
Abstract
Background:
Poorly differentiated thyroid carcinoma (PDTC) has an unfavorable prognosis. Surgical management is the principal treatment approach. In addition, radioiodine treatment and external beam radiotherapy (EBRT) are given to reduce the risk of local relapse. Despite aggressive therapy, the response to treatment tends to become increasingly poorer over time. The objective of this study was to investigate the induction of apoptosis by EBRT as a function of p53 and bcl-2 protein levels in PDTC. The predictive value of these molecules with respect to treatment efficacy was evaluated.
Materials and Methods:
Two different cell lines of PDTC (FTC-133 and ML-1) were irradiated with a dose of 30 Gy. Apoptotic cells were quantified using terminal deoxynucleotidyltransferase-dUTP nick-end labeling staining without irradiation, 48 and 96 hours after irradiation. The protein levels of p53 and bcl-2 were measured simultaneously using flow cytometry and western blotting. The cell cycle distribution was determined.
Results:
Untreated FTC-133 cells showed a high rate of apoptosis, a high protein level of p53, and a low bcl-2 protein level. Forty-eight hours after irradiation, a slight reduction in apoptotic cells was observed in conjunction with an increase in bcl-2 and p53 protein levels. The slightly reduced fraction of apoptotic cells remained at the same level up to 96 hours after irradiation, whereas the p53 protein level was further downregulated. The cell cycle distribution showed a significant G2/M arrest after 48 hours and recovery 96 hours after irradiation. ML-1 cells did not show any detectable p53 levels and revealed a low rate of apoptosis which significantly increased 48 hours after irradiation. Ninety-six hours after irradiation, a decrease in apoptosis was detectable. The protein level of bcl-2 increased significantly within 48 hours and decreased 96 hours after irradiation. The cell cycle distribution showed a G2/M arrest after 48 hours without a recovery 96 hours after irradiation.
Conclusions:
The p53 and bcl-2 expression profiles and the observed apoptotic rates of FTC-133 and ML-1 under irradiation are consistent with a more aggressive FTC-133 phenotype. Alterations in p53- and bcl-2 protein levels yield predictive information for EBRT efficacy.
Introduction
The accepted approach for the treatment of differentiated thyroid carcinoma is the surgery (near/total thyroidectomy) followed by radioiodine treatment. In poorly differentiated thyroid carcinoma (PDTC), only complete surgical resection can enable a cure. However, a complete surgical resection is not possible in the majority of cases, so that radical surgery is not indicated but airway management is needed at all events. External beam radiotherapy (EBRT) is used for locoregional control and for the palliative treatment of symptoms. Even a positive response at the start of EBRT does not necessarily lead to a good prognosis. Using aggressive therapy protocols, such as hyperfractionated radiation therapy in combination with chemotherapy and surgery, the 3-year survival rate for anaplastic thyroid carcinoma was found to be less than 10% (3).
The difference between anaplastic and PDTC remains unclear due to the lack of consensus regarding criteria and definitions. In 1983, Sakamoto et al. (4) introduced the term “poorly differentiated thyroid carcinoma” based on such histological features such as nonglandular components with a solid, trabecular, and/or scirrhous growth pattern and then others added other histological features such as diffuse sclerosing. Overall these variants show a more aggressive behavior than the types of differentiated thyroid carcinoma, even though certain PDTC do not invariably have a poor prognosis. For example, the solid variant of PTC shows some of the histological features described, but patients tend to be younger, and the overall prognosis is similar to that of a classical PTC (5).
One of the most widely accepted definitions of PDTC was proposed by Burman et al. (6) and it states that “poorly differentiated thyroid carcinoma is a concept proposed to include carcinomas of follicular thyroid epithelium that retain sufficient differentiation to produce scattered small follicular structures and some thyroglobulin, but generally lack the usual morphologic characteristics of papillary and follicular carcinoma.”(7).
The main problem in the course of PDTC therapy is the increasing resistance of tumor cells to therapies such as EBRT. Even though a good response is shown in the beginning, a rapid relapse often follows.
