
Abstract
Background:
Maternal thyroid hormones (THs), especially thyroxine (T4), are crucial to early brain development in the mammalian embryo. Epidemiological studies and case reports have shown that maternal subclinical hypothyroidism may result in significant negative effects on pregnancy and neurodevelopment of the fetus. To understand the mechanism responsible for these neurological alterations, we induced maternal subclinical hypothyroidism in pregnant rats. Behavior and several genes that are under the control of THs were evaluated in the offspring of TH-deficient rats.
Methods:
A total of 60 female rats were divided into three groups: (i) maternal subclinical hypothyroidism (total thyroidectomy with T4 infusion), (ii) maternal hypothyroidism (total thyroidectomy without T4 infusion), and (iii) control (sham operated). All rats were mated 10 days after the start of infusion. The infusion continued until 10 days postpartum. Pups were sacrificed at postnatal day 3 (PND 3), PND 7, and PND 21. The hippocampus was collected and tested for brain-derived neurotrophic factor (BDNF) and Rap1 protein expression by Western blotting and for BDNF and neural cell adhesion molecule mRNA expression by real-time polymerase chain reaction. On PND 41–PND 49, rat pups explored the Morris water maze. Time spent in the quadrant previously containing the platform was recorded.
Results:
This study found decreases in BDNF mRNA expression (on PND 3) and protein level (on PND 3 and PND 7) in hippocampi of pups from subclinical hypothyroidism dams (p < 0.05). Rap1 protein expression was higher in maternal subclinical hypothyroidism offspring than in control offspring at PND 7 and PND 21. No change was observed in neural cell adhesion molecule mRNA expression in the maternal subclinical hypothyroidism offspring. In addition, results from the Morris water maze revealed that pups from the subclinical hypothyroidism dams showed deficits in long-term memory, spending less time in the platform quadrant (p < 0.05) during testing. There was a trend toward a deficit in short-term memory (p > 0.05) in this group as well.
Conclusions:
The long-term memory deficits of pups born to maternal subclinical hypothyroidism dams likely related with decreasing in BDNF mRNA expression and protein level as well as increasing in Rap1 protein expression in hippocampi.
Introduction
There is growing concern that TH insufficiency, even subtle insufficiency, in circulating maternal THs in the first and second trimester of pregnancy may adversely affect neurodevelopment in offspring, resulting in irreversible cognitive deficits (3 –5). Pregnant women with undetected subclinical hypothyroidism or even serum-free T4 concentration at a low but normal range (10th percentile) had children with adverse neurodevelopment and significantly lower development quotients when assessed in infancy (2). These findings strongly suggest that maternal THs play important roles in brain development before birth.
While previous studies have shown a clear correlation between maternal thyroid status and neurological outcome of offspring, the molecular basis underlying these events has not been thoroughly investigated. The physiological effects of TH are the end result of the regulation of a series of target genes. Brain-derived neurotrophic factor (BDNF), neural cell adhesion molecule (NCAM), and Rap1 are known to be directly regulated by TH. These molecules play essential roles during the critical period of fetal brain development (6,7). The aims of this study were to investigate the effects of maternal subclinical hypothyroidism during pregnancy on gene and protein expression under the control of THs of rat offspring. In addition, the effect of maternal subclinical hypothyroidism on memory in offspring was also examined.
Materials and Methods
Animals
Specific pathogen-free nulliparous female Wistar rats (n = 60), weighing 200–220 g, were purchased from the Animal Research Unit, Vitalriver Corporation, and housed in a climate-controlled specific-pathogen-free laboratory (temperature 24°C ± 1°C and relative humidity 55% ± 5%) with a 12-hour light/dark photoperiod. All animals were permitted free access to normal rat chow and tap water. All experimental procedures were approved by Animal Care and Use Committee at China Medical University, which complies with the National Institute of Health Guide for the Care and Use of Laboratory Animals.
Experimental design
All female rats were mated with normal males. Day of conception was confirmed by microscopic observation of spermatozoa (from vaginal smears) and was designated as embryonic day 0. The rats were housed in individual cages until delivery. Levels of total T4 and thyrotropin (TSH) in serum of the dams were monitored every 3 days during pregnancy to confirm the efficacy of treatment. Maternal weight was measured daily. The day of birth was designated as postnatal day 0 (PND 0). On PND 1, the number of pups in each litter was recorded. To minimize disturbance, pups were weighed on PND 3, PND 7, and PND 21. The pups (n = 12) removed from the litters on different ages were sacrificed and their brains were removed quickly. The hippocampi were dissected out, frozen in liquid nitrogen, and stored at −70°C until the day of assay. The remaining pups from each litter were weaned at PND 21. After weaning, pups were separated into male and female cages. Behavioral testing commenced at PND 40. The project timeline is described in Figure 1.

