
Abstract
Background:
If methods of differentiating stem cells into thyrocytes can be perfected, they may provide a ready source of normal thyrocytes for basic research and clinical application. We developed a novel culture method capable of differentiating mouse embryonic stem (ES) cells into thyroid follicular cells.
Methods:
E14 mouse ES cells were allowed to differentiate into embryoid bodies and then stimulated with thyroid-stimulating hormone, insulin, and potassium iodide. The resulting differentiated cells were observed for expression of thyrocyte-specific mRNA transcripts with reverse transcriptase (RT)–polymerase chain reaction. To definitively identify thyrocytes, we simultaneously observed the thyrocyte-specific proteins, thyroid transcription factor-1 and PAX-8, with dual-color immunofluorescent labeling. The cells were further characterized by electron microscopy.
Results:
The ES cells were successfully differentiated into thyrocytes. Differentiated cells expressed PAX-8, thyroid-stimulating hormone receptor, sodium/iodide symporter, thyroperoxidase, and thyroglobulin mRNAs, and coexpressed thyroid transcription factor-1 and PAX-8 proteins. The extent of differentiation was further explored by electron microscopy, which showed that differentiated cells had ultrastructural features similar to adult human thyrocytes, whereas the cells from unstimulated cultures were mostly disintegrated and lacked developed organelle structures.
Conclusions:
These data show that E14 mouse ES cells can be differentiated into thyrocytes by culturing with thyroid-stimulating hormone, insulin, and potassium iodide. The development of reliable methods to produce thyroid cells from ES cells is important to future research in thyroid biology and medical applications.
Introduction
The differentiation of the thyroid lineage is a high priority because the thyroid is a critical hormone-producing gland that is involved in various pathologies, including goiter, thyroid adenomas, and hypothyroidism (8,9). Basic understanding of the biology of the thyroid has been hindered by the lack of a good model system. Use of thyroid lineage cells derived from ES cells may provide a convenient model system to study thyroid biology and provide the foundations for research into treatments for thyroid diseases. Further, ES cell–derived thyrocytes may themselves be used as a treatment for diseases of the thyroid.
One successful culture technique using human recombinant thyroid-stimulating hormone (TSH) has been developed for directing differentiation of thyroid follicular cells from the CCE mouse ES cell line (2,7). This article presents a different set of culture conditions using TSH and insulin to differentiate thyroid follicular cells from the E14 mouse ES cell line. Many of the features of the thyroid follicular cells derived from the two techniques are similar. Notably, the present study uses a different ES cell line, which has not previously been shown to be able to differentiate into thyrocytes. This study also examined the ultrastructure of the induced cells and showed expression of a key thyroid transcription factor, thyroid transcription factor-1 (TTF-1). The previous studies had varying successes at detecting expression of thyroglobulin (Tg), the precursor to the thyroid hormones triiodothyronine (T3) and tetraiodothyronine (T4). The cells induced in this work produced Tg as well.
In the method presented here, TSH and insulin were used together to induce differentiation of thyrocytes from ES cells. Thyrocytes are characterized by the expression of some specific genes such as Tg, thyroperoxidase (TPO), TSH receptor (TSHR), and the sodium/iodide symporter (NIS). These genes were chosen for reverse transcriptase (RT)-PCR analysis to detect thyrocytes. It is known that thyroid tissue-specific transcriptional regulation is mediated by a set of three transcription factors: TTF-1, thyroid transcription factor-2 (TTF-2), and Pax-8 (a member of the pax family of paired domain-containing genes) (10 –12). TTF-1 and Pax-8 are coexpressed only in thyroid follicular cells or thyroid precursors (13,14). In our study, double-color confocal immunofluorescence of TTF-1 and Pax-8 proved that thyrocytes were induced in our ES culture system.
The ES cell–derived mouse thyroid model systems should be invaluable to furthering research into thyroid and ES cell biology. In the future, the elements of these mouse ES cell techniques may be extended to developing techniques for differentiating human stem cells into thyrocytes for basic research and clinical use.
Materials and Methods
Mouse ES cell culture
E14 mouse ES cells (a gift from Dr. Xu, Harvard University, and preserved in the Cord Blood Library of the Second Affiliated Hospital of Sun Yat-Sen University) were cultivated on a feeder layer of mitomycin C–inactivated mouse embryonic fibroblasts. The cultivation medium consisted of Dulbecco's modified Eagle's medium (Gibco, Grand Island, NY) supplemented with 15% ES cell–qualified fetal bovine serum (FBS) (Gibco), 2 mM L-glutamine (Gibco), 1.5 × 10−4 M β-mercaptoethanol (Sigma, St. Louis, MO), 0.1 mM nonessential amino acids (Gibco), and 10 μg/mL leukemia inhibitory factor (LIF) (Sigma). ES cells were divided rapidly, were monitored daily, and were passaged every 2 days in a 1:3 ratio.