The intended effect of irradiation is the induction of apoptosis in tumor cells (8). Apoptosis, also known as programmed cell death, refers to the disintegration of cells into membrane-bound particles that are eliminated by phagocytosis or by shedding (9). Numerous apoptosis-regulating genes are known to play an important role in the development and progression of cancer. The tumor suppressor protein p53 is an important controller of DNA damage. Whenever this guardian of the genome (10) detects DNA damage, p53 induces a cell cycle arrest, allowing molecular repair. If this repair is not possible, p53 ensures by sensing genotoxic stress that the damaged cell will be destroyed by apoptosis in response to genotoxic stress. A mutated p53 can cause an insufficient elimination of damaged cells so that these can continue to proliferate. In turn, carcinogenesis can occur. A high rate of p53 mutational inactivation is one of the most frequent molecular alterations in all human cancers (11).
Mutations in p53 are almost exclusively detected in poorly differentiated thyroid tumors (12). This suggests that inactivation of p53 leads to dedifferentiation and the development of more aggressive tumor cell growth (13). The lower levels of p53 in well-differentiated tumors indicate that p53 expression may correlate with tumor progression (12).
Another apoptotic pathway in mammalian cells is regulated by cytochrome-c release from mitochondria (14), managed by bcl-2 proteins forming channels that facilitate cytochrome-c transport. Cytochrome-c causes caspase activation, which leads to apoptosis. Members of the bcl-2-family show proapoptotic and antiapoptotic reactions. They were first isolated as a gene involved in B-cell lymphoma and represent a group containing a dozen proteins (15). The expression of bcl-2 as a known inhibitor of apoptosis in normal thyroid epithelium (16) is crucial for the regulation of apoptosis in thyroid carcinomas. Therefore, an overexpression may induce cell proliferation because cells that are already destined for death may undergo further mutations (17). Lu et al. (18) showed that in thyroid carcinomas, bcl-2 expression correlates with tumor cell differentiation. It has also been shown by Lu et al. (18) that bcl-2 inhibits apoptosis activated by wild-type p53, whereas the mutant p53 inhibits apoptosis. Mutations of the bcl-2 gene are little known. Even though the mutation exists, it does not have an effect on proliferation (19).
Despite the fact that irradiation initially reduces the number of carcinoma cells in the treatment of PDTC, the reason why a frequently increased tumor cell resistance occurs during further treatment remains unclear. Our hypothesis is that a mutation of p53 and bcl-2 with complete loss of function over the period of treatment leads to a rapid selection of resistant subclones of tumor cells.
Currently, there are no data regarding apoptosis after radiotherapy of thyroid carcinoma.
The objective of this study was to investigate the induction of apoptosis as a function of p53 and bcl-2 protein levels in PDTC after irradiation. A possible predictive value of these molecules with respect to treatment efficacy was evaluated.
Materials and Methods
Cell cultures
Tissue culture media
Cells were grown in Dulbecco's modified Eagle's medium (PanSystems, Nuremberg, Germany) containing 100 μM sodium pyruvate, 2 mM
ML-1 cell line
The ML-1 cell line was established from a recurrent tumor of a FTC (stage pT4) of a 50-year-old woman (20) and added to the histological type of PDTC. Doubling time was 4 days. The cells were able to take up glucose and secrete thyroglobulin, chondroitin sulfate, fibronectin, thyroxine, and triiodothyronine. They were tumorigenic in nude mice.
The ML-1 cell line is commercially available at the DSMZ (Braunschweig, Germany; Depositor: Schoenberger J,
ML-1 cell line is positive for cytokeratin, cytokeratin-8, and cytokeratin-18 and negative for cytokeratin-7 and cytokeratin-17. Gene array analysis using the Illumina HumanWG-6 v3 Expression BeadChip Kit (Illumina, San Diego, CA) according to the instructions of the manufacturer shows that ML-1 cell line is PAX8 positive and negative for TTF-1 and p53.
FTC-133 cell line
The FTC-133 cell line was derived from a 42-year-old male with FTC (21). The cells possess differentiated thyrocyte function and show thyrocyte response to local active growth factors and thyrotropin. The cell line belongs to the histological type of PDTC. Mutation of p53 and other complex chromosomal changes are detectable (22).
The FTC-133 cell line is commercially available at HPACC (Salisbury, United Kingdom; Depositor: Koehrle J,
As described by Rath et al. (23), FTC-133 cell line expresses thyroglobulin, TFF-1, and PAX8.