Schematic of experimental timeline. E15, embryonic day 15; MWM, Morris water maze; PND, postnatal day; T4, thyroxine.
Animal models of maternal subclinical hypothyroidism
Dams were divided into three groups: subclinical hypothyroidism (Sub, n = 20), hypothyroidism (Hypo, n = 20), and control (n = 20).
Maternal subclinical hypothyroidism was induced by the surgical removal of the thyroid gland (total thyroidectomy) under 10% chloral hydrate sedation (3 mL/kg body weight [BW], i.p.) after a 7-day adaptation period, as previously described (8). Rats were provided with 0.1% (w/v) calcium lactate in the drinking water after surgery and maintained on normal rat chow. Four weeks after surgery, as serum T4 concentrations fell below the detectable level (<1.0 μg/dL) and BW was around 220 g, rats were implanted with subdermal (dorsal) osmotic minipumps (ALZET®; models 2001 or 2002, delivering 1.0 or 0.5 μL/h, respectively; Alza Corporation). The osmotic minipumps delivered a constant rate of T4 (Sigma Chemical Co.) at a dose of 1.0 μg/100 g BW per day. The rats were mated 10 days after pump implantation. The pumping rate was increased to 1.05 μg per 100 g BW per day starting on the 15th day postconception and remained as such through the 10th day postpartum. The concentration for T4 replacement was selected based on previous studies by Escobar et al. (8) and our pilot study. These dosages resulted in elevated serum TSH level and normal total T4 level (maternal subclinical hypothyroidism rats).
Maternal hypothyroidism was induced by total thyroidectomy without T4 treatment but rats were implanted with osmotic minipumps containing physical sodium chloride solution. The control group underwent sham surgery (surgery as described above, but without removal of the thyroid gland) and was implanted with osmotic minipumps containing physical sodium chloride solution.
Hormone measurements
Total T4 was measured by immunochemiluminometric assay according to the manufacturer's instructions (Diagnostic Products Corporation), with inter- and intraassay coefficients of variation of 3.58%–6.67% and 1.26%–3.20%, respectively. Plasma TSH concentrations were determined by a chemiluminescent immunoassay (IMMULITE; Diagnostic Products Corporation). The inter- and intraassay coefficients of variation for TSH were 1.57%–4.93% and 1.23%–3.38%. Based on >95% specific binding, the limit of detection for total T4 was 1.0 μg/dL. For total T4, any samples below the level of detection were recorded at 1.0 μg/dL for statistical purposes. TH data for dams are summarized in Table 1.
Values are expressed as mean ± SEM.
p < 0.05, versus same-day Sham control.
p < 0.01, versus Hypo group.
The sensitivity of the chemiluminescence immunoassay for total T4 was 1.0 μg/dL.
E10, embryonic day 10; E13, embryonic day 13; E17, embryonic day 17; PND, postnatal day; SEM, standard error of the mean; TSH, thyrotropin; TT4, total thyroxine.
Morris water maze
The Morris water maze (MWM) is used to test the spatial learning ability and memory of rodents. A video camera mounted directly above the maze was linked to a computer with video-tracking software (Jiliang Ltd.) to automatically record the escape latency (time taken to find the platform on each trial), path length (distance traveled by the animal), and swimming speed of all rats.
On PND 40, one male and one female from each litter were randomly selected from the three groups (n = 12 litters/group), to start MWM training. All behavioral training was carried out as described previously (9). Briefly, on PND 40–PND 44 rats completed four trials per day to locate a submerged platform with a maximum swim time of 60 seconds. Animals were tested for short-term memory 30 minutes after the last trial on P44. During testing, the rat was placed into the pool as in training trials, except that the hidden platform was removed from the pool. Memory consolidation was indicated by the length of time spent in the target quadrant (quadrant previously containing the platform). Training for long-term memory extended from PND 45 to PND 48, and followed the same methods as PND 40–PND 44, with each animal completing four trials day with a maximum swim time of 60 seconds. The platform was removed and animals were tested for long-term memory 24 hours after the last trial. As with short-term memory, time spent in the quadrant previously containing the platform was measured.
