
Abstract
Thyroid hormone (TH) receptor (TR) plays critical roles in vertebrate development. Transcription studies have shown that TR activates or represses TH-inducible genes by recruiting coactivators or corepressors in the presence or absence of TH, respectively. However, the developmental roles of these TR cofactors remain largely unexplored. Frog metamorphosis is totally dependent on TH and mimics the postembryonic period in mammalian development during which TH levels are also high. We have previously proposed a dual function model for TR in the development of the anuran Xenopus laevis. That is, unliganded TR recruits corepressors to TH-inducible genes in premetamorphic tadpoles to repress these genes and prevent premature metamorphic changes and subsequently, when TH becomes available, liganded TR recruits coactivators to activate these same genes, leading to metamorphosis. Over the years, we and others have used molecular and genetic approaches to demonstrate the importance of the dual functions of TR in Xenopus laevis. In particular, unliganded TR has been shown to recruit histone deacetylase-containing corepressor complexes in premetamorphic tadpoles to control metamorphic timing. In contrast, metamorphosis requires TH-bound TR to recruit coactivator complexes containing histone acetyltransferases and methyltransferases to activate transcription. Furthermore, the concentrations of coactivators appear to regulate the rate of metamorphic progression. Studies in mammals also suggest that the dual function model for TR is conserved across vertebrates.
Introduction
During human development, TH begins to function very early (3,6,11). The thyroid gland is developed within the first 12 weeks of gestation. In vitro organ culture studies of human thyroid tissue suggest that fetal TH production begins as early as 10 weeks of gestation, although significant TH production probably does not occur until 20 weeks or later (11). In the fetal plasma, TH levels, especially 3,5,3′-triiodothyronine (T3), remain low until about 4 months prior to birth when endogenous synthesis of TH leads to a rapid increase in plasma TH levels (3,11). The TH levels reach a peak at birth and remain high several months after birth. Thus, it may not be surprising that TH deficiency during the neonatal period leads to irreversible, profound neurological deficit and mental retardation (6,12,13). In addition to fetal TH, maternal TH also plays an important role in early embryogenesis (14,15). On the other hand, excess TH also impairs embryo development, leading to increased miscarriages and lower birth weights (16). Although TH is clearly important for embryogenesis in humans and other mammals, it has been difficult to investigate the underlying mechanisms in mammals. This is in part due to the difficulty to manipulate the uterus-enclosed mammalian embryos and to separate the direct effects of TH on embryos from the indirect maternal effects caused by altering TH levels.
Amphibian Metamorphosis as a Model for Vertebrate Development
Amphibian metamorphosis is a process that is totally dependent on TH. The metamorphosis of anurans has been studied as a model of TH action for about a century (17,18). This process involves extensive changes in essentially every single organ/tissue of the tadpole (4,19). Tadpole-specific organs such as the tail and gills are completely resorbed, while adult specific organs such as the limbs develop de novo. Most of the organs and tissues such as the intestine and brain are present in both the tadpole and frog but undergo drastic remodeling often involving degeneration of larval tissues and development of the adult ones. Despite the diverse changes, all are under the control of TH in an organ-autonomous manner for most, if not all organs (3,4,19).
Anuran metamorphosis bears strong similarities to the postembryonic development in mammals at morphological and molecular levels. For example, the adult intestines of both frogs and mammals have similar structural organizations with extensive epithelial folding to increase the luminal surface for nutrient absorption (20). Anuran and mammalian intestines also share similar developmental pathways. They both initially develop into a simple tubular structure consisting of mostly endoderm-derived epithelial cells. While this initial simple organ is more developed in anuran tadpoles for it to be functional in an independent living tadpole, both the amphibian and mammalian larval/embryonic intestines subsequently undergo similar morphogenetic transformations to form the complex adult organ (20,21).
Likewise, many other processes that occur during amphibian metamorphosis also take place during postembryonic development in mammals (3 –5). These include the switching of the hemoglobin genes from larval (tadpole) or fetal types to frog or adult types, respectively, the dramatic increase in the serum albumin levels, skin keratinization, the induction of urea cycle enzymes, and the developmental progression and restructuring of the central and peripheral nervous systems, etc. Additionally and most obviously, both mammals and (most) anurans change their living habitat after this postembryonic period, from an aquatic (amniotic) to a terrestrial environment, accompanied by the maturation of the lung for air breathing. All these changes occur in the presence of high levels of plasma TH during both metamorphosis and mammalian postembryonic development. More importantly, the metamorphic changes in tadpoles occur in a free-living organism and can be induced by the addition of physiological levels of TH to the rearing water, making study of postembryonic development in frogs an ideal model.
Gene Regulation by TH Receptor (TR)
The biological effects of TH are mainly mediated through gene regulation by TRs, although nongenomic effects of TH have been well documented (1,2,22 –25). There are two evolutionary-conserved TR genes, TRα and TRβ, in all vertebrate species and both are members of the nuclear hormone receptor superfamily (1,2,4,22,24,26). TRs are capable of functioning as monomers, homodimers, and heterodimers with 9-cis-retinoic acid receptors (RXRs), which belong to the same superfamily. For genes induced by TH, TR-RXR heterodimers bind to TH response elements (TREs) in/around the promoters of target genes even in the context of chromatin (27). In the absence of TH, TR represses transcription from their promoters; when TH is present, TR activates their transcription, accompanied by chromatin disruption (27 –29). There are also genes that are repressed by the presence of TH. Much less is known about how TH represses these genes. Thus, the discussions here will focus only on TH-induced gene expression.