Induction of apoptosis by external beam irradiation
The cells were cultured in 10 cm tissue culture plates, and the culture plates were placed on the acceleration treatment couch. A plate of perspex that was 2 cm thick was positioned above and below the tissue culture plates to compensate for the build-up effect. As previously described by Pohl et al. (24), the irradiation was delivered via an anterior portal by a 6 MV linear accelerator (3 Gy/min; Primus, Siemens, Nuernberg, Germany) at room temperature. Dosimetric evaluations were performed to guarantee a homogenous dose distribution. The chosen dose was 30 Gy to score the maximal amount of apoptosis (25).
Evaluation of apoptosis by terminal deoxynucleotidyltransferase-dUTP nick-end labeling
To evaluate the rate of apoptosis, terminal deoxynucleotidyltransferase-dUTP nick-end labeling (TUNEL) testing was performed by flow cytometry using the MEBSTAIN Apoptosis Kit Direct (Medical and Biological Laboratories, Immunosource, Halle-Zoersel, Belgium). TUNEL staining was based on the specific labeling of terminal deoxynucleotidyltransferase (TdT) to 3′-OH ends of fragmented DNA generated by apoptosis (26). ML-1 and FTC-133 cell cultures (control group, 2 and 4 days after irradiation; each culture contains 5 × 106 cells) were fixed with 4% paraformaldehyde at 4°C. Then cells were washed twice with phosphate-buffered saline (PBS) containing 0.2% bovine serum albumin (BSA). For permeabilization, cells were incubated for 30 minutes at −20°C with 70% ethanol and washed twice as previously described. TdT reaction reagent was added to the cell pellet and incubated for 1 hour at 37°C. Cells were washed twice with PBS containing BSA. After resuspension in 700 μL of the buffer, the cells were analyzed by flow cytometry according to the routine protocol as previously described (27). Ten thousand cells were recorded for each sample, and all experiments were conducted in triplicate.
Negative controls (cells incubated in the absence of TdT) and positive controls (cells treated with DNase I [100 μg/mL] for 1 hour at 37°C) were used for voltage and gain adjustments and assignment of histogram regions.
Flow cytometric analysis
Thyroid cancer cells were investigated in the native state (without irradiation), 48 and 96 hours after irradiation. The monoclonal antibodies of bcl-2 (Chemicon, Hofheim, Germany) and p53 (Coulter Immunotech, Marseille, France) were added to 105 ML-1 or FTC-133 cells that had been prepared and fixed in ethanol (70%) as previously described by Grimm et al. (28). After incubation in the dark at room temperature for 60 minutes, they were washed three times with PBS containing 2% FCS and treated with unconjugated antibodies. After incubation for 45 minutes at room temperature with fluorescein isothiocyanate–conjugated anti-mouse-IgG, they were washed using PBS containing 2% FCS. The antibody-labeled cells were analyzed with a Facscan flow cytometer (Becton Dickinson, Heidelberg, Germany) equipped with an argon laser. Cells exerting fluorescence intensities above the upper limit of the negative control distribution were considered positive.
Western blot analysis
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis and immunoblotting were performed according to the routine protocols (28,29). The following antibodies were used for this study: bcl-2 (Upstate, Lake Placid, NY) and p53 (BD Transduction Laboratories, San Jose, CA). Quantification of the blots was carried out by densitometric analysis (Personal Densitometer No. 50301, Molecular Dynamics, Krefeld, Germany). Bicinchoninic acid was used as a standard protein for determination with the bicinchoninic acid system.
Cell cycle distribution
Analysis of the cell cycle distribution of thyroid cancer cells was performed under control conditions (without irradiation), 48 and 96 hours after irradiation. After individual incubation periods, cells were harvested in a subconfluent state using PBS supplemented with 0.02% trypsin/ethylenedia-minetetraacetic acid, washed twice, and fixed by 70% MeOH on ice for at least 1 hour. For univariate DNA staining, fixed cells were washed twice again with PBS supplemented with 0.2% BSA followed by incubation in the same buffer, supplemented with 10 U/mL RNase (20 minutes, 37°C). Subsequently, propidium iodide (PI) was added to a final concentration of 25 g/mL. Samples were incubated for 15 minutes and analyzed by flow cytometry using a commercial (three laser) FACSCantoII flow cytometer (BD Biosciences, Heidelberg, Germany). Cells were kept in the dark during preparation (30).
Statistical evaluation
SPSS 11.5 was used for statistical analysis. All data are expressed as mean ± standard deviation. All parameters were tested for deviation from the Gaussian distribution based on the Kolmogorov–Smirnov test. We compared the cases using the independent sample t-test or the Mann–Whitney test (based on results from the normality test). Differences at the level of p < 0.05 were considered significant.