Adult human thyrocytes
As controls, human thyroid cells were collected from the biopsy of an adult female patient diagnosed with nodular goiter who was undergoing bilateral subablation of her thyroid. The normal thyroid tissue was collected in a 50 mL centrifuge tube containing RPMI-1640 medium. The human thyroid cells were then purified and cultured by routine methods. The procedure was approved by the hospital Institutional Review Board, and informed consent was obtained from the patient.
Differentiation of ES cells
To induce formation of EBs, ES cells were trypsinized and pipetted several times to obtain single-cell suspensions. The singe-cell ES cell suspension was seeded into the semisolid methylcellulose (MC) medium supplemented with 15% FBS, 2 mM L-glutamine, 1.5 × 10−4 M β-mercaptoethanol, 0.1 mM nonessential amino acids, and 0.25 μM ascorbic acid. MC medium was prepared as follows: a stock MC solution was prepared from 11 g MC (Gibco) dissolved in triple distilled water to a final volume of 500 mL, autoclaved, and stored at 4°C; the MC stock was mixed 1:1 with 2 × Iscove's modified Dulbecco's medium. Cultures were maintained in a humidified chamber with a 5% CO2/air mixture at 37°C. During culture, medium was replaced when the pH indicator in the medium changed color, typically every 3 days. Cultures were then observed for formation of EBs.
To induce differentiation into thyrocytes, the cultures containing EBs were further supplemented with 10 μg/mL human insulin (Sigma) and 1 mU/mL human TSH (Sigma) and allowed to grow for 6 days. After culturing for 6 days, 5–10 EBs per well were transplanted onto 0.1% gelatin-coated six-well plates in liquid Iscove's modified Dulbecco's medium containing 15% FBS, 2 mM L-glutamine, 1.5 × 10−4 M β-mercaptoethanol, 0.1 mM nonessential amino acids, 10 μg/mL insulin, and 1 mU/mL TSH. Transplantation was performed by pipetting the MC medium, containing EBs, into a centrifuge tube and mixing it with phosphate-buffered saline (PBS); upon settling, the EBs separated into the lower layer of PBS and could be isolated.
The plated 6-day-old EBs were allowed to differentiate for a further 11 days. The EBs were thus grown for 17 days (designated day 1 through day 17), the same amount of time they would have differentiated in vivo. To provide an environment permissive for thyroid hormone production, on day 13, after the expected time of expression of the NIS, the medium was further supplemented with 4 × 10−6 M potassium iodide (KI).
In some experiments, EBs were allowed to form and further differentiate in the absence of insulin, TSH, and KI. Differentiated cells grown with insulin, TSH, and KI were referred to as TSH-positive differentiated cells, whereas the cells grown without these factors were referred to as TSH-negative differentiated cells.
RNA isolation and gene expression analysis
Total RNA was collected from mouse ES colonies, days 8 and 10 TSH-positive differentiated cells using Trizol, and was reverse transcribed into cDNA using a Thermoscript™ RT-PCR system (Invitrogen, Carlsbad, CA). Polymerase chain reaction (PCR) was performed using standard protocols with Taq polymerase (Invitrogen). Amplification conditions were as follows: initial denaturation at 94°C for 3 minutes followed by denaturation at 93°C for 45 seconds, annealing at 53–65°C for 30 seconds, extension at 72°C for 45 seconds, and a final extension at 72°C for 10 minutes. The amount of cDNA in each sample was normalized using β-actin as a control. The amplified PCR products were separated by gel electrophoresis, and the identity of each PCR product was confirmed by direct sequencing. Each PCR product matched its published gene sequence. Forward and reverse primer sequences from 5′ to 3′ direction and the length of the amplified products were as follows: Oct4, GGCGTTCTCTTTGGAAAGGTGTC and ACCACATCCTTCTCTAGCCCTCGA (313 bp); PAX-8, TGCCTTTCCCCATGCTGCCTCCGTGTA and GGTGGGTGGTGCGCTTGGCCTTGATGTAG (298 bp); TPO, TGCCAACAGAAGCATGGTCAAC and GCACAAAGTTCCCATTGTCCAC (425 bp); Tg, TGGGATGTGAAAGGGGAATGGTGC and GTGAGCTTTTGGAATGGCAGGCGA (418 bp); TSHR, GAGTGTGCGTCTCCACCCTG and TTCCAGCCGCTGCAGAGTTGC (213 bp); NIS, GCTCTCATCAGCTACCTAACTG and CTCAGAGGTTGGTCTCAACATC (244 bp); β-actin, TTCCTTCTTGGGTATGG and GAGCAATGATCTTGATCTTC (200 bp).