TR accomplishes gene regulation by recruiting corepressor or coactivator complexes to the target promoters, respectively (Fig. 1A). Many TR-interacting proteins have been isolated and characterized through in vitro and cell culture studies (2,30 –45). Two highly related proteins N-CoR (nuclear corepressor) and SMRT (silencing mediator of retinoid and thyroid hormone receptors) are among the best-studied TR-binding corepressors. Both bind to unliganded TR and form large complexes containing histone deacetylase 3 (HDAC3), transducin beta-like protein 1 (TBL1)/TBL1-related protein 1 (TBLR1), and G-protein pathway suppressor 2 (GPS2) (Fig. 1A) (30 –34,36,46 –50), suggesting that unliganded TR represses transcription in part through deacetylation of the local histones at target promoters.

(
In the presence of TH, TR releases corepressors and recruits coactivators. The known TR coactivators include ATP-dependent chromatin-remodeling proteins, histone acetyltransferases such as CREB-binding protein (CBP) and p300, and TRAP/DRIP/mediator complex that associates with the recruitment and activation of RNA polymerase II (2,35 –42,51,52). Among the coactivators, the role of the SRC family (SRC1, 2, 3) in gene regulation by TR has been studied extensively. SRCs bind to TR in the presence of TH and form complexes with other cofactors, including CBP/p300, protein arginine methyltransferase 1 (PRMT1), and coactivator-associated arginine methyltransferase 1 (CARM1 or PRMT4) (38,40 –42,53 –58), suggesting that SRCs activate transcription in part through acetylation or methylation of local histones in the promoter region (Fig. 1A).
TR Function During Frog Development
A dual function model
Based on the developmental expression of TRs and RXRs and the ability of TR to activate and repress TH-inducible genes in a TH-dependent manner, we have previously proposed a dual function model for TR during the development of the South African clawed toad Xenopus laevis (Fig. 1B) (59,60). According to the model, TH-inducible genes are not regulated by TH or TR when the hormone and receptor levels are low or absent. Thus, the lack of activation or repression by the thyroid system allows for normal embryonic development. By stage 45 when a free feeding tadpole is formed upon the completion of embryogenesis (61), the embryonic roles of these genes have past. The increased expression of TR and RXR genes just after embryogenesis, especially TRα and RXRα, leads to the formation of TR-RXR heterodimers that bind to the TREs of these genes, leading to the recruitment of corepressor complexes containing N-CoR or SMRT due to the lack of TH. This results in the repression of the genes via histone deacetylation. The hypothesis here is that this gene repression is important to ensure proper tadpole growth and to prevent premature metamorphic organ transformations.
After stage 55, the synthesis and secretion of endogenous TH elevate the levels of TH in the plasma and tadpole tissues. The binding of TH to chromatin-bound TR releases the corepressor complexes from the target genes with concurrent recruitment of the coactivator complexes, such as those containing SRC-p300-PRMT1 (Fig. 1B). This results in local histone acetylation and methylation and gene activation, leading to metamorphic transformations in the tadpoles.
In vivo evidence for the dual function model
The first in vivo functional study in X. laevis showed that in early Xenopus embryos lacking endogenous TR and TH, overexpression of TR upon microinjection of its mRNA into fertilized eggs was capable of repressing or activating endogenous gene expression (62). Interestingly, overexpression of TR and RXR together, but not either one alone, represses endogenous TH response genes while the addition of TH leads to the reversal of the repression and further activation of the TH response genes. This suggests that TRs function as heterodimers with RXR in vivo and that endogenous TR and RXR normally expressed later at tadpole stages should repress or activate these genes in the absence or presence of TH, respectively.
To determine whether TR binds to endogenous target genes during development, we adapted the chromatin immunoprecipitation assays (ChIP) to analyze the binding of endogenous TR to endogenous target genes in whole embryos/tadpoles or tadpole tissues (63). This assay provided us with direct evidence that there is little or no binding of TR and RXR to endogenous TREs during embryogenesis while both are bound to TREs in tadpoles as predicted by the model. Subsequently, we and others have shown that TRs bind to target promoters in different organs independently of TH in both X. laevis and a related species X. tropicalis and more importantly recruit coactivators and corepressors as the model predicts (Fig. 2) (57,63 –70).
