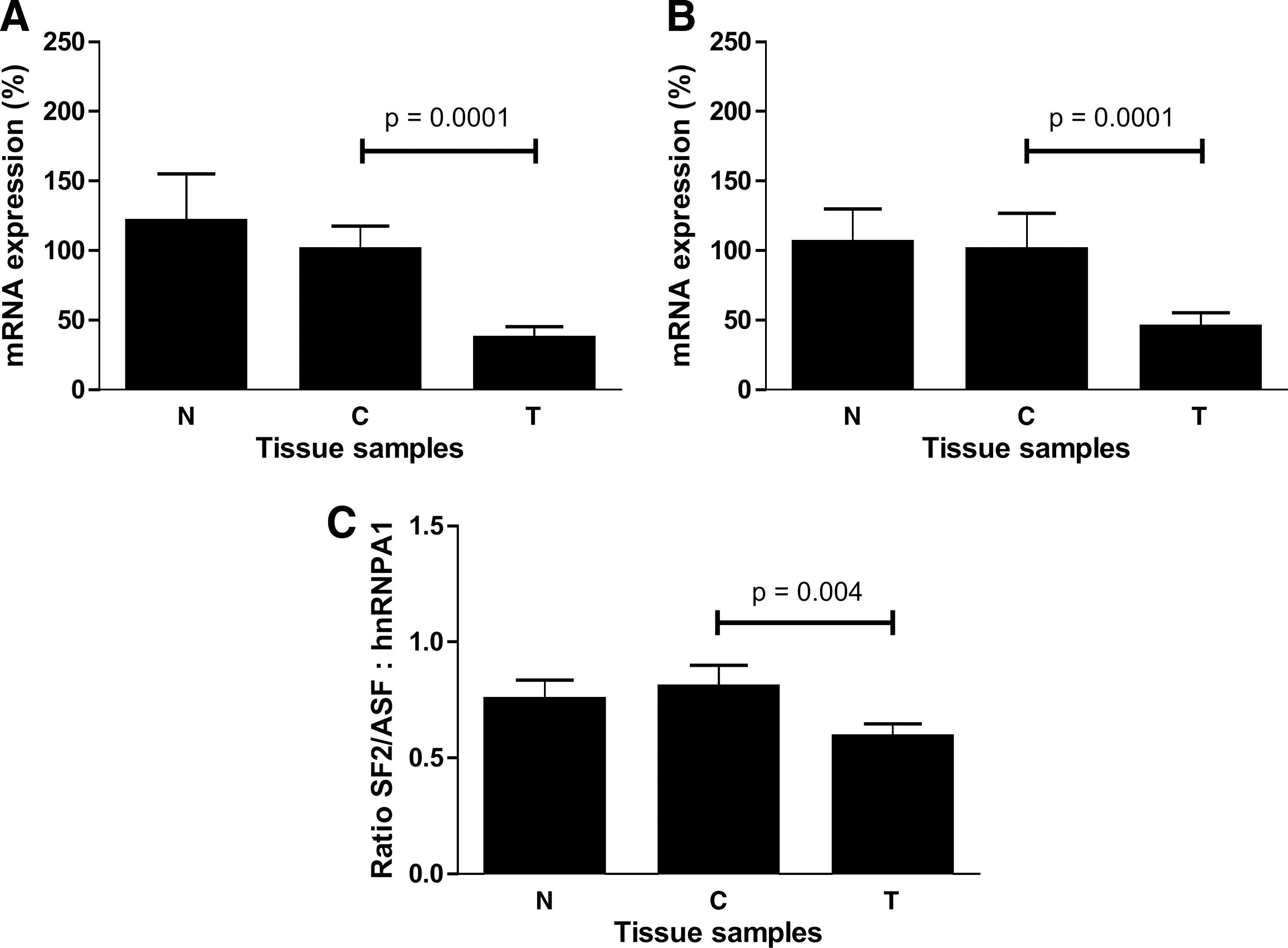
Expression of splicing factors (
To check if there is any relation between the expression of various transcripts of DIO1 and the ratio of the two splicing factors, we performed a correlation analysis of the ratios of these transcripts in individual samples C and T. We found a positive correlation (r = 0.4788; p = 0.0381) between the ratio of DIO1 transcripts possessing and lacking the region encoding the active center of the enzyme (group D:group C) and the ratio SF2/ASF:hnRNPA1 mRNAs in control samples (Fig. 4A). No such correlation was observed in samples T (Fig. 4B).