Dual-color immunofluorescence
Adult human thyrocyte primary cultures or differentiated day 15 EB cultures were trypsinized and pipetted to form single-cell suspensions, then seeded onto 24-well plates in the same type of medium in which they had previously been cultured, and were allowed to attach for 24 hours and form monolayers suitable for immunofluorescent imaging. The adherent cells were then processed for immunofluorescent observation. At room temperature, the cells adhering to the plates were fixed in 95% ethanol for 10 minutes, permeabilized in 0.1% Triton X-100 in PBS for 3 minutes, and incubated in 5% bovine serum albumin (BSA) for 30 minutes, before being incubated overnight at 4°C with a mixture of primary antibodies diluted in 1% BSA in PBS. The cells were then washed in PBS, incubated for 30 minutes with a mixture of Cy3-tagged rabbit anti-mouse (Chemicon, Temecula, CA), and fluorescein isothiocyanate–tagged rabbit anti-goat secondary antibodies (KPL, Gaithersburg, MD) diluted 1:200 in 1% BSA in PBS, and given a final PBS wash. The primary antibodies were mouse monoclonal anti-TTF-1 antibody from Neo Markers (Lab Vision, Fremont, CA) and goat anti-PAX8 (Santa Cruz, Santa Cruz, CA).
Images were collected with a Zeiss LSM 510 (Jena, Germany) confocal laser scanning microscope. Image pairs were collected in both red and green channels and merged using the software provided with the microscope.
Electron microscopy
Cells from day 17 cultures or adult human thyrocyte primary cultures were trypsinized and centrifuged to form a pellet. The cell pellets were processed as follows: fixed for a minimum of 4 hours in PBS containing 2.5% glutaraldehyde and 2% paraformaldehyde; two 10-minute PBS washes; postfixed 1 hour in PBS containing 1% osmium tetroxide; three 10-minute PBS washes; sequentially dehydrated in 50%, 70%, 80%, 95%, and 100% ethanol; permeated for 1 hour in a 1:1 mixture of acetone and resin; permeated with 100% resin for at least 8 hours; embedded in capsules with freshly prepared resin; and polymerized at 60°C for at least 8 hours. The resulting tissue blocks were semi-thin sectioned under a light microscope or ultrathin sectioned with an ultramicrotome. Ultrathin sections were negative stained in uranyl acetate and lead citrate and observed with an electron microscope.
Results
ES cell colonies and differentiation
E14 mouse ES cells were maintained in the undifferentiated state by coculturing with mitomycin C–inactivated mouse embryonic fibroblasts and then adding LIF to obtain pure ES cells. Morphologically, the ES cells first formed a monolayer on top of the feeder layer, and later, as the ES cells divided, began to form into separate, rounded mounds of cells (Fig. 1). Upon transfer to permissive culture conditions in a suspension culture lacking LIF, the ES cells started to differentiate into EBs (Fig. 2). When these EBs were transplanted onto the gelatin-coated plates for further differentiation with or without TSH and insulin, the EBs began attaching to the dish and differentiating into various types of specialized cells (Fig. 3). The differentiating cells are seen as morphologically different cells surrounding the EBs, migrating out of the EBs onto the culture dish.