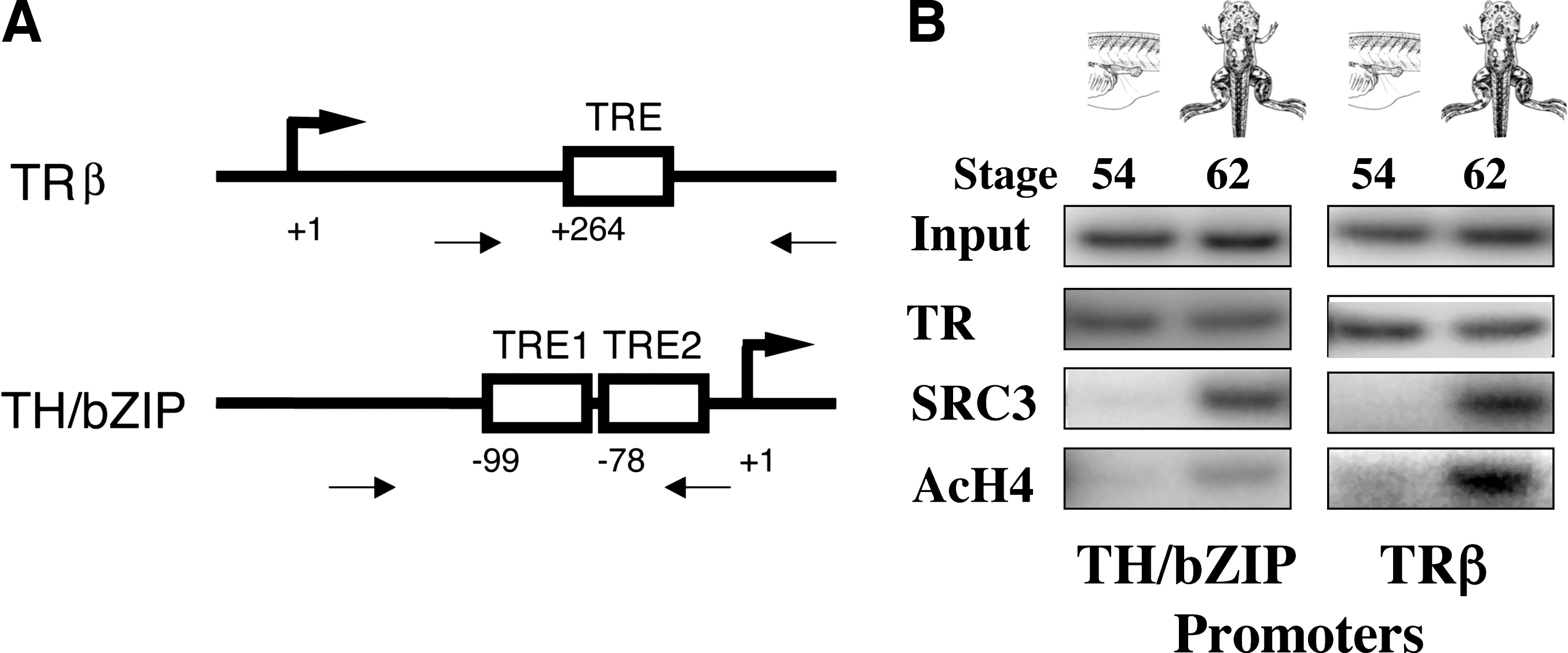
TH-dependent recruitment of coactivator SRC3 and histone acetylation at TH-responsive promoters, TRβA and TH/bZIP, in the intestine of tadpoles during natural metamorphosis. (
Several different laboratories have investigated the requirement for TR-dependent gene activation during metamorphosis by using a dominant negative TR. This mutant TR contains a C-terminal deletion that prevents ligand binding and coactivator recruitment. Overexpression of this dominant negative TR through transgenesis or in vivo gene transfer inhibits TH-induced gene activation and metamorphosis in X. laevis (71 –75). The dominant negative TR expressed in transgenic animals has been shown to compete against endogenous TR for binding to endogenous