Correlation between expression ratios of DIO1 transcripts group D:group C and splicing factors SF2/ASF:hnRNPA1. (
Identification of DIO1 splicing variants
PCRs performed on cDNA from all samples revealed a complex pattern of bands, differing between patients and also different for samples C and T of a given patient (Fig. 5). The strongest bands corresponded to the expected sizes of the four transcript variants present in ENTREZ-GENE database (881 bp for transcript variant 1, 689 bp for variant 2, 737 bp for variant 3, and 681 bp for variant 4). However, we noticed also bands of lower as well as higher molecular weight. Cloning and sequencing revealed that the bands corresponded to known as well as new transcript variants of DIO1 gene (Fig. 6). Three of the seven not previously reported transcript variants were found exclusively in the tumor samples (numbers 9, 10, and 11 in Fig. 6). All of them lacked the region encoding the active center of the enzyme.
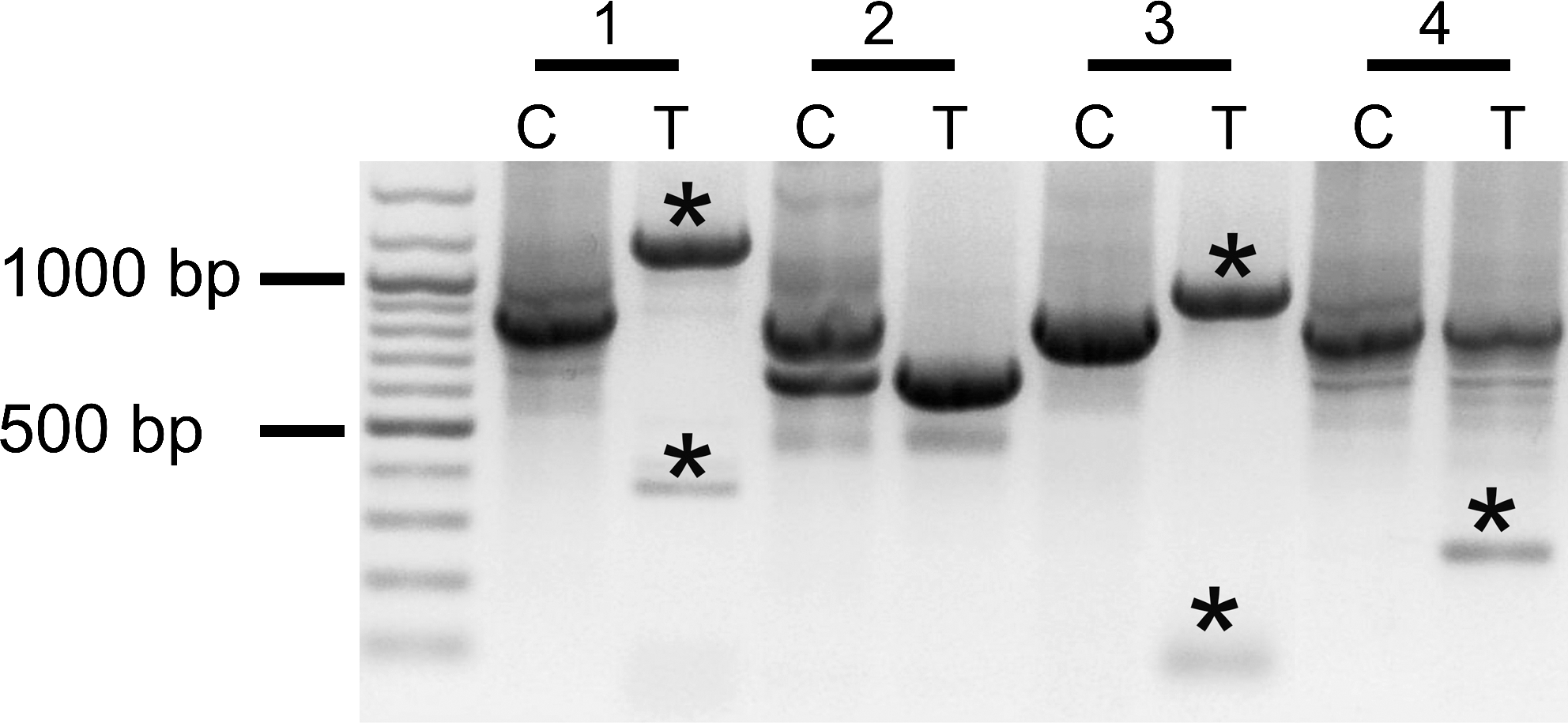
Various spliced isoformes of DIO1. Profiles from four representative patients showing differences between control samples C and cancer samples T and the presence of canonical as well as novel splicing variants (*).