Results
Flow cytometric analysis of TUNEL-positive cells
The results of the measurement of apoptosis are listed below.
ML-1 cell line
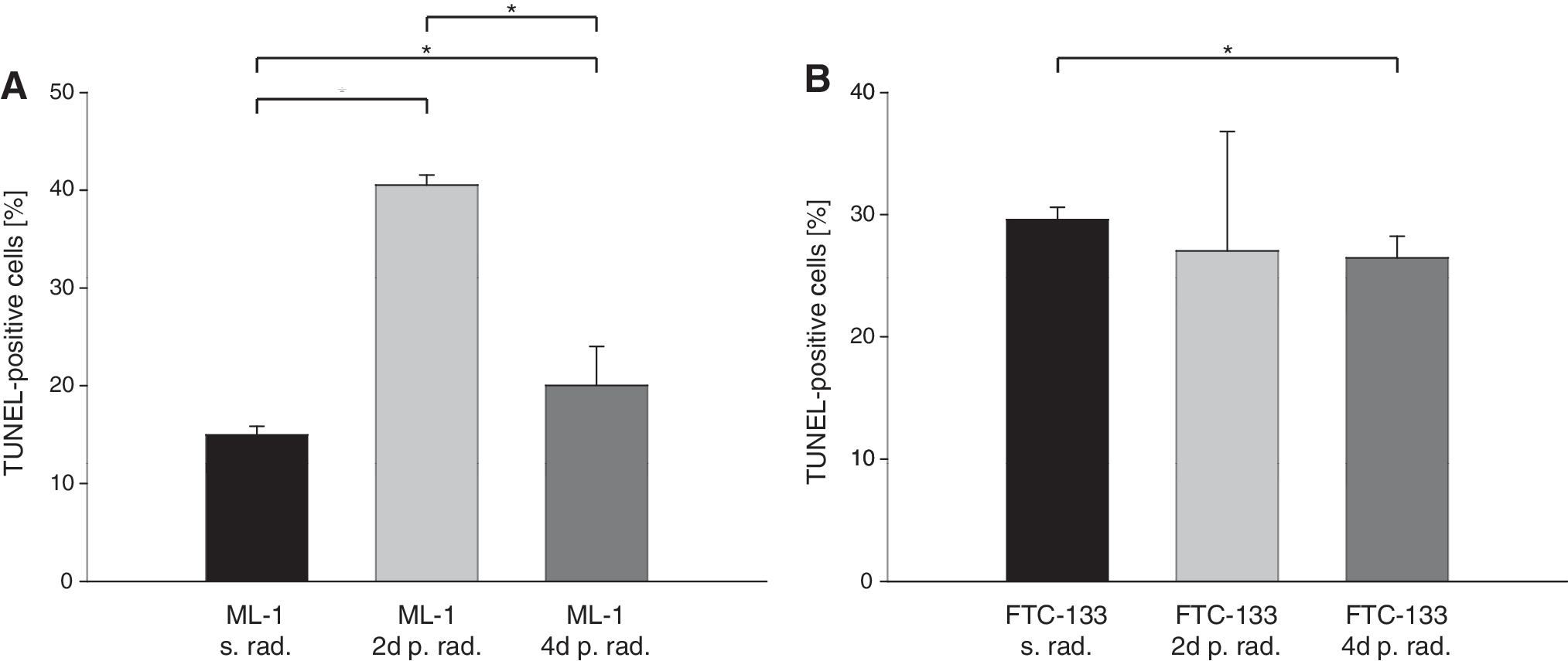
As shown in Figure 1A, irradiation of ML-1 cells induces significant changes in apoptosis. Before irradiation, the fraction of cells that was TUNEL positive was 15.0 ± 0.9%, indicating a low rate of apoptosis. Forty-eight hours after irradiation, the percentage of TUNEL-positive cells increased significantly up to 40.5 ± 1%, showing an induction of apoptosis. Ninety-six hours after irradiation, a significant decline was detectable with only 20.0 ± 4.0% of TUNEL-positive cells.