Real-time polymerase chain reaction
Total RNA was isolated from hippocampi of neonatal rat pups at three ages (PND 3, PND 7, and PND 21) using TRIzol Reagent (Invitrogen) according to the manufacturer's instructions. Total RNA (1 mg) was reversely transcribed with random primers using a reverse transcription reagent kit (Promega) following the manufacturer's protocol. Complementary DNA was prepared and stored at −80°C for subsequent fluorescent real-time quantitative polymerase chain reaction (PCR). All amplification reactions were performed in triplicate on the ABI PRISM 7500 sequence detection system (Applied Biosystems). The primer sequences for BDNF were 5′-ATCCACTGAGCAAAGCCGAAC-3′ (forward) and 5′-CAGCCTTCATGCAACCGAAGTA-3′ (reverse). The primer sequences for NCAM were 5′-ACAGCGGTGAACCGTATTGGA-3′ (forward) and 5′-AATTGTACCTGTGCTGTGCTGGAG-3′ (reverse). For each sample, 200 ng of cDNA template was added to 12.5 μL of QuantiTect SyBR green master mix (TaKaRa) and 0.5 μL of primers (final concentration, 10 μM each) to a total volume of 25 μL in RNAse-free water. The following thermal cycling parameters were used: one cycle at 95°C for 1 minute, followed by 40 cycles of 94°C for 15 seconds, and 60°C for 1 minute. Fluorescent product was detected at the last step of each cycle. Melting curve analysis was performed for each reaction to ensure a single peak indicating selectivity for the target gene. The reference housekeeping gene GAPDH was utilized for normalization of PCR data, using the GAPDH-specific primers 5′-GGCACAGTCAAGGCTGAGAATG-3′ (forward) and 5′-ATGGTGGTGAAGACGCCAGTA-3′ (reverse). Cycle thresholds (Ct) were automatically determined within the log-linear phase of the reaction using Sequence Detection Systems plate utility software (Applied Biosystems). The comparative cycle threshold method (ΔCt method) was used to calculate relative expression levels (10).
Western blotting
Hippocampi dissected out from pups at three ages (PND 3, PND 7, and PND 21) were washed with ice-cold 1 × phosphate-buffered saline and homogenized by sonication in radioimmunoprecipitation assay buffer containing protease inhibitors for 1 minute. Homogenates were incubated on ice for 15 minutes followed by centrifugation at 12,000 rpm for 20 minutes at 4°C to precipitate debris. The supernatant was collected, and the protein concentration was measured by the BCA Protein Assay Reagent Kit (Bio-Rad, Laboratories). Samples (50 μg) were fractionated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (10% gel for Rap1 and 15% gel for BDNF). Protein was transferred onto a polyvinylidene fluoride membrane (Bio-Rad Laboratories) and blocked with 5% skim milk in Tris-buffered saline including 0.1% Tween-20 (TBS-T buffer) for 2 hours at room temperature, and then subjected to immunoblot analysis with antibodies to Rap1 (1:1000; Cell Signaling) and BDNF (1:1000; Chemicon). After incubation in primary antibodies, immune complexes were detected with horseradish-peroxidase-conjugated secondary antibodies (Bios). Chemoluminescence was detected using ECL reagent (GE).
Statistical analysis
All values were expressed as mean ± standard error of the mean. For behavioral tests, escape latency times in the hidden platform trial were analyzed with two-way analysis of variance (ANOVA) of repeated measures, while one-way ANOVA was conducted on the data obtained from the testing phase using statistical software (SPSS12.0). Following ANOVA, the Student–Newman–Keul's post hoc analysis was used for pair-wise comparisons of data. Differences were considered statistically significant at p < 0.05.