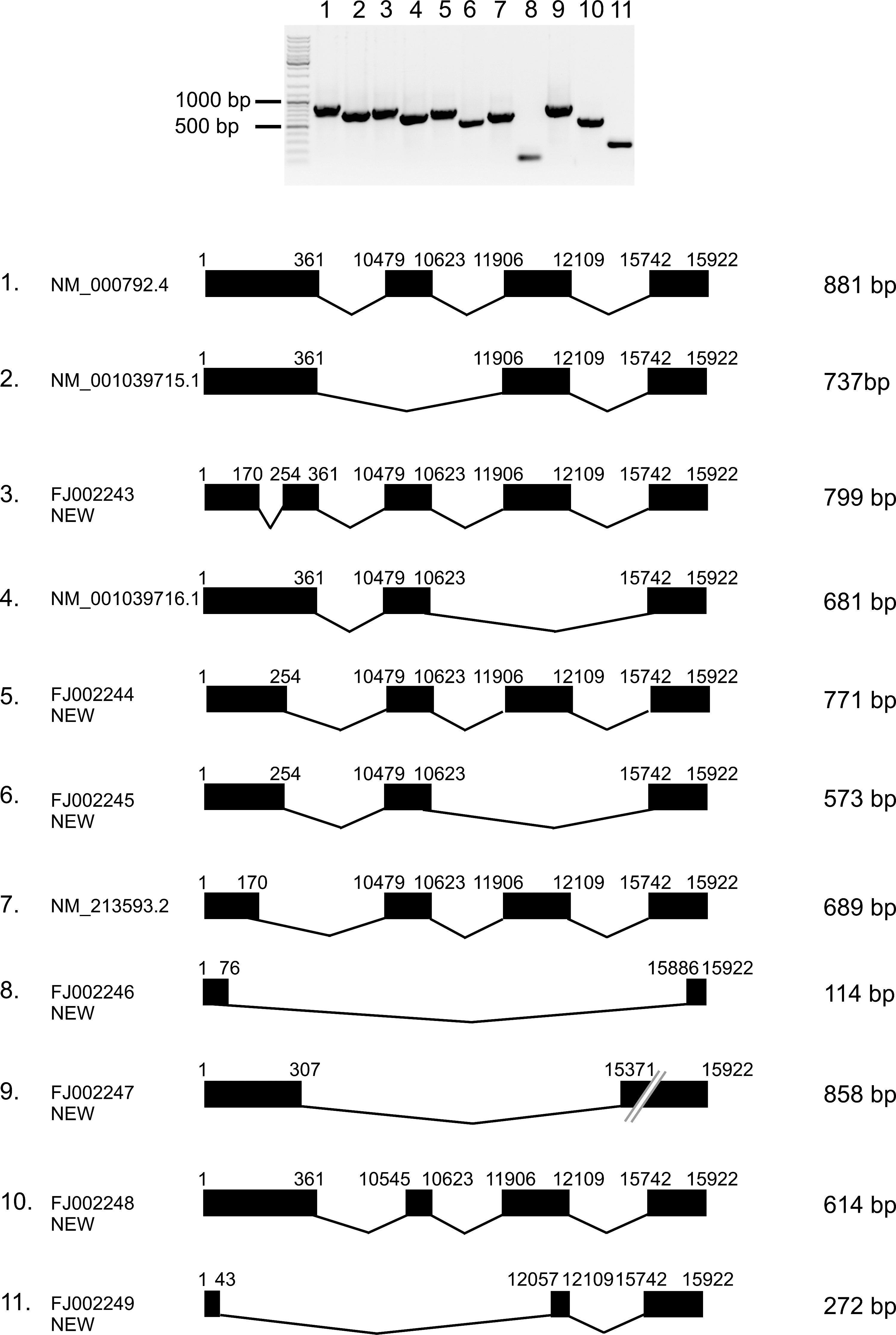
Splicing variants of DIO1 from control and cancer samples. Variants 1–8 were found in N, C, and T samples, variants 9–11 exclusively in cancer samples. Upper panel shows electrophoretic analysis of the cloned splicing variants. Lower panel shows the exonic structures. Exons are shown as black boxes. Numbers above exons refer to nucleotide positions in the DIO1 gene (GenBank locus: NC_000001.9, region 54132449–54149347).