target genes, leading to the retention of corepressors at the target genes even in the presence of TH (75). These findings indicate an essential role of TH-bound TR in gene regulation and metamorphosis.
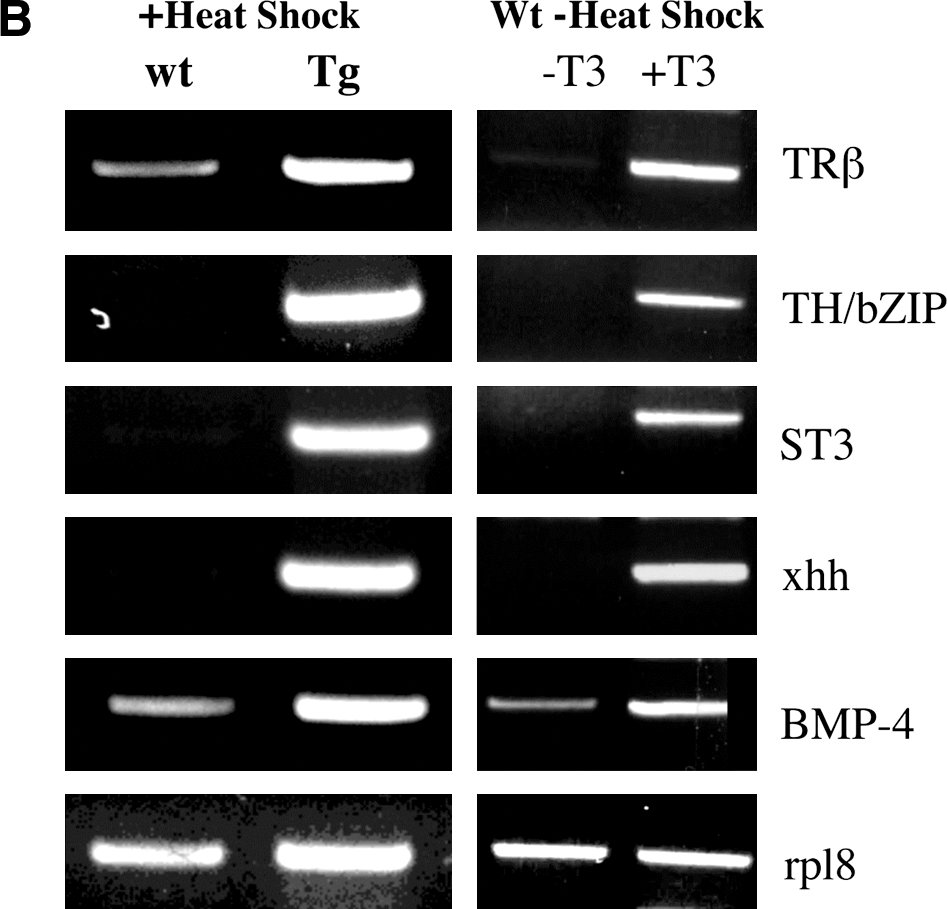
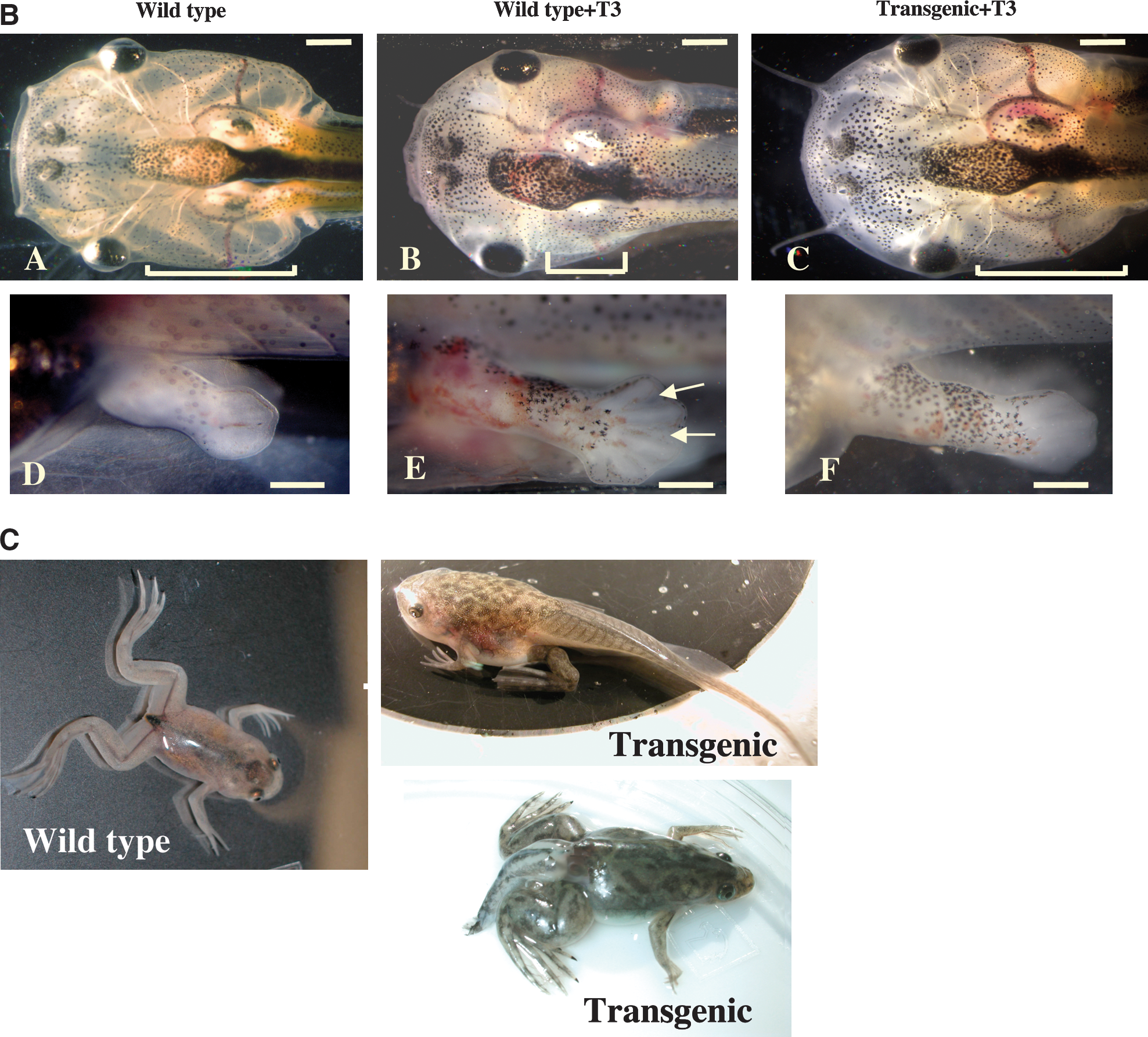
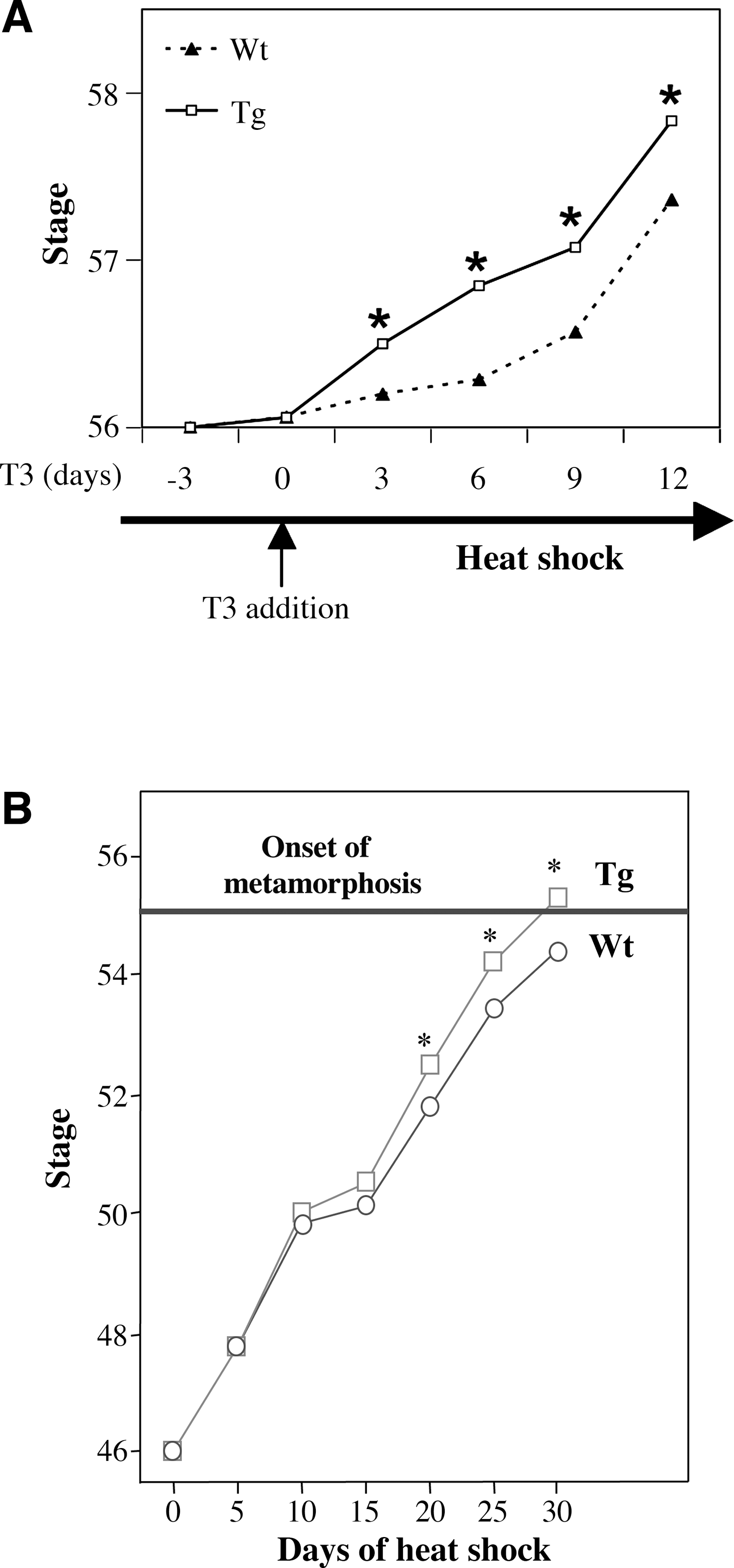
To determine whether TR is sufficient to mediate the metamorphic effects of TH, we have generated a dominant positive TR that cannot bind to TH (due to a small deletion at the C-terminus) but constitutively activate transcription (due to the N-terminal fusion of the strong viral activator VP16) (76). As early embryogenesis is adversely affected by inappropriate TR function, especially the activated (TH-bound) form of TR (62), we introduced the dominant positive TR under the control of a heat shock-inducible promoter into developing animals through transgenesis. When premetamorphic transgenic tadpoles and their wild-type siblings were subjected to heat shock treatment to induce the transgene expression, within several days the transgenic tadpoles underwent metamorphic changes typical of TH-treated premetamorphic tadpoles, a phenomenon not observed in the wild-type siblings (Fig. 3A) (76). Furthermore, ChIP assays have shown that the dominant positive TR is bound to endogenous target genes (76). This leads to the activation of the same genes that are normally induced by TH (Fig. 3B) (76). Thus, TR is not only necessary but also sufficient to mediate the effects of TH on all the parameters that were measured.
