
Abstract
Background:
Rapid deconjugation of administered thyroxine glucuronide (T4G) to thyroxine (T4) has been observed in human subjects. The goal of this study was to characterize, in the rat, the location and kinetics of this deconjugation process and to identify whether early conjugation of T4 to T4G occurs in the liver.
Methods:
Normal male rats received intraportal radiolabeled T4 and T4G. Tissues were assayed 1 to 15 minutes after the injection.
Results:
After T4G injection, assay of the kidneys, and to a lesser extent assay of the liver and plasma, showed presence of considerable T4, the result of deconjugation. In the kidneys, 79 ± 12% (mean ± SD%) of total (T4 + T4G) was in T4 form. In the liver and plasma, T4 was 10 ± 9% and 6 ± 5%, respectively, of the total (T4 + T4G). No significant conjugation of T4 occurred in any of the tissues assayed during this short time period. Secretion in the bile of administered T4G as T4G increased with time.
Conclusions:
These data, together with the previous observations in human subjects, suggest that T4G, with its greater tissue distribution, may serve as a medium for transfer of T4 into tissues not subject to direct transfer of T4 from the circulation. Hence, the true volume of distribution of T4 is postulated to be greater than that measured by tracer studies of T4 distribution.
Introduction
The bacteria in the lower intestine contain glucuronidases that hydrolyze the glucuronides, so that no glucuronides are present in the feces except when the bowel has been sterilized (1). Glucuronidases have been demonstrated in other tissues, but deconjugation of T4G has usually not been considered to be a major process elsewhere than in the lumen of the lower intestine.
Studies in human subjects given radiolabeled T4G orally or intravenously (2) showed that there is a more widely distributed deconjugation process, and that deconjugation occurs rapidly after T4G administration. These studies also showed deconjugation to be reversible, so that, over time, the T4/(T4 + T4G) ratio became constant. The results were interpreted to indicate that both conjugation of thyroid hormone with glucuronic acid and subsequent deconjugation occurred widely and rapidly. Because the volume of distribution of labeled T4G in these human subjects was found to be much greater than that of labeled T4, it was postulated that transition to and from the glucuronide form might be a mechanism for intracellular transfer of T4.
The goals of the present study in rats were to identify the timing and location of deconjugation of T4G and to ascertain whether glucuronide conjugation of T4 by the liver occurred during the early minutes after arrival in the liver. Not all planned experiments were completed because of laboratory closure. Presented here are the results for which conclusions are statistically justified.
Methods
131I-labeled T4 was prepared in our laboratory using the chloramine T iodination method and purified by high-performance liquid chromatography (HPLC) as previously reported (3). 125I-labeled T4G was prepared from commercially supplied 125I T4 incubated with commercially available uridine diphosphate-glucuronosyltransferase and purified by HPLC, also as previously reported (4). HPLC of aliquots of the injected material added to nonradioactive rat serum and extracted together with the tissue samples showed 83.2 ±5.5% (mean ± SD%) of the administered 125I to be in T4G form and 93.4 ± 4.6% of the administered 131I to be in T4 form. The primary contaminant was iodide (10.3 ± 3.7% of 125I and 2.4 ± 3.1% of 131I). The 125I-T4G contained 2.5 ± 3.0% T4 and the 131I-T4 contained 0.4 ± 0.2% T4G.
Twenty-one adult male Sprague Dawley rats (mean weight 387 g) were anesthetized with intramuscular ketamine and xylazine. Laparotomy was performed, and the portal vein was cannulated. Approximately 5 μC 131I-labeled T4 and 5 μC 125I-labeled T4G were injected into the portal vein cannula at time zero. The purpose of the intraportal route for injection was to detect any early glucuronide conjugation of T4 by the liver.
At the experimental time planned for a given rat (between 1 and 15 minutes after injection), a blood sample was taken from the heart, and the rat was killed by removal of the heart. The liver, both kidneys, and the bile in the gall bladder were removed for study. Plasma was separated from the blood sample by centrifugation. Plasma volume was estimated from the rat's weight at .042 mL/g.