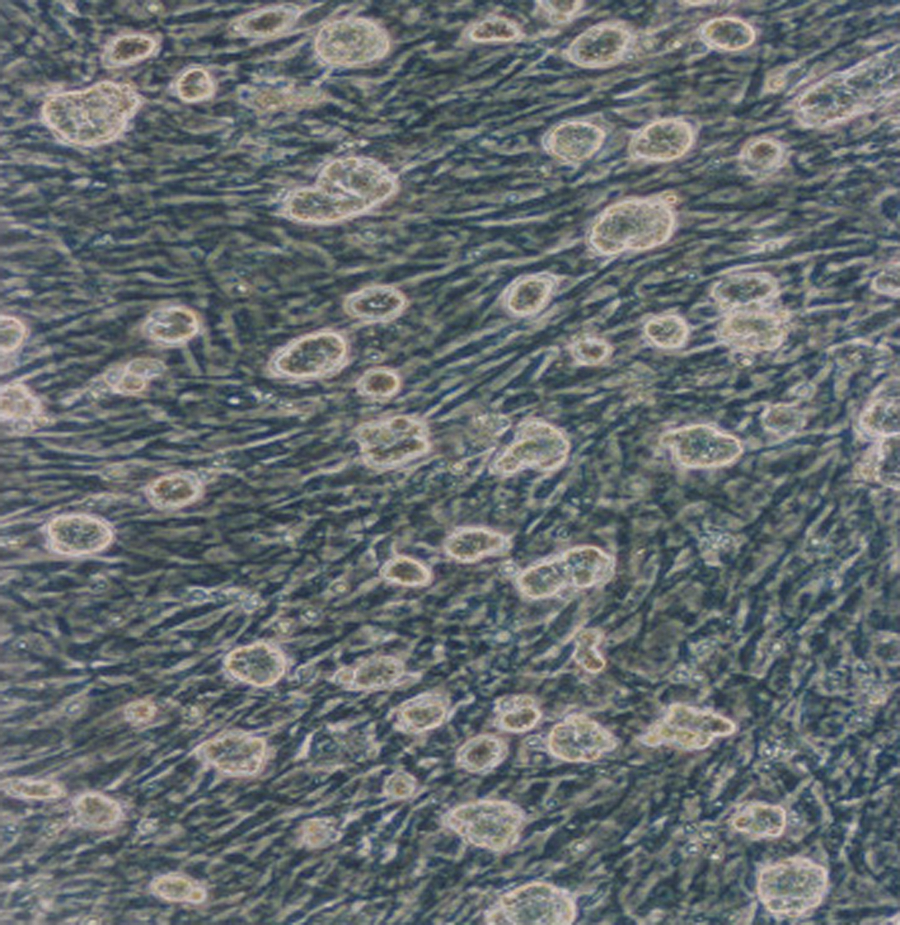
The ES cells were cultivated on a feeder layer of mitomycin C–inactivated mouse embryonic fibroblasts, and grew into rounded, multilayered clusters. Magnification: × 100. ES, embryonic stem.
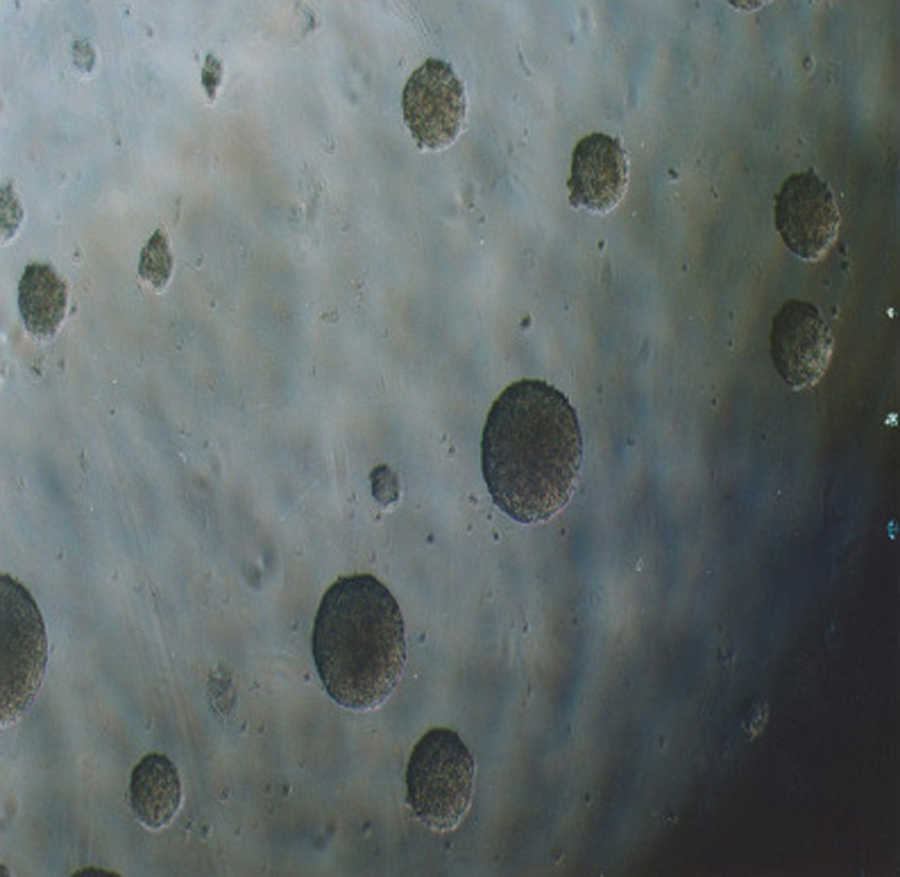
The ES cells started to differentiate into embryoid bodies when cultured in suspension without leukemia inhibitory factor. Magnification: × 40.