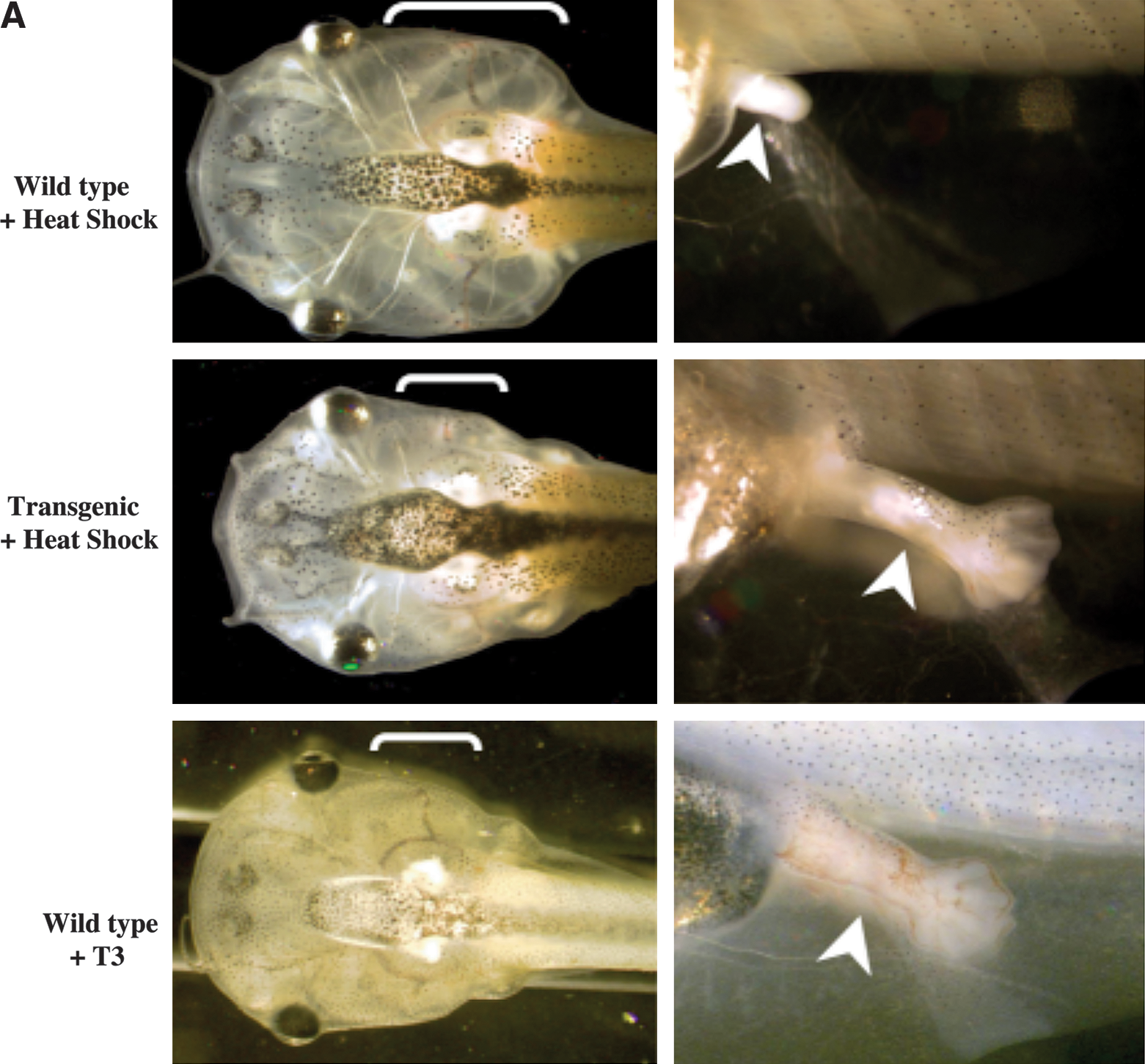
Expression of a TH-independent, constitutively active, dominant positive TR in transgenic tadpoles initiates metamorphosis. (
Roles of TR Cofactors During Frog Development
Coactivator recruitment is essential for metamorphosis
There are two components in the transcriptional activation by TH, the relief of the repression caused by unliganded TR and additional activation due to liganded TR, with the former representing the major fraction of the gene induction by TH in many reporter assays. The relative contributions from corepressor release and coactivator recruitment upon TH binding to TR have been difficult to determine in vivo. Cofactor knockout mice often have relatively mild phenotypes due to cofactor redundancy or embryonic lethal phenotypes, thus revealing little information about their roles in development. In addition, many cofactors function with multiple transcription factors and this makes it difficult to pinpoint the role of a particular cofactor to a nuclear hormone receptor pathway even when gene knockout and transgenesis result in easily identifiable phenotypes, such as mice deficient in N-CoR, p300, SRC1-3, or TRAP220 (77 –82). Frog metamorphosis, however, is totally dependent on TH. There has been no evidence that any ligands for other nuclear hormone receptors can induce morphological changes in premetamorphic tadpoles in the absence of TH, although some can modulate TH-induced changes (4,83). Furthermore, TR is both necessary and sufficient for mediating the effects of TH. This makes it possible to correlate developmental phenotypes during metamorphosis with specific cofactor function in the TR signaling pathway. We have focused our effort on SRC coactivator complexes. We have previously shown that the coactivators SRC3, p300, PRMT1, and CARM1 are expressed during X. laevis metamorphosis with the expression of SRC3 and PRMT1 upregulated indirectly by TH (57,58,84). Furthermore, ChIP assays by us and others have shown that SRC3, p300, and PRMT1 are recruited by TR to endogenous target genes in a TH-dependent manner (Fig. 2) (57,64 –66,85). Surprisingly, SRC3 recruitment to target genes is gene and organ dependent. It is recruited to the promoter regions of both of the two endogenous target genes analyzed, TH/bZIP (a transcription factor gene) and TRβ, in the intestine but only to the TH/bZIP gene in the tail (65), in agreement with those reported by Havis et al. (66). These results suggest that tissue content and promoter context affect cofactor specificity.