(
FTC-133 cell line
Compared with ML-1 cells, FTC-133 cells demonstrated contrasting behavior as shown in Figure 1B. Control cultures showed 29.6 ± 1.0% TUNEL-positive cells, indicating a high rate of apoptosis initially. Forty-eight hours after irradiation, the percentages of TUNEL-positive cells declined significantly to 27.0 ± 9.8% and remained at 26.5 ± 1.8% 96 hours after irradiation as measured by flow cytometry.
Detection of bcl-2 measured by flow cytometry and western blot analysis
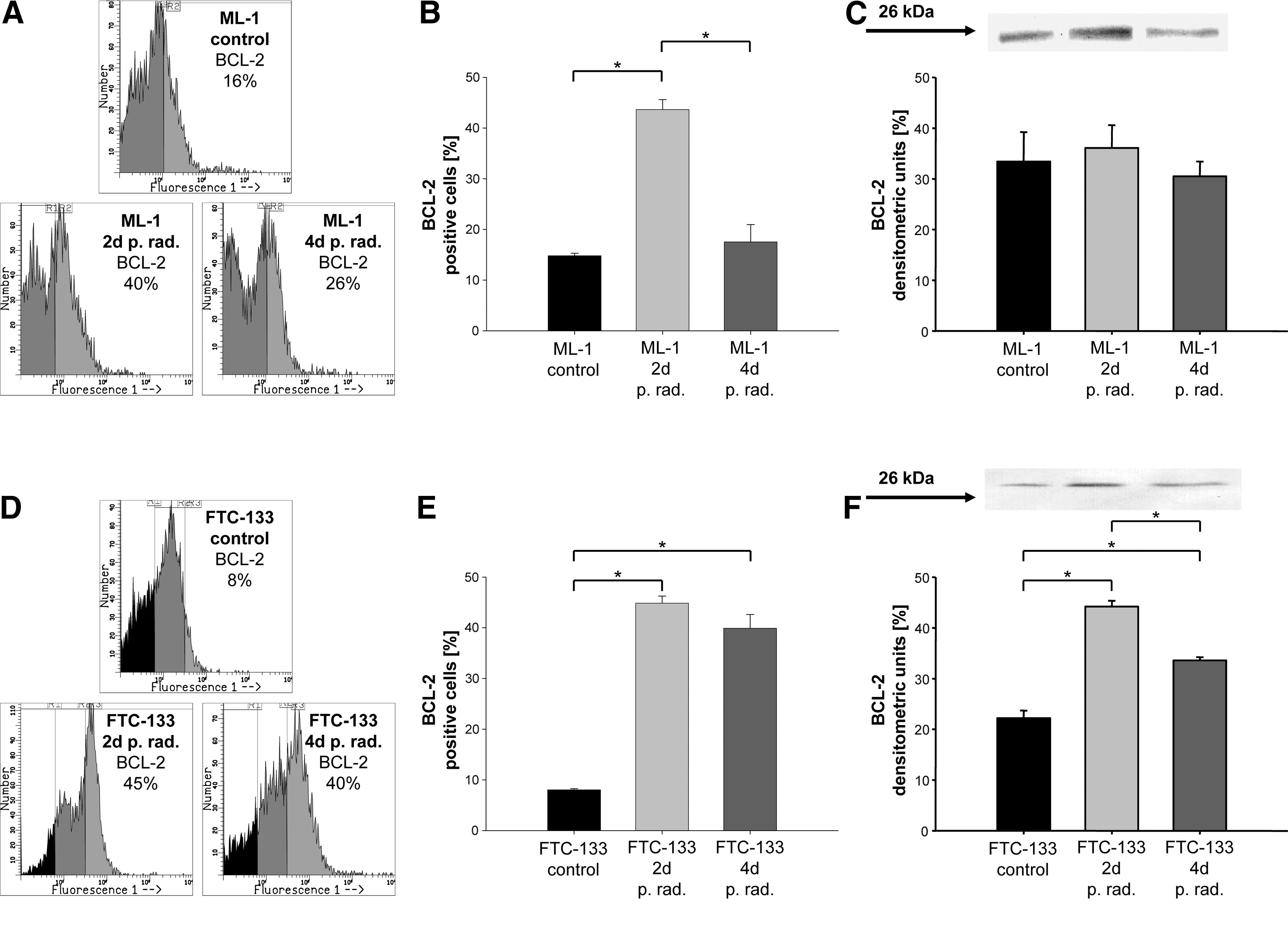
ML-1 cells showed a low percentage of bcl-2-positive cells before irradiation. Forty-eight hours after irradiation, we detected an increase of up to 40% of cells; 96 hours after irradiation, bcl-2-positive cells decreased to 26%. The concentration of bcl-2 protein measured by western blot analysis confirmed the flow cytometric data (Fig. 2A–C).