Results
Maternal and pup assessment
Manipulation of thyroid status in dams produced the expected effects on circulating levels of TH. Sub and Hypo dams exhibited significantly elevated serum TSH compared with euthyroid controls (p < 0.05 and p < 0.01, respectively; Table 1). Hypo dams exhibited significantly lower circulating levels of total T4 (for all days tested; p < 0.01); however, there were no significant differences between the Sub group and euthyroid controls in total T4. There were no significant differences in litter sex ratio at P1 between the Sham control and thyroid manipulated groups. The average pup BW in the Sub group and Hypo group was slightly but not significantly lower than that in the control group on PND 21. On PND 40 there were no significant differences in TSH and total T4 levels between the Sham control and Sub group pups (p > 0.05; data not shown). However, TSH level was higher and total T4 level was lower in Hypo group than those in control group and Sub group pups (p < 0.01; data not shown). The mean BW of pups on PND 40 were as follows: control group, 122.11 ± 3.66 g; Sub group, 117 ± 4.75 g; and Hypo group, 90.90 ± 4.92 g (control vs. Hypo group; p < 0.01).
MWM test
Spatial learning was assessed using the MWM. There was no sex difference in the escape latency in the MWM over the trial days for any of the three groups (p > 0.05; data not shown). Therefore, male and female data were grouped together for analysis. The mean escape latency did not differ between any of the three groups on the first 3 days of training, but there were significant differences in the mean escape latency between the Sham control and Sub group as well as between the Sub and Hypo groups on the last day of training (PND 44; Fig. 2A).
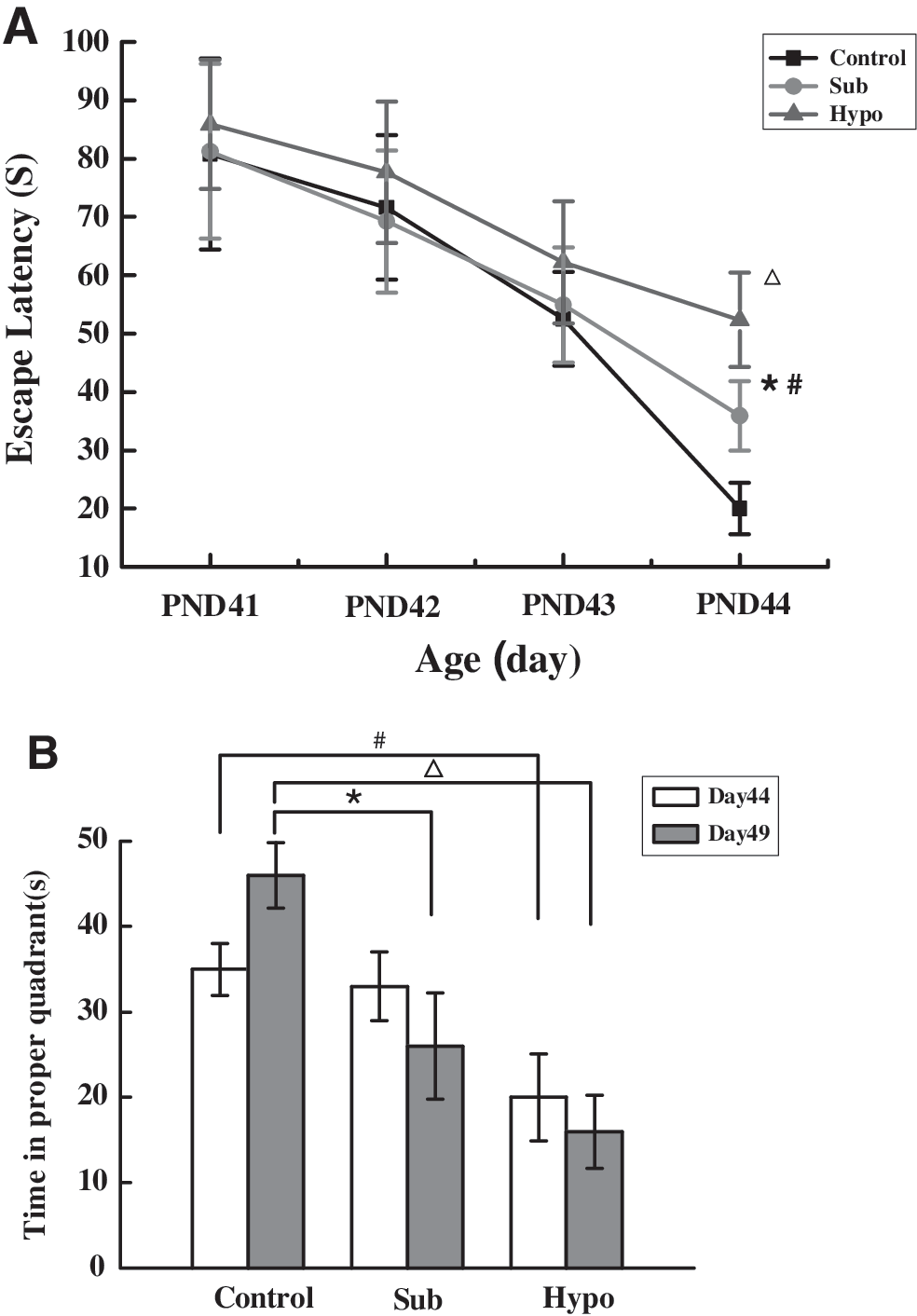
Performance of pups in the MWM task. Data are mean ± SEM. (
There were no significant differences among the three groups in swimming velocity during testing for short-term or long-term memory. There was a tendency toward a deficit in short-term memory (measured on PND 44) in the Sub group, as this group spent a shorter amount of time in the target quadrant. However, this trend did not reach statistical significance. The long-term memory testing showed that the Sub and Hypo groups spent less time in the target quadrant than controls (p < 0.05 for all comparisons; Fig. 2B) indicating a memory deficit.