To investigate the role of coactivator recruitment during frog metamorphosis, we generated transgenic tadpoles constitutively overexpressing a Flag-tagged dominant negative form of SRC3, which contains only the receptor interacting domain of Xenopus SRC3 under the control of the ubiquitously expressing CMV promoter (64). The mutant SRC3 is recruited by liganded TR to endogenous target genes, thus competing away the endogenous wild type SRC3 and presumably other TR-binding coactivators as well (Fig. 4A) (64). As expected, it does not affect the TH-induced release of corepressors (Fig. 4A). Interestingly, despite the release of the corepressors upon TH treatment of the transgenic tadpoles, there is little change in histone acetylation at the target genes, indicating that the release of histone deacetylase-containing corepressors is not sufficient to cause significant increases in local histone acetylation in developing tadpoles (Fig. 4A). Surprisingly, but consistent with the histone acetylation result, there is little induction of TH-inducible genes upon TH treatment of the transgenic animals (64). Thus, corepressor release has relatively small contributions to the TH-induced increases in histone acetylation and transcription of endogenous target genes in developing tadpoles, unlike those observed in many studies with reporter genes in cell cultures and other model systems. In addition, morphological analyses have shown that the dominant negative SRC3 inhibits essentially all aspects of TH-induced metamorphosis (Fig. 4B) and also inhibits or delays natural metamorphosis (Fig. 4C) (64). These results indicate that coactivator recruitment is required for gene activation by TH and metamorphosis. Our findings are consistent with those from a study with a TR antagonist by Lim et al. (86), where the TR antagonist NH-3 was shown to prevent the binding of SRC coactivators to TR and inhibits metamorphosis as well as gene activation by TR in tadpoles. They further suggest that corepressor release is a minor component of TH signaling needed to effect postembryonic organ remodeling during metamorphosis. On the other hand, as described below, premature release of corepressors in premetamorphic tadpoles can alter development.
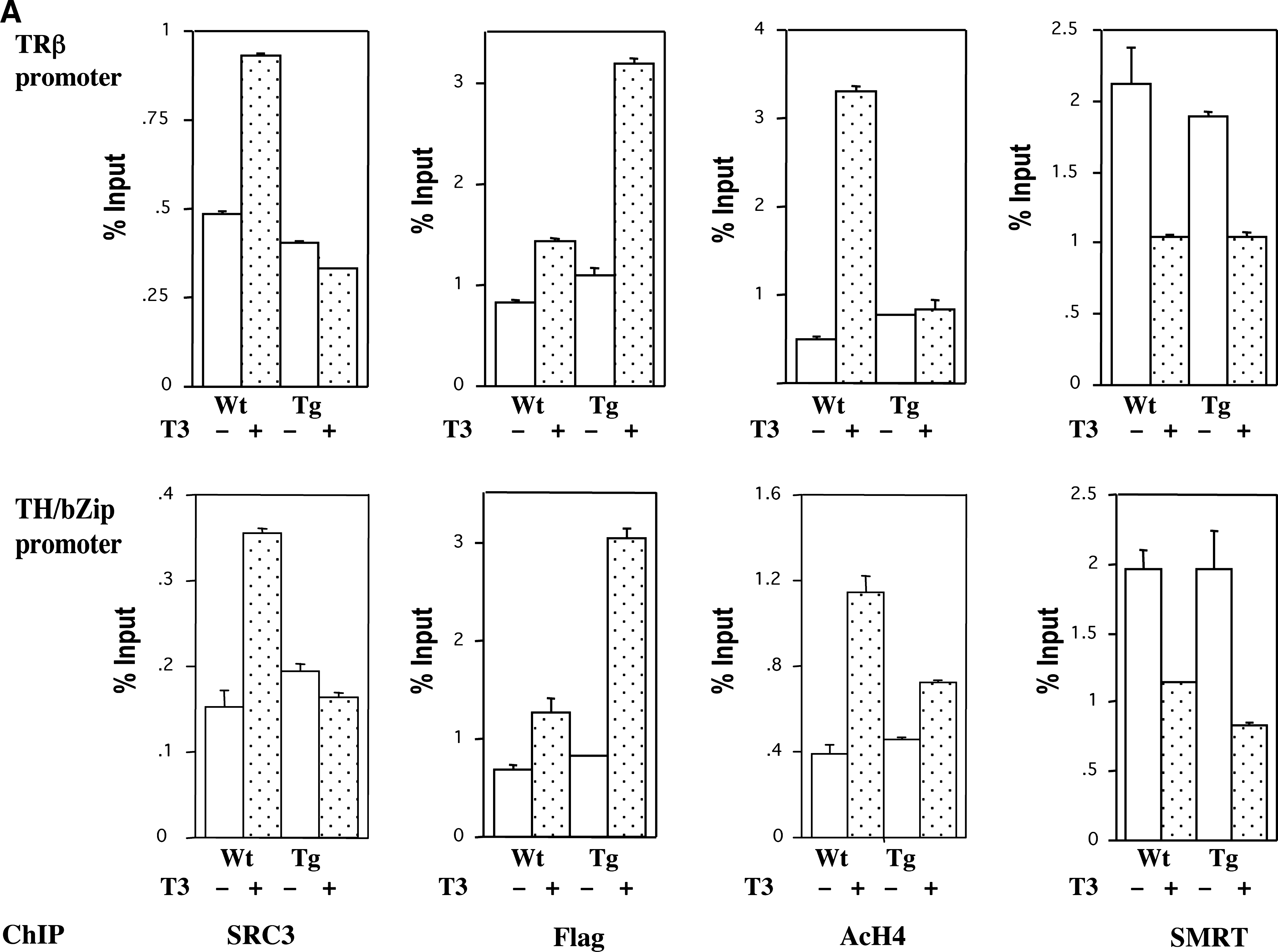
Transgenic, constitutive expression of a Flag-tagged dominant negative coactivator SRC3, containing the receptor interaction domain of Xenopus laevis SRC3, inhibits natural and TH-induced metamorphosis by competing for recruitment to endogenous target genes. (
Since the dominant negative SRC3 blocks the binding of all coactivators to liganded TR, it remains possible that SRC family members do not play an essential role in gene regulation by TR and metamorphosis. To determine whether and how SRCs participate in metamorphosis, we have investigated the role of SRC-binding protein p300 during metamorphosis. As stated above, Xenopus p300 is recruited to TH-responsive promoters in a TH-dependent manner (66,85). To investigate whether complexes containing SRCs and p300 or its related protein CBP are required for metamorphosis, we have generated a Flag-tagged dominant negative form of p300 that contains only the SRC-binding domain, which is expected to bind to SRCs and thus disrupt the function of SRC-p300/CBP complexes but not other coactivator complexes such as TRAP/DRIP/mediator complex. Transgenic expression of this dominant negative form of p300 under the control of the CMV promoter has similar effects as the dominant negative form of SRC3 (85). That is, it inhibits TH-induced gene activation and metamorphosis as well as natural metamorphosis. Since this dominant negative form of p300 does not interfere with the direct binding of coactivators to liganded TR but specifically interferes with the function of SRC-p300/CBP complexes, these results demonstrate an essential role of such complexes in gene regulation by TR and metamorphosis.
Coactivator levels regulate rate of metamorphic progression