(
Only 8% of bcl-2-positive cells could be found in control FTC-133 cells. Forty-eight hours after irradiation, a significant increase of up to 45% was detectable that then decreased to 40% bcl-2-positive cells 96 hours after irradiation. The western blot analysis confirmed these data (Fig. 2D–F). No significant changes within the control groups of ML-1 and FTC-133 cells after 2 and 4 days were detectable.
Detection of p53 measured by flow cytometry and western blot analysis
Figure 3A and B shows 63% of p53-positive cells in the control group as measured by flow cytometry. Forty-eight hours after irradiation, a slight increase up to 67% was detectable. Ninety-six hours after irradiation, FTC-133 cells showed a significant decrease to 57% of p53-positive cells. The western blot analysis shown in Figure 3C confirmed the upregulation of the p53 level 48 hours after irradiation and downregulation 96 hours after irradiation. Further, no significant changes within the control groups of FTC-133 cells were detectable after 2 and 4 days.

(
ML-1 showed no expression of p53 as revealed by flow cytometry and western blot analysis.
Cell cycle distribution
Inhibition of cell cycle progression (Fig. 4A, B) 2 days after irradiation has been proven by a decrease in the G1-phase fraction in both cell lines (ML-1: control 80%; 48 hours 42%/FTC-133: control 56%; 48 hours 19%) and an increase in G2/M-fraction lines (ML-1: control 13%; 48 hours 45%/FTC-133: control 30%; 48 hours 74%).

Cell cycle distribution is shown without irradiation and 48 hours as well as 96 hours after irradiation of ML-1 (
In contrast to the ML-1 cells, which show no change in cell cycle distribution (continuing G2/M arrest) between 2 and 4 days after irradiation (G1: 48 hours 42%; 96 hours 42%/G2/M: 48 hours 45%; 96 hours 46%), the FTC-133 cells show a recovery of G2/M arrest 4 days after irradiation (G1: 48 hours 19%; 96 hours 47%/G2/M: 48 hours 74%; 96 hours 45%). All the changes that were identified in cell cycle progression are significant (Fig. 4A, B).
Discussion
The main problem in treating a PDTC is that resistance develops against all kinds of therapy, especially irradiation. As shown by Gamble et al. (31), even a nontumorigenic thyroid cell line can transform into a tumorigenic cell line in consequence to in vitro exposure to radiation. Interestingly, 62% of the developed cell lines had p53 mutations. It is already known that a mutation in the p53 tumor suppressor gene is associated with dedifferentiation in thyroid cancer (32) and that other apoptotic markers like bcl-2 also play a role. If there is a correlation between the degree of expression of apoptotic markers such as p53 and bcl-2 and the degree of differentiation of the tumor, the regulation of these apoptotic markers by a thyroid carcinoma cell line in vitro could give an indication as to whether a chosen therapy such as irradiation will work.
In this study, we attempted to determine the regulation of apoptotic markers p53 and bcl-2 in the two different PDTC cell lines—ML-1 and FTC-133—with and without radiation exposure to see if such therapy leads to radioresistance and whether response to therapy can be predicted on the basis of these apoptotic markers.
ML-1 cells are able to take up glucose and secrete thyroglobulin, chondroitin sulfate, fibronectin, thyroxine, and triiodothyronine. They are tumorigenic in nude mice, and protein levels of p53 are not detectable.
FTC-133 cells retain their differentiated thyrocyte function and show thyrocyte response to local active growth factors and thyrotropin. Mutation of p53 and other complex chromosomal changes are detectable (33). Even if two similar FTC cell lines were used, the ML-1 cell line is slightly highly differentiated (34). Only FTC-133 shows detectable levels of p53. Because the latter is known to be the more aggressive neoplasm of the two cell lines, the regulation of p53 under radiation stress is of special interest.