Effect of maternal subclinical hypothyroidism during pregnancy on BDNF and Rap1 expression in the hippocampi of rat pups
Real-time PCR showed that BDNF mRNA expression was downregulated at PND 3 in the Sub group compared to the Sham control group (p < 0.05; Fig. 3C). Western blot analysis showed that the level of BDNF protein expression was lower in the Sub group than in the Sham control group at PND 3 and PND 7 (p < 0.05; Fig. 3A, B). BDNF mRNA and protein expression was significantly decreased in the Hypo group at PND 3, PND 7, and PND 21 when compared with the control and Sub groups (all p < 0.01; Fig. 3A–C).
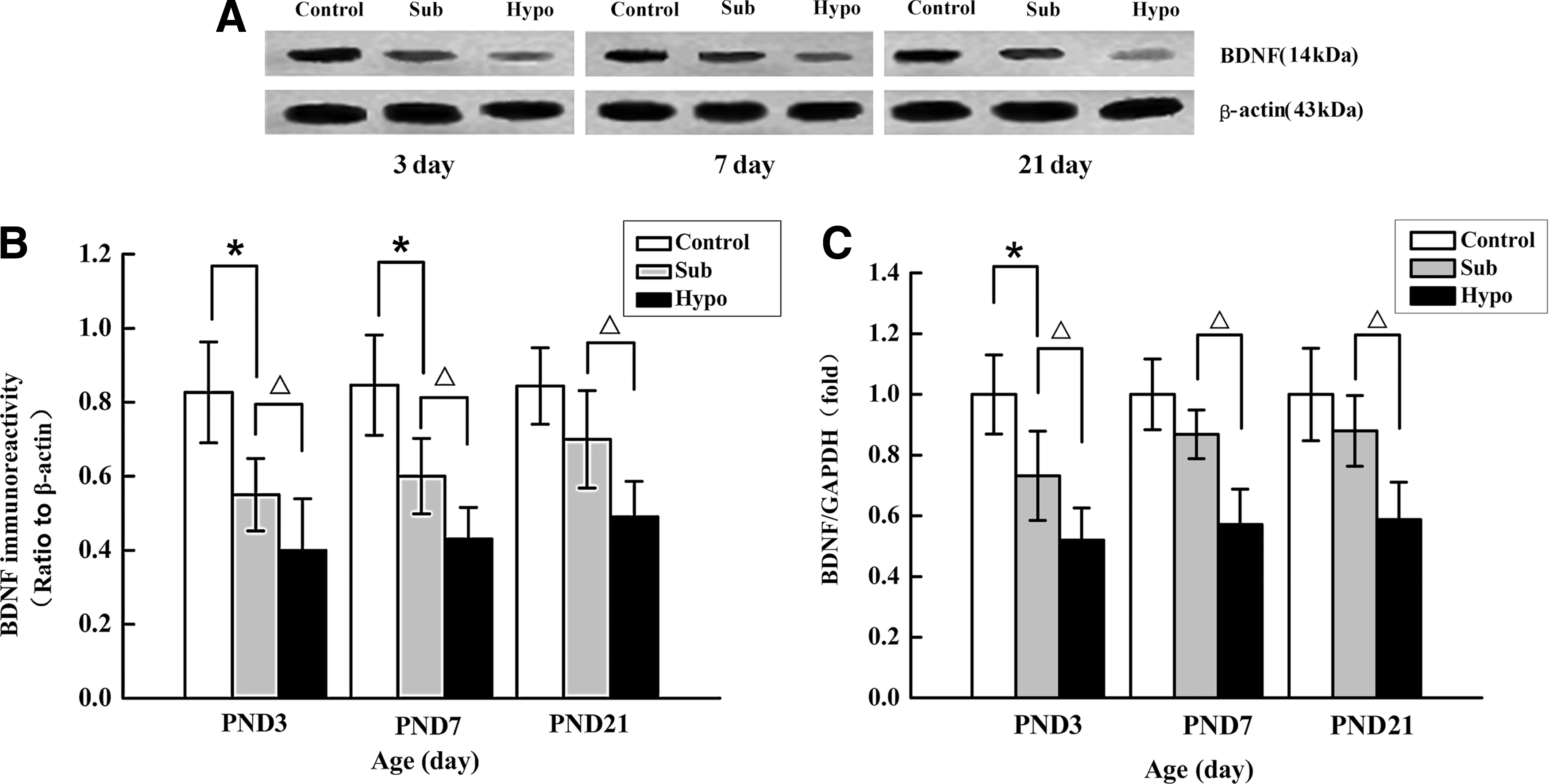