Discussion
This is the first report analyzing the alternative splicing of type 1 deiodinase mRNA in cancer. We identified 11 splicing variants, among them 7 new ones, previously not reported. All of 11 splicing variants were identified in T samples, while only 8 of them occurred in C and N samples. Four transcripts cloned from control and tumor samples correspond to known DIO1 mRNA splicing variants, deposited in Entrez Gene database as reference sequences: transcript variant 1 (NM_000792.4), variant 2 (NM_213593.2), variant 3 (NM_001039715.1), and variant 4 (NM_001039716.1). Three other transcripts partially correspond to the sequences deposited as “related sequences,” cloned from human liver tissue: AY560382.1 (number 3 in Fig. 6), AY560379.1 (number 5 in Fig. 6), and AY560376.1 (number 6 in Fig. 6). However, clones identified in our study are longer in both upstream and downstream directions. We cannot exclude the possibility that this difference can result from different position of primers used in our study.
Three of the new transcript variants presented in this study were cloned exclusively from tumor samples (numbers 9, 10, and 11 in Fig. 6). Recent reports show that splicing variants of different genes can serve as markers of cancerogenesis (25,26). Also, DIO1 expression was proposed as a differentiation marker in cancer cells (11,12). Further analysis is needed to confirm if the transcripts identified in our study are specific for cancerous tissues and if they can suit as possible markers of cancerogenesis.
It is not known whether protein products of the alternative splicing variants of DIO1 exist in vivo and we were not able to check it due to the lack of specific antibodies. However, it has been suggested that the protein isoforms resulting from alternative splicing of DIO1 gene transcript could contribute to DIO1 activity in porcine kidney (13). Translation prediction shows that the cloned splicing variants can give proteins of different properties than those of the wild-type enzyme (Fig. 7). Our previous results (23) have shown that DIO1 activity poorly correlated with mRNA level in healthy renal tissues. This observation suggests significant posttranscriptional regulation of DIO1 expression. Interestingly, seven of the splicing variants cloned in this study possess a premature termination codon (PTC). In four variants (numbered 3, 8, 9, and 11 in Fig. 7) PTC is located more than 50 nucleotides upstream of the final exon–exon junction. Such alternative splice forms are believed to be recognized by nonsense-mediated mRNA decay mechanism (NMD) (27). NMD is generally considered a mechanism of degradation of PTC containing mRNAs and thus protecting cells from potentially harmful effects of abnormally truncated proteins. It was also suggested that NMD, coupled with alternative splicing, may serve as a regulatory mechanism of gene expression (28). Interestingly, DIO1 belongs to the group of selenoproteins, and it was found that NMD regulates the expression of selenocysteine-codon containing transcripts (29). In our previous work we showed that the activity of DIO1 in ccRCC samples is not detectable (23). Thus, although purely speculative, it is possible that what we observe in ccRCC tissue samples is a possible shift in alternative splicing, leading to production of PTC forms that are degraded by NMD, resulting in lowered DIO1 protein level. Pursuant to the above, the previously observed lowered activity of DIO1 in ccRCC would result from not only lowered transcription rate but also downregulated translation due to NMD of PTC variants.