ML-1 cells showed a low percentage of bcl-2-positive cells before irradiation as well as a low percentage of TUNEL-positive cells, indicating a low amount of apoptosis and a majority of living cells. Most of the cells were in the G1 fraction. Apoptosis is a physiological process regulating the cell population in the thyroid gland (35). One of the functions of bcl-2 is to protect against cell death. In thyroid carcinomas, bcl-2 expression correlates with tumor cell differentiation (36). Forty-eight hours after irradiation, an increase of up to 40% of bcl-2-positive cells was detectable. This was accompanied by enhanced apoptosis and an inhibition of cell cycle progression. The upregulation of bcl-2 could be interpreted as an attempt by the cells to block apoptosis. After 96 hours, bcl-2-positive cells decreased to 26%, whereas apoptosis was still high, and no recovery of cell cycle arrest was observed.
The significant increase in bcl-2-positive cells (2 days after irradiation) as a turnover reaction could be defined as a transitional stage that is independent of cell line origin. Four days after irradiation, a significant decrease in bcl-2 levels could be measured. Loss of the apoptosis-inhibiting function of bcl-2 leads to induction of apoptosis.
In the FTC-133 cell line, a high rate of spontaneous apoptosis together with a low inhibitory impact of the basely expressed bcl-2 can be observed in untreated cells. With regard to the high protein level of p53 without irradiation, it could be presumed that p53 is performing its function as the guardian of the genome (37) by allowing repair, or in cases in which cellular repair is not possible, it ensures that the damaged cell will be destroyed by apoptosis in response to genotoxic stress. However, Fagin et al. (13) described a high prevalence of mutations in the p53 gene in PDTC and suggested that inactivation of p53 leads to dedifferentiation manifested by a more aggressive behavior. Forty-eight hours after irradiation a slight increase in p53-positive cells was detectable. Ninety-six hours after irradiation, FTC-133 cells showed a significant but slight decrease of p53-positive cells. Hence, a turnover reaction is also detected in FTC-133. This is reflected by a high level of p53 and bcl-2 and an inhibition of cell cycle progression by irradiation. This has been substantiated by a decrease in the G1-phase fraction 48 hours after irradiation. On day 4 after irradiation, there was a decline in p53 levels, and an increase in bcl-2-positive cells occurred compared with the nonirradiated control cells. These alterations implicated by the increase of the antiapoptotic bcl-2 were accompanied by a reduced rate of apoptosis. However, a reduced level of p53 is no longer capable of fulfilling its important function as a controller of DNA damage. Consequently, no appropriate repair or induction of apoptosis could be initiated as indicated by the recovery of G2/M arrest 96 hours after irradiation.
In summary, an appropriate constant and sufficient therapeutic success could be shown only for the irradiated ML-1 cells. The increase in bcl-2 levels correlates with an increase in apoptotic cells. The FTC-133 thyroid carcinoma cell line, which is known to be more aggressive than ML-1 (34), although both cell lines originate from FTC, shows contrasting behavior. After irradiation, the fraction of apoptotic cells declined, whereas the fraction of living tumor cells increased. A correlated increase in bcl-2-positive cells and a decrease in the number of p53-positive cells was also seen. One possible reason for this could be the selection of radioresistant clonogenic cells by irradiation. It is also possible that mutations in the p53 pathway are induced by therapeutic interventions such as EBRT. As described by Gamble et al. (31), this might be responsible for the development of new subclones. Whether such p53 mutations also lead to anaplastic transformation is still unclear, although this seems very likely because several in vitro studies have indicated that wild-type p53 can be reintroduced into PDTC cell lines with a nonfunctional p53. The result was a slower growth and a redifferentiation of cells with a higher sensitivity to chemotherapy and radiotherapy (38,39).
The demonstrated p53 and bcl-2 expression profiles and the observed apoptotic rates of FTC-133 and ML-1 PDTC 2 after irradiation also proven by the results of cell cycle distribution are in agreement with a more aggressive FTC-133 phenotype. Alterations in p53 and bcl-2 protein levels in connection with individual apoptotic rates under irradiation stress are predictive for EBRT efficacy. The alterations could be regarded as an early marker for response to treatment or rather of a buildup of resistance to irradiation during the course of treatment. Further studies will be required to establish whether irradiation leads to a selection of already resistant subclones or induces resistance.
Footnotes
Acknowledgments
We thank Elisabeth Schmidt-Bruecken from the Department of Obstetrics and Gynecology, University of Regensburg, Germany, and Henriette Dam from the Department of Nuclear Medicine, University of Regensburg, Germany for their excellent technical support.
Disclosure Statement
The authors declare that no competing financial interests exist.