Aliquots of the sampled tissues were extracted in acid ethanol and the supernatant lyophilized. The dried supernatant was reconstituted with 40:60 acetonitrile:water and assayed on HPLC (3). Results were corrected for separately measured extraction efficiency for T4 and T4G and were expressed as % administered dose in the liver, in the two kidneys, in the cumulated bile, or in the plasma volume.
Results
Success of injecting into portal circulation
Despite the fact that the tracers were injected into the cannulated portal vein, mean T4 in the liver after T4 injection was only 47%, and mean liver T4G after T4G was only 27%. The plasma values fitted to single exponential decay curves with zero values of 50.7% for T4 and 67.1% for T4G. This indicates that the injection was, in effect, split between direct delivery to the liver via the portal vein and delivery into the general circulation. Appearance of high levels of T4G in the bile of four of the rats suggests that some of the injection may have been into the biliary collecting system.
Bile T4 and T4G
After T4 administration, neither T4 nor T4G measured in the bile accounted for as much as 1% of the administered dose. The amount measured in the bile did not correlate with time.
On the other hand, after T4G administration, both T4 and T4G biliary excretion correlated with time, although there was little T4 present (under 1%, except for one sample at 1.2%). T4G appeared promptly in the bile. In four of the rats, over 20% of the T4G dose appeared in the bile as T4G, despite the short study period (Fig. 1).
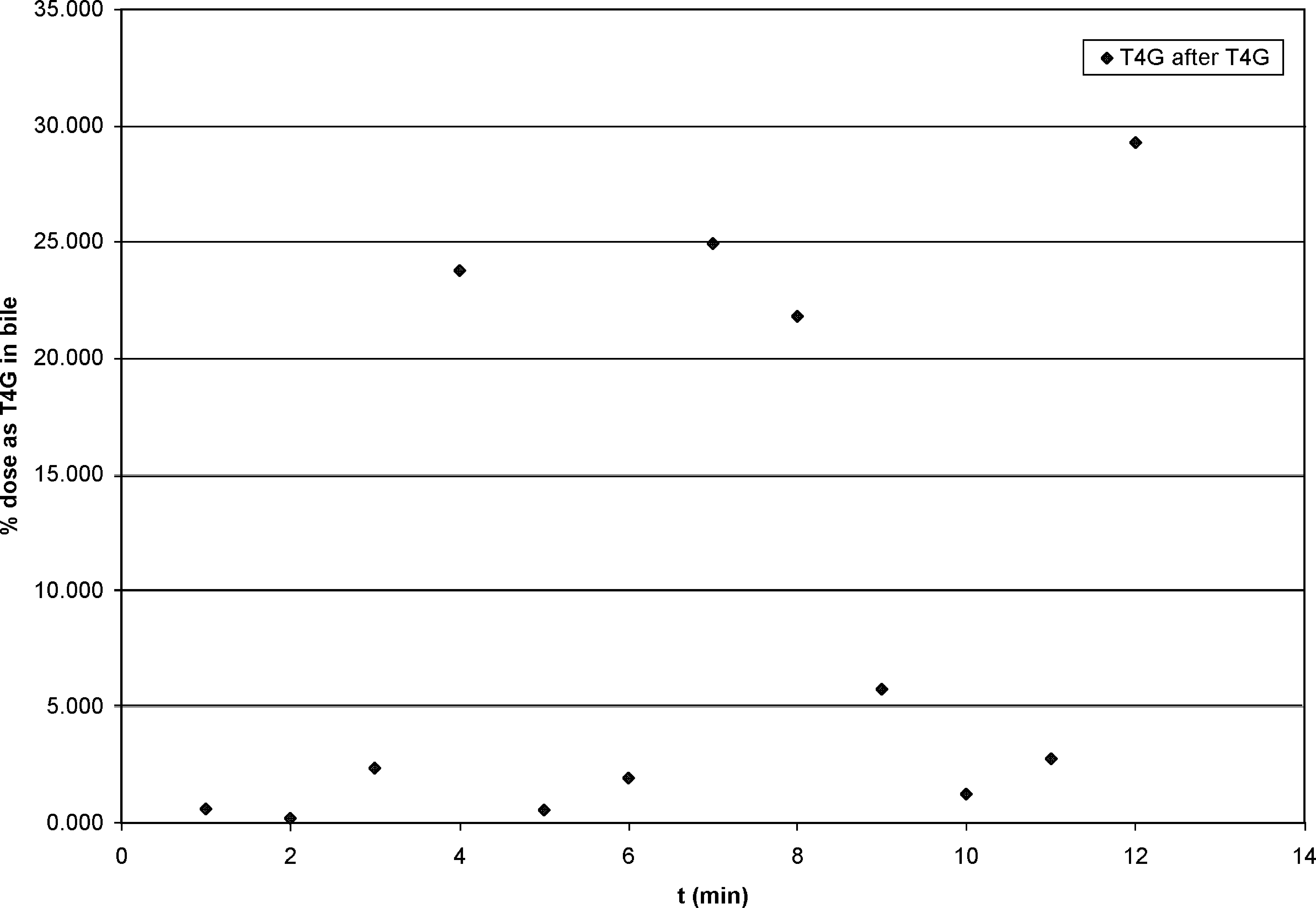
Cumulated biliary thyroxine glucuronide (T4G) after portal injection of T4G. Despite wide scatter, the correlation of excretion with time is 0.57.