Expression of BDNF protein and mRNA levels in the developing rat pup hippocampus during various stages of postnatal development. (
The results of Western blot analysis for Rap1 are shown in Figure 4. Expression of Rap1 was slightly increased in Sub and Hypo group at PND 3, but there was no significant difference compared with control group. A statistically significant upregulation for Rap1 was observed in the Sub and Hypo groups at PND 7 and PND 21 when compared with the Sham control group (p < 0.05 and p < 0.01, Sub and Hypo, respectively).
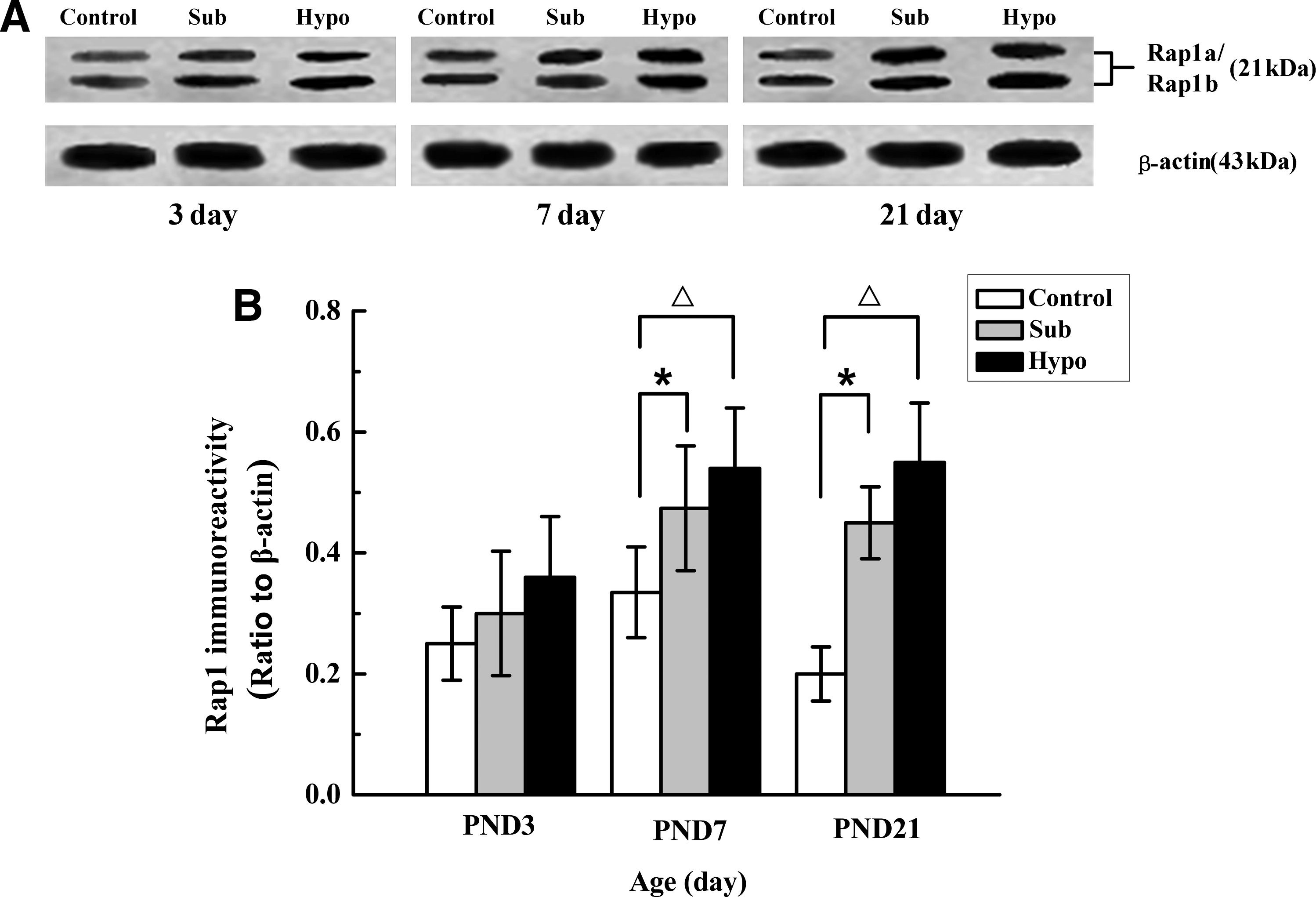
(
Effect of maternal subclinical hypothyroidism during pregnancy on NCAM gene expression in the hippocampi of rat pups