Morphologically differentiated cells are seen migrating out of the day 10 embryoid bodies onto the culture dish surface. Magnification: × 40.
Expression of thyrocyte-specific mRNA
To assess whether thyrocytes had differentiated in the EB cultures, RT-PCR was used on the total RNA from each culture to test for the presence of mRNA from genes known to be expressed by either undifferentiated cells or thyrocytes. In the undifferentiated state, ES cell cultures expressed the undifferentiated stem-cell marker Oct4, but no thyroid cell markers. In RNA samples from day 8 cultures, the TSH-positive differentiated cells expressed the thyrocyte markers PAX-8, NIS, TSHR, TPO and Tg, but not Oct4; and in RNA samples from day 10 cultures, the same expression pattern was seen, with Tg and TPO expressed more markedly (Fig. 4). All cultures expressed the β-actin control. The findings from these expression studies indicate that ES cells have the potential to differentiate into the thyrocyte lineage in vitro. However, ES cells are able to differentiate into all cell types and different cell types are known to express the PAX-8, NIS, and TSHR; it is conceivable that these mRNA were produced by other cell types that differentiated in culture.
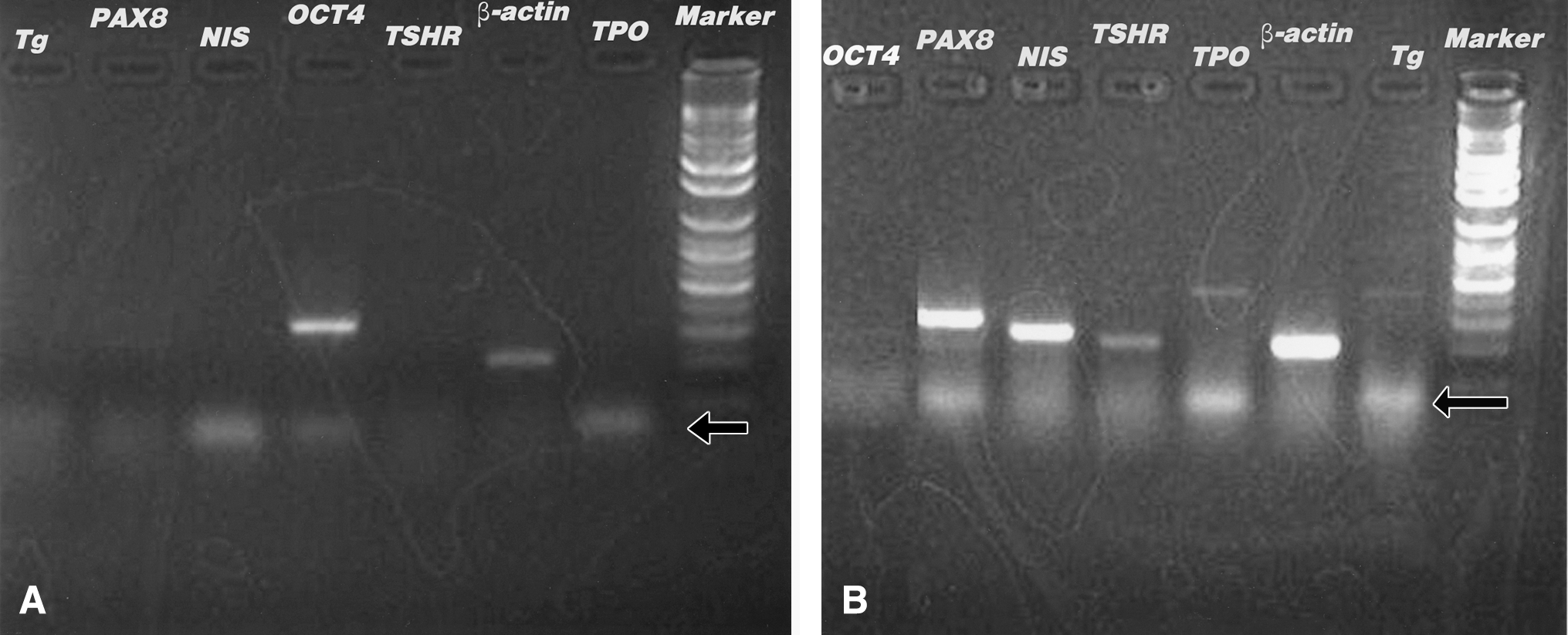
RT–polymerase chain reaction was performed for β-actin (expected product 200 bp) as a control and as tests: Tg (418 bp), PAX-8 (298 bp), NIS (244 bp), Oct4 (313 bp), TSHR (213 bp), and TPO (425 bp). ES cells expressed the undifferentiated stem-cell marker Oct4, but no thyroid cell markers (

Dual-color immunofluorescence
To conclusively demonstrate that the thyrocytes differentiated from the ES cells, dual-color immunofluorescence was used to observe the expression of both TTF-1 and PAX-8 proteins in individual cells, something unique to thyrocytes. Some cells from the day 15 TSH-positive culture coimmunostained with both anti-TTF-1 antibody and anti-PAX-8 antibody, whereas other cells from the TSH-positive culture stained with only one antibody. Among cells from the TSH-negative culture, only rare cells were stained with any antibody. In the