Liver, kidney, and plasma T4 and T4G activity with time after portal injection
As mentioned above, the temporal patterns of both T4 and T4G in plasma fitted to declining exponential functions. The liver T4 concentration after T4 portal injection increased through the 15-minute experimental period, while the liver's T4G concentration after T4G rose to a maximum at about 3 minutes and then declined. The kidney data showed no apparent temporal patterns.
Conjugation of T4
During the 15-minute experimental timeframe, no significant evidence of conjugation of T4 to T4G was seen in the liver, the kidneys, or the plasma.
Deconjugation of T4G
On the other hand, deconjugation of T4G to T4 occurred consistently early after portal injection. Of the three organs assayed (plasma, liver, and kidneys), deconjugation was most prominent in the kidneys, with 79 ± 12% (mean ± SD%) of the total (T4 + T4G) measured in the kidney tissue present in T4 form. The corresponding percentages for the liver and plasma were 10 ± 9% and 6 ± 5%, respectively. Figure 2 shows the individual T4/(T4 + T4G) ratios after T4G injection in the kidneys, compared with the plasma and liver. This ratio in the plasma increased with time (correlation coefficient 0.74), indicating that there is return of T4 into the plasma after T4G administration. The kidney was by far the most important site of deconjugation.
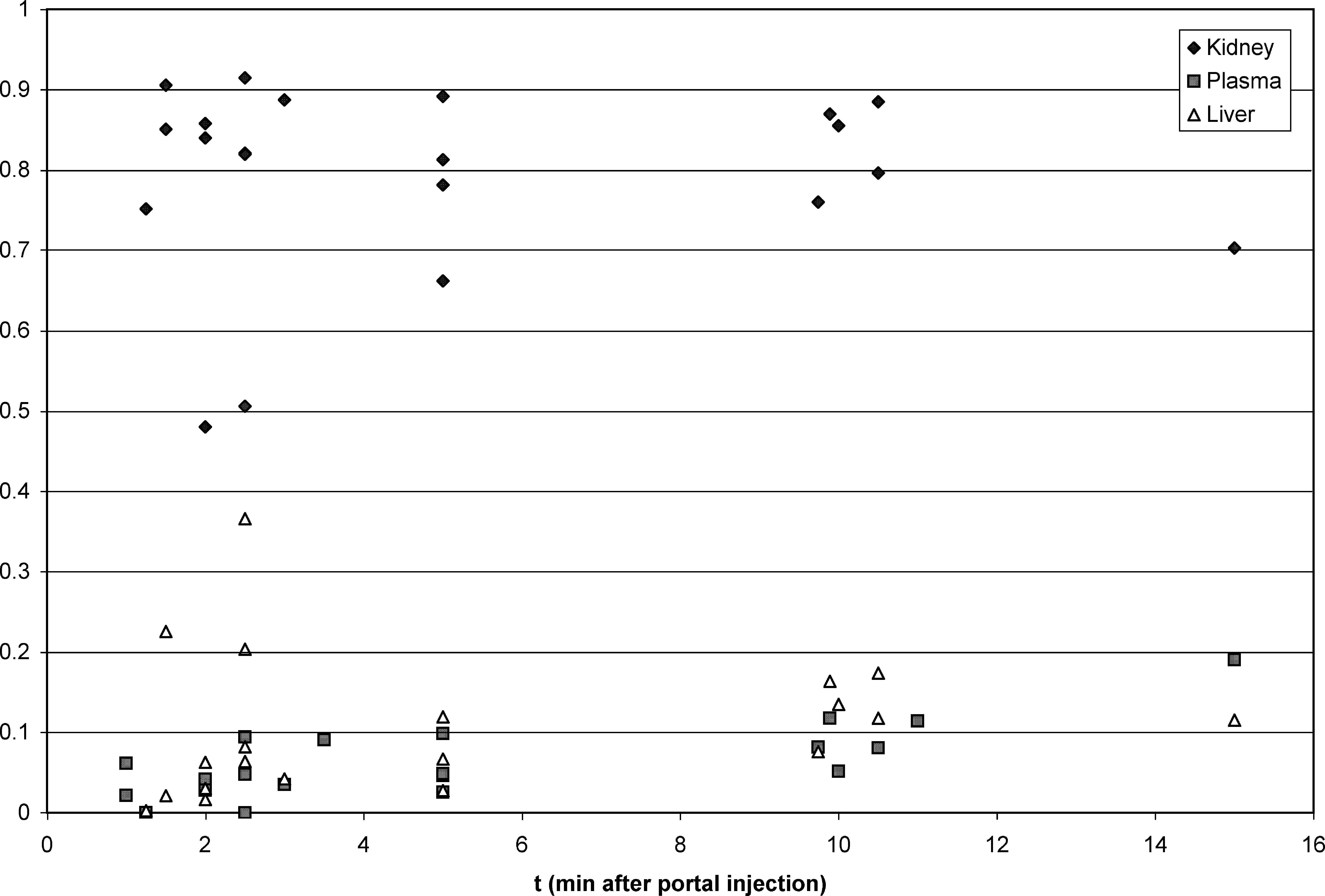