In agreement with previously published results (7), NCAM mRNA expression was upregulated in the Hypo group at PND 3, PND 7, and PND 21 compared to the Sham control group (p < 0.05; Fig. 5). However, no significant differences in NCAM mRNA levels were observed between the Sub group and Sham control group (p > 0.05).
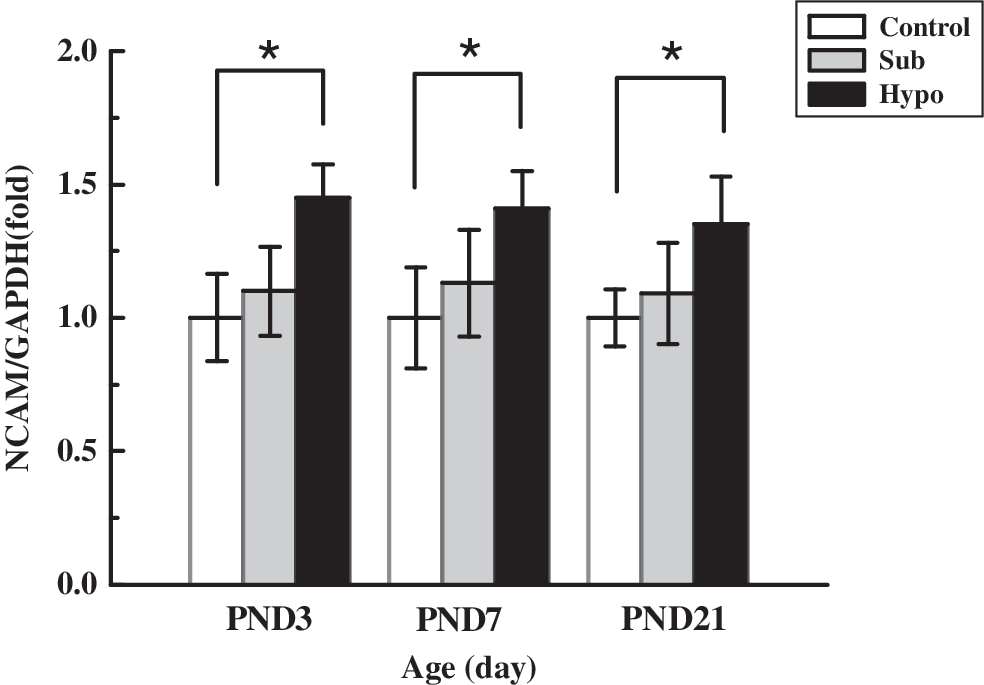
NCAM mRNA expression in the developing rat pup hippocampus. NCAM expression was found significantly increased in the Hypo group at PND 3, PND 7, and PND 21 (*p < 0.05). No significant changes in NCAM mRNA levels were detected between the Sub and Sham control groups (p > 0.05). Values for each individual were normalized with the reference gene GAPDH. Each bar represents the mean ± SEM. NCAM, neural cell adhesion molecule.