control, all the adult human thyrocytes stained simultaneously with the two antibodies. Representative results are shown in Figure 5. Coexpression of TTF-1 and PAX-8 definitively identified thyrocyte lineage cells in the TSH-positive culture. Without the presence of TSH, insulin, and KI, thyrocytes did not develop in our culture system.
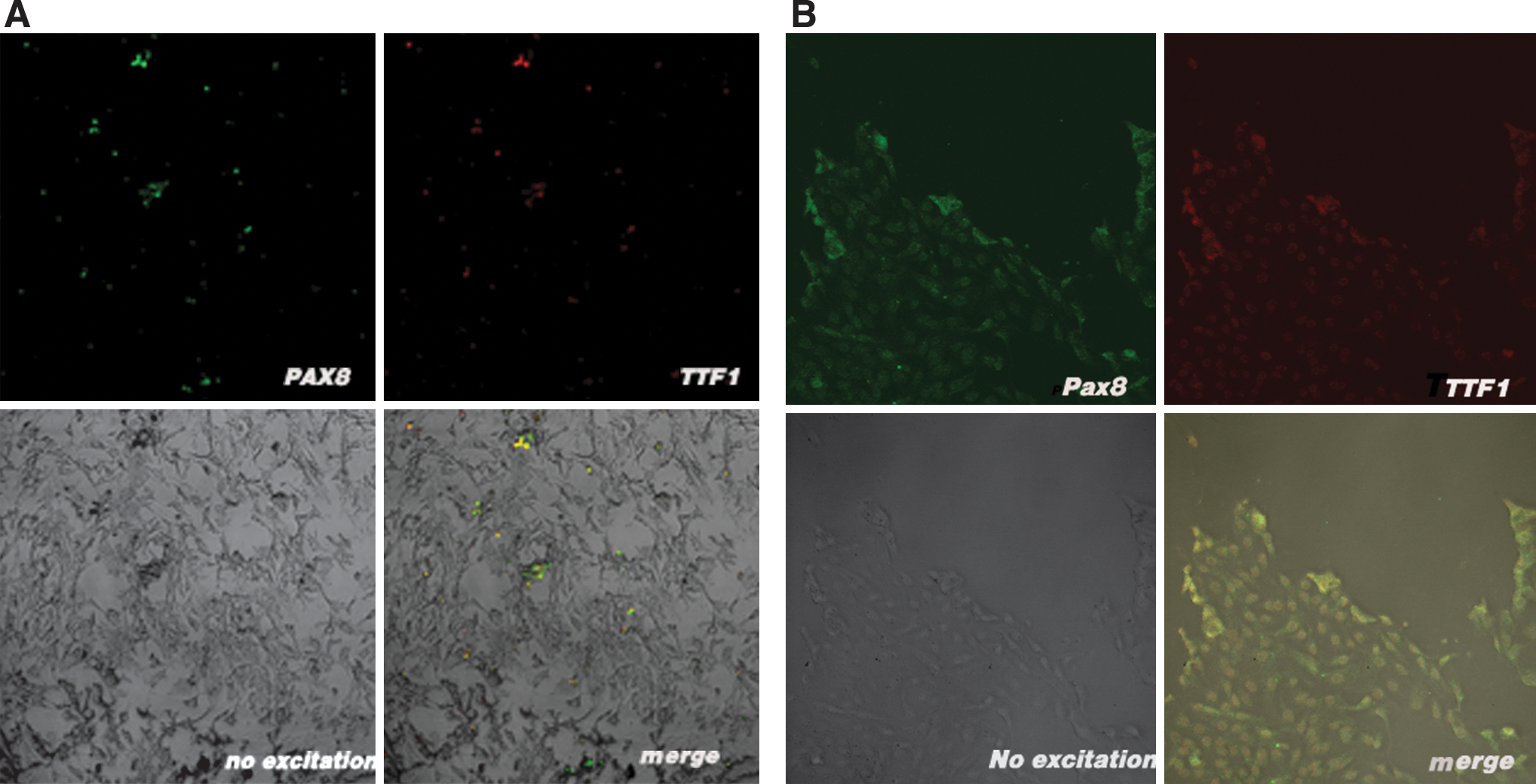
Confocal images of cells stained simultaneously with anti-TTF-1 and anti-PAX-8 antibodies. Some TSH-positive differentiated cells coimmunostained for both TTF-1 and PAX-8 (

Electron microscopy
The ultrastructure of adult human thyrocyte cells was compared with that of the cells from day 17 TSH-positive and TSH-negative cultures (Fig. 6). The adult human thyrocytes displayed microvilli (MV), rough endoplasmic reticulum (RER), Golgi bodies, and secretory granules. Cells from TSH-positive cultures also displayed MV, RER, and large Golgi bodies, but no secretory granules. No cells from the TSH-negative cultures displayed the array of MV, RER, and Golgi bodies seen in the other samples; instead, many apoptotic bodies and cell fragments were present, and few organelles were seen in the intact cells.

Microvilli (MV), rough endoplasmic reticulum (RER), and Golgi bodies (GOL) were remarkably visible in some TSH-positive differentiated cells (magnification: (
Discussion
The pluripotent nature of mouse ES cells was formally demonstrated by their ability to contribute to all tissues of adult mice, and studies during the past 20 years have led to the development of appropriate culture conditions and protocols for the generation of a broad spectrum of cell lineages (3 –6). Two studies have focused on differentiation of CCE strain mouse ES cells into thyrocytes (2,7). This study indicated the feasibility of inducing differentiation of E14 strain mouse ES cells into thyrocytes under different culture conditions and also explored the ultrastructure of the differentiated cells.