Ratio of thyroxine (T4) content to total (T4 + T4G) content in the kidney compared to that in the liver and plasma. By t-test, the difference between kidney and liver and the difference between kidney and plasma are both significant at the <0.0001% level. Liver values exceed plasma values at only the 4% level of significance.
The T4/(T4 + T4G) ratios after T4G injection in the four animals with unusually high biliary T4G excretion after T4G did not differ significantly from those of the other animals. Data for the kidney are shown in Figure 3.
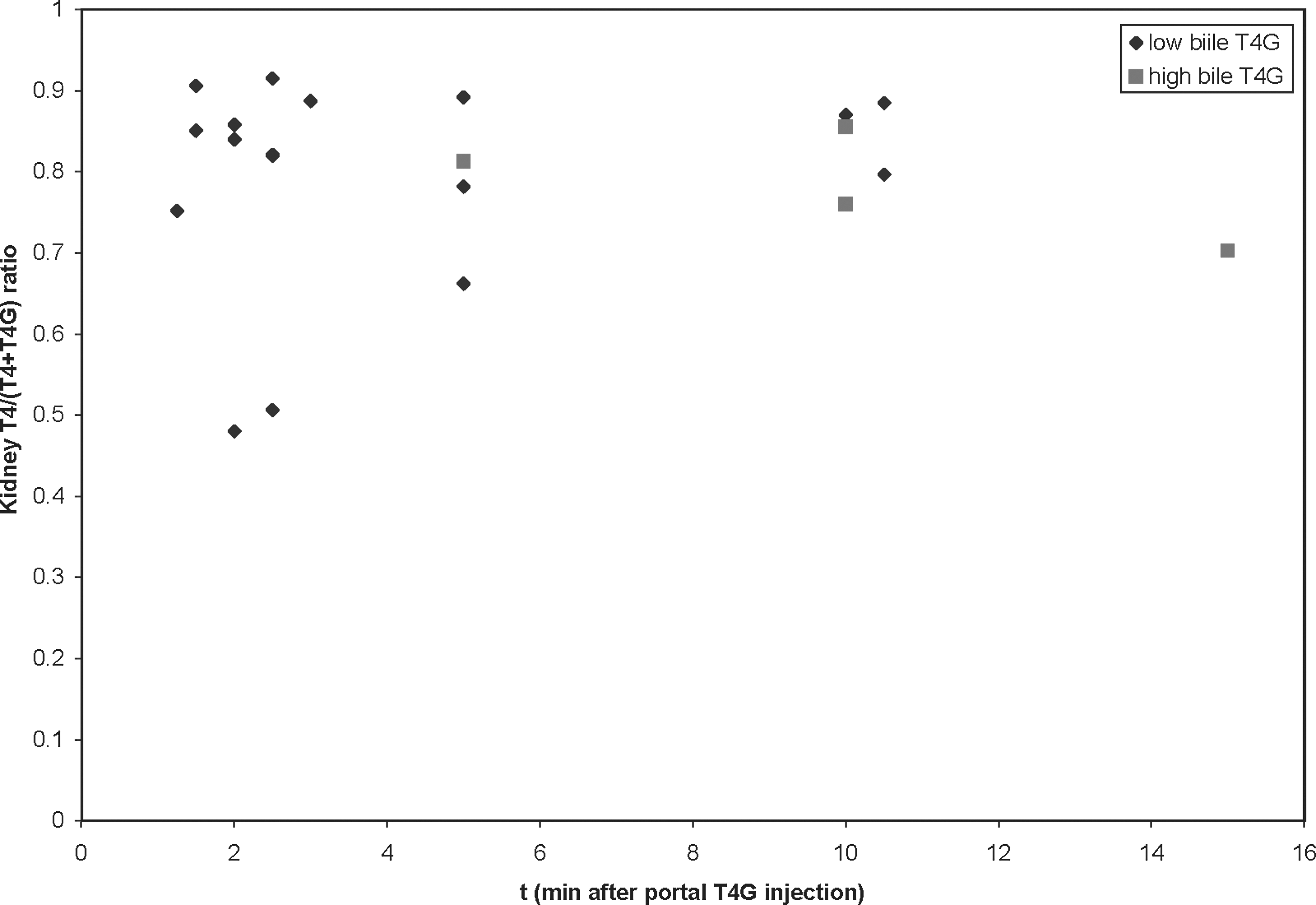
Ratio of T4 content to total (T4 + T4G) content in the kidney, identifying the four rats with high biliary T4G excretion. Their deconjugation rate did not differ significantly from that of the other animals.
Discussion
Despite their paucity, these data clearly show deconjugation of T4G by the rat kidney to occur promptly after T4G administration. Because of local constraints, it was not possible to explore whether some characteristic other than presence of ample beta glucuronidase in the kidney accounts for this phenomenon or to assess urinary radioactivity. However, the author believes that further work by others will confirm that the kidney is an active site of T4G deconjugation. Other sites of deconjugation undoubtedly occur, especially the intestine, as others have demonstrated (1). However, much of the T4 resulting from T4G deconjugation in the colon would be expected to be excreted in the feces, as the colon is not an active site of T4 absorption (3). This is believed to be the first demonstration of release of T4 into the rat's general circulation after T4G administration.
This finding confirms the inference drawn from human studies (2) that deconjugation of T4G is prominent, and this study identifies the kidney as a major site of deconjugation. The study in human subjects also showed tracer T4G to have a greater volume of distribution than does tracer T4. Tissue distribution in T4G form followed by deconjugation to T4 could well be a mechanism that increases the effective volume of distribution of T4 to a value greater than that established from study of the kinetics of tracer intravenous T4 alone.
Estimates of human total body iodine derived from long-term tracer studies (5) predict total human iodine content to be about 15 mg, of which about 4.6 mg is extra thyroidal. This 4.6 mg of extrathyroidal iodine is much more than the 1.1 mg of extrathyroidal iodine estimated from the combined measurements of the plasma-exchangeable T4, T3, reverse T3 (rT3), tetrac, and iodide pools. The nature of this extra iodine has not been established. Results of the present study suggest that tissue T4G and the T4 transported to tissues as T4G, which is subsequently deconjugated, contribute to this previously unmeasured iodine pool.
Footnotes
Acknowledgments
The author thanks Lichi (Gigi) Hsu for expert technical assistance. This work was supported by DVA Medical Research (Program 821) funds.
Disclosure Statement
The author declares that no competing financial interests exist.