Discussion
Before midgestation the mother is the only source of T4 for the developing fetal brain. It has been suggested that TH is required for normal corticogenesis in the rat during early gestation (5,11,12). Escobar et al. (8) found neuronal migratory defects in the fetal cortex and in the CA1 and CA3 regions of the hippocampus resulting from mild maternal hypothyroxinemia. These defects could potentially cause behavioral deficits seen in these offspring. However, few in vivo experimental studies have focused on clarifying whether or how maternal subclinical hypothyroidism affects brain development during gestation. This study demonstrated for the first time that maternal subclinical hypothyroidism during pregnancy affects TH responsive gene expression and a decline in memory performance in offspring.
The neurotrophin family member BDNF plays a key role in neuronal differentiation and memory consolidation. These roles include calcium-dependent modification of dendritic structure and synaptogenesis, as well as the formation of long-term potentiation (LTP) in the hippocampus (13). BDNF is involved in LTP-related cytoskeletal changes and memory storage (14). Intrahippocampal administration of BDNF affects short-term behavioral plasticity in adult rats (15). Previous studies have reported that maternal hypothyroidism significantly reduced BDNF expression in the rat pup hippocampus (11,16). This study found that BDNF protein expression was decreased in the developing pup hippocampus from both subclinical and hypothyroid mothers. Further, this downregulation of BDNF protein was correlated with a decrease in BDNF mRNA expression during the early PND 3, but not at later time points (PND 7 and PND 21). A hypothetical explanation for these findings is that the effects of maternal subclinical hypothyroidism could produce changes at the level of proteins rather than more permanent changes in mRNA expression. Further, these results reinforced the hypothesis that the developing rat is more sensitive to thyroid disruption than the adult rat (16).
Rap1 is a member of the Ras family of small GTP-binding proteins and is directly regulated by THs during neural development (17). Activated Ras induces synaptic strengthening and is critical for LTP (18,19). In neurons, Rap1 signaling can antagonize the Ras-ERK signaling pathway, thereby negatively regulating synaptic strength. Rap1 is an important downstream signaling molecule that mediates BDNF stimulation of ERK in the central nervous system. BDNF induces the rapid and transient activation of MAP kinase in a Ras-dependent manner (20). In this study through comparisons with the control group we found that Rap1 protein remained at a higher level in the hippocampi of offspring from subclinical hypothyroidism dams at PND 7 and PND 21. We hypothesize that a persistent higher Rap1 level may antagonize the Ras-ERK signaling pathway, which may participate in the effect of mild maternal thyroid deficiency on the developing brain.
A previous study (7) found through in situ hybridization that NCAM expression was increased in the hypothyroid rat brain at PND 15 and PND 20. While NCAM is important for normal memory functions and is regulated by TH in the neonatal rat brain, expression of this molecule in offspring was not significantly affected by mild maternal thyroid manipulation (Sub group) in this study. It is possible that compensatory mechanisms restore normal levels of NCAM in the face of a mild reduction of TH. These results suggest that NCAM may not be involved in the memory impairment of pups from subclinical hypothyroidism dams.
The MWM tests hippocampus-dependent spatial memory, the closest parallel to episodic memory in humans (21). The results of this study demonstrated equal acquisition and retention of learning in the water maze by rats in all three groups (no significant differences among the three groups during training). However, a clear deficit in both short-term and long-term spatial memory was observed in maternal subclinical hypothyroidism offspring.
In summary, the results of this study indicated an effect of maternal subclinical hypothyroidism on the developmental and behavioral deficits found in offspring. Maternal subclinical hypothyroidism depressed expression of BDNF at both the genetic and protein levels and upregulated Rap1 protein expression early in postnatal development. Both BDNF and Rap1 have been shown to play important roles in cognitive performance.
Footnotes
Acknowledgments
This study was supported by grants from the Healthcare Industry Special Project (200802008), the Social Development Foundation of Liaoning Province ([2005]36–39), and the Key Laboratory Support Special Program of Liaoning Province (20070905), Shenyang, all in China.
Disclosure Statement
The authors declare that no competing financial interests exist.