The culture conditions used in this study were intended to simulate the developmental environment of the mouse thyroid based on what is currently known. TSH and insulin were intended as thyrocyte differentiation inducers. TSH is the main hormone controlling adult thyroid gland physiology and is important during thyroid development. In the absence of either TSH or a functional TSHR, the thyroid gland develops to a normal size, whereas the expressions of TPO and the NIS are greatly reduced (8,15,16). TSH was used as the sole differentiating factor during induction of thyrocytes from CCE mouse ES cells. Besides TSH, insulins such as insulin or insulin-like growth factor I) have been associated with thyroid development; studies have shown that insulin and insulin-like growth factor-I regulate a thyroid-specific nuclear protein that binds to the Tg promoter (9,17). To further provide an environment suitable for thyrocyte development, KI was added to the medium after the cells were presumed capable of iodine uptake via the NIS. Iodine is necessary for the production of the primary thyroid gland products, the thyroid hormones T3 and T4.
RT-PCR analysis showed that the cells in the TSH and insulin-stimulated ES cultures had differentiated (having lost Oct4 mRNA expression) and expressed the thyroid-associated transcripts PAX-8, TSHR, NIS, TPO, and Tg by day 8. Dual-color immunofluorescence confirmed that PAX-8 protein was being coexpressed with TTF-1 protein in many cells on day 15, demonstrating that thyrocytes existed in the culture. These thyrocytes were likely the source of the thyroid-associated mRNA transcripts.
It is known that TTF-1 and PAX-8 are expressed at the onset of thyroid gland formation at E8–9.5 during mouse embryogenesis, and that TSHR, TPO, NIS, and Tg are expressed at E14 only when the gland reaches its final location (18). In contrast, TSHR, TPO, NIS, and Tg were expressed by day 8 in our culture conditions. Reasons for the difference in timing may be that these mRNAs were expressed in non-thyroid cells induced in culture which express the genes earlier; that expression of these genes is inhibited by other cells in normal embryos in vivo; or that the differentiation and development in culture is not perfectly parallel to normal in vivo developmental timing.