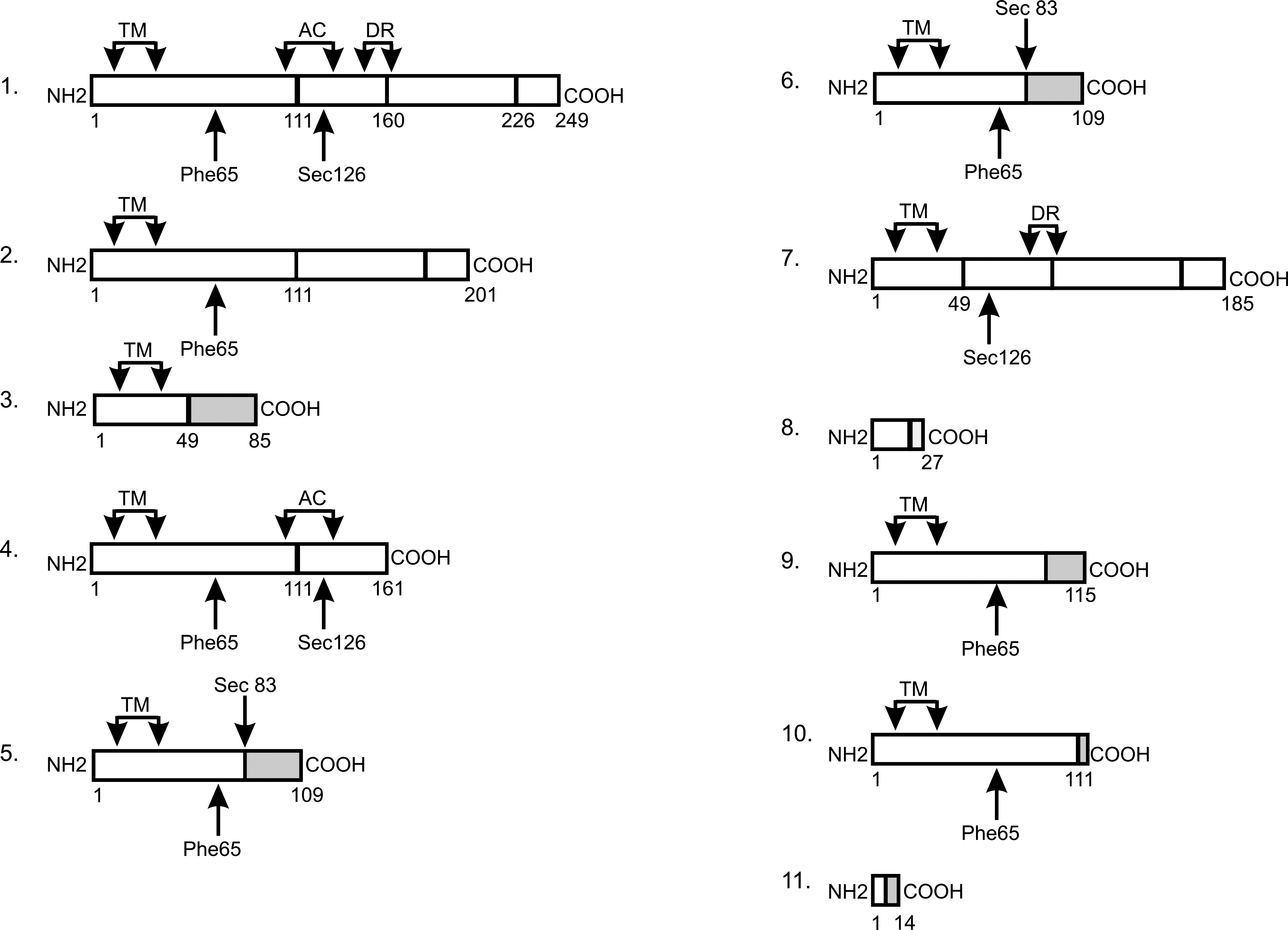
Translation prediction for the DIO1 splicing variants. The numbering refers to the numbers in Figure 6. Protein regions corresponding to exons are boxed. Regions of frameshift are shaded. Key regions of the enzyme are marked. TM, transmembrane region; AC, active center; DR, dimerization region; Phe65, rT3 substrate specificity; Sec126, catalytically active residue.