Our results were similar to the thyrocyte differentiation of the CCE mouse ES cell line by Lin et al. (7) and Arufe et al. (2). Both experiments showed mRNA expression of PAX-8, TSHR, NIS, and TPO. Lin et al. (7) had weak expression of Tg, whereas Arufe et al. (2) were unable to detect Tg mRNA expression. CCE strain ES-derived thyrocytes express TSHR, NIS, PAX-8, and TTF-2 proteins (2,7). This study demonstrated Tg mRNA expression as well as all other previously studied mRNA or proteins except TTF-2; this study is the first to demonstrate TTF-1 protein expression in ES-derived thyrocytes. Arufe et al. speculate that different ES cell lines have varying propensities to differentiate into certain cell lineages (2). This study shows that the E14 ES cell line is also capable of differentiating into thyrocytes.
Some TSH-positive cells stained with both anti-TTF-1 and anti-PAX-8 antibodies, conclusively identifying them as thyrocytes. However, other cells in this culture stained with only one or the other antibody. It is known that the transcription factor TTF-1 is expressed in embryonic diencephalons, lung, and thyroid, whereas the transcription factor PAX-8 is expressed in kidney, the developing excretory system, and thyroid (13,14). Cells that were only stained by one antibody could have been from these other lineages or could have been incompletely differentiated thyrocytes because only rare cells coexpressed TTF-1 and PAX-8 proteins in TSH-negative differentiated cells. Although the two antibodies stained all the adult human thyroid cells, it is conceivable that our inducing approach induced ES cell differentiation into thyrocytes in vitro.
Having successfully induced thyrocytes from the ES cells, we sought to explore whether these cells could produce thyroid hormones. KI was provided in the medium from day 13, and the supernatant from each medium change after day 13 was saved. These samples were tested for the presence of T3 and T4, but none was detected (data not shown). Assuming that there might be thyroid hormone production below the detection limit of our test, we used electron microscopy to look for signs of secretory activity in day 17 cells. Adult human thyrocytes had MV, RER, Golgi bodies, and secretory vesicles, whereas cells from TSH-negative cultures showed none of these structures. Cells from TSH-positive cultures had abundant MV, RER, and large Golgi bodies, but lacked secretory vesicles; it is conceivable that these cells were producing proteins for secretion which were being stored in the large Golgi bodies, but were not able to secrete them, as evidenced by the lack of secretory vesicles.
The lack of secretory vesicles in cells from TSH-positive cultures may be due to the cultivation conditions being insufficient to effect complete cell development and differentiation (19,20). In common with many other epithelial cell types, the control of proliferation and morphological differentiation of the thyrocyte appears to be dependent upon the integrated actions of a number of growth and tissue-differentiating factors beyond TSH and insulin (9,21 –23).
Another approach to characterize the differentiation of the thyrocytes is to measure their iodide uptake. For technical reasons, we were unable to perform this test. Notably, Arufe et al. measured iodide uptake in CCE mouse ES cells that were differentiated into thyrocytes (2). They found that maintenance of cell cultures in TSH was required for iodide uptake. Because our cells were maintained in TSH-containing cultures, they may also be able to uptake iodide. We recommend further studies to determine if iodide uptake takes place in E14 mouse ES cells stimulated with our conditions.
In summary, we have established a method for differentiating E14 mouse ES cells into thyrocytes. Previous reports have shown that three-dimensional culture is better than adherent culture for thyrocyte growth in vitro (24 –27). Our next study will attempt to use three-dimensional culture to differentiate ES cells into organotypic EBs capable of secreting thyroid hormones.
Author Contribution
Ningyi Jiang: conception and design, administrative support, data analysis and interpretation, and final approval of manuscript.
Yingying Hu: conception and design, data collection and analysis, interpretation of results, and manuscript writing.
Xiongying Liu: conception and design and data collection and analysis.
Yanfeng Wu: administrative support.
Hong Zhang: conception and design.
Guibing Chen: data collection and analysis.
Jiugen Liang: important intellectual contributions.
Xianping Lu: important intellectual contributions.
Sheng Liu: important intellectual contributions.
Footnotes
Acknowledgments
This work was supported by the National Nature Science Foundation of China (grant no. 30370423) and Guangdong Natural Science Foundation (grant no. 04009634). We thank Dr. Xu for generously providing the ES cells.
Disclosure Statement
The authors declare that no competing financial interests exist.