An interesting issue is whether the observed changes in DIO1 expression in ccRCC may effect the T3 levels in plasma or kidney tissues. According to previously published results, DIO2 but not DIO1 activity is the major source of serum T3 (30). In 2008, Panicker et al., however, showed that polymorphism in DIO1 leads to alteration in the ratio between circulating free T3 and free T4 (31). Unfortunately, the serum of patients is not available for analysis under this study; therefore, this problem remains for further research. Interestingly, analyzes performed recently on another set of tissue samples have shown that ccRCC tissue possess lowered T3 and variable T4 levels compared to nontumoral paired control samples (manuscript in preparation). Whether this coincides with lowered DIO1 activity remains to be clarified.
Results published previously have shown disturbances in expression and functioning of TRs in ccRCC (3,4). These observations, together with results presented in this paper suggest that the pathway of thyroid hormone action is disturbed in ccRCC. Since thyroid hormone is an important regulator of expression of oncogenes and tumor suppressors (5,6), it is possible that T3 pathway disturbance might be one of the origins of neoplastic transformation of ccRCC. This hypothesis, however, needs further investigation.
The analysis of ratios of groups of transcripts showed that the balance between transcripts encoding active (group D) and nonactive (group C) enzyme was disturbed in ccRCC samples. The expression levels of both groups of transcripts were similar in control samples C (Fig. 2E). However, the ratio of group D to group C transcripts in tumor samples was more than threefold lower than in control samples. Such a result suggests disturbed alternative splicing of DIO1 pre-mRNA in cancerous tissues leading to increase in skipping of exon 2. One of the mechanisms of alternative splicing is that of 5′ splice site selection (32) regulated by the ratio of splicing factors SF2/ASF and hnRNPA1 (16). When the level of hnRNPA1 is higher than that of SF2/ASF, distal 5′ splice site is preferred, which in the case of the analyzed DIO1 variants would mean the predominance of variants devoid of exon 2 (group C) over variants with this exon (group D). In our experiments the ratio of SF2/ASF:hnRNPA1 correlated with the ratio of group D:group C in control but not in tumor samples (Fig. 4), in which the expression of both SF2/ASF and hnRNPA1 was significantly, but not proportionally, lowered resulting in a decreased SF2/ASF:hnRNPA1 ratio. This suggests that alternative splicing of DIO1 could be regulated via SF2/ASF and hnRNPA1 and that this regulation could be lost in renal cancer. This hypothesis needs further research.
Previous reports have shown that the alternative splicing of other genes is also disturbed in ccRCC. It was shown that expression of specific splicing variants of survivin (19), CD-44 adhesion molecule (20), and TCF-4 gene (22) was related to tumor progression in ccRCC. These results, together with present study, suggest that disturbances in alternative splicing in renal cancer can result from common changes in splicing regulatory mechanisms. It was shown that SF2/ASF can function as a proto-oncogen and its overexpression can lead to immortal transformation of rodent fibroblasts (33). The same paper reported also that SF2/ASF is upregulated in various human tumors. The authors observed ≥2-fold overexpression of SF2/ASF mRNA in less than 5% clear cell renal cell carcinoma samples analyzed. In our study, however, all the analyzed samples had at least twofold decrease of expression. This discrepancy could result from primers we used in real-time PCR analysis, which detected solely transcript variant 1 of SF2/ASF, in contrast to Karni et al. (33), who detected as well transcript variant 1 as 2. It is known, however, that the active splicing factor SF2/ASF is synthesized from transcript variant 1, and it is doubtful if transcript variant 2 undergoes translation at all (34).
In conclusion, we show that expression and alternative splicing of type 1 deiodinase is disturbed in ccRCC. These disturbances are accompanied by changes in expression of splicing factors SF2/ASF and hnRNPA1. The new DIO1 splicing variants cloned exclusively from cancer samples suggest their use as potential molecular markers of renal cancer.
Footnotes
Acknowledgments
This work was supported by the Polish State Committee for Scientific Research Grants N401 3547 33 (to A.P.-W.) and 2P05A 128 26 (to A.N.), and the Medical Centre of Postgraduate Education Grant 501-2-1-22-37/07 to A.P.-W.
The authors would like to thank Professor Janusz Nauman for critical reading of the article.
Disclosure Statement
The authors declare that no competing financial interests exist.
This work was presented in part at the 9th European Congress of Endocrinology, April 18–May 2, 2007, Budapest, Hungary.