To further analyze the role of SRC-p300 complexes during metamorphosis, we have taken a complementary approach to the above function-inhibition experiments. We have analyzed the role of another component of the SRC-p300 complexes, namely, PRMT1, a histone methyltransferase (57). In the presence of TH, PRMT1 is highly upregulated in the tadpole intestine and is recruited to endogenous TH-target genes during metamorphosis. When a Flag-tagged wild-type Xenopus PRMT1 was introduced into transgenic tadpoles under the control of a heat shock–inducible promoter, heat shock treatment led to ubiquitous overexpression of the Flag-tagged transgenic PRMT1 and its recruitment to target genes in a TH-dependent manner, accompanied by increased target gene expression. More importantly, overexpression of PRMT1 also accelerated both natural and TH-induced metamorphosis (Fig. 5A) (57). Thus, coactivator levels appear to influence the rate of the progression of metamorphosis in different organs/tissues.
Cofactors regulate developmental rate and timing. (
Corepressor binding to unliganded TR controls metamorphic timing
The dual function model for TR predicts that gene repression by unliganded TR is important for proper development and this repression utilizes corepressors. Many studies have firmly established that N-CoR/SMRT complexes participate in gene repression by unliganded nuclear receptors including TRs in cultured cells (32 –34,36,46 –50). Furthermore, transgenic overexpression of a dominant negative N-CoR in the liver relieves the repression by unliganded TR in mouse (87). In addition, specific gene targeting that mutated the receptor interacting domain of N-CoR, which resulted in a mutant N-CoR that cannot bind to TR, in the liver also abrogates the repression of TH-inducible genes in hypothyroid mice (88), demonstrating the importance of N-CoR in gene repression by unliganded TR in mouse liver in vivo. During Xenopus development, low levels of TRs are expressed during embryogenesis (89 –91). A recent study suggests that corepressor binding by unliganded receptor expressed during this period is essential for eye development (92). As reviewed above, upon the completion of embryogenesis and the formation of a free living tadpole by stage 45, TRα is highly expressed in X. laevis. When we injected dominant negative N-CoR peptides into the tail of premetamorphic X. laevis tadpoles, the expression of a co-injected, TR-dependent reporter gene was upregulated in the absence of TH (69), suggesting that endogenous TRs and corepressors are capable of repressing gene expression in tadpoles.
To directly investigate the role of corepressors in premetamorphic tadpoles, we have generated transgenic animals expressing a dominant negative form of Xenopus N-CoR, which contains two copies of the receptor interacting domain of N-CoR (93), under the control of a heat shock–inducible promoter. This dominant negative N-CoR competes effectively against endogenous N-CoR for binding to unliganded TR in the frog oocyte, leading to the derepression of a reporter gene injected into the oocyte nucleus (93). As this dominant N-CoR binds only to unliganded TR, it is expected to affect TR function only during the developmental stages 45 to 55 when TR expression is high while TH levels are low (Fig. 1B), a window period of about 30 days. Thus, we subjected transgenic tadpoles carrying the dominant negative N-CoR and their wild type siblings to daily heat shock treatment, starting at stage 46. Heat shock treatment led to overexpression of the dominant negative N-CoR and upregulation of most of the TH-response genes analyzed, although the extent of the increase (about twofold or less) was much less than that due to TH treatment (indicating that derepression represents only a minor fraction of the gene activation induced by TH in the developing tadpoles, consistent with the dominant negative coactivator studies above). More importantly, transgenic animals developed faster, reaching stage 55, the onset of metamorphosis, by as much as 7 days earlier than the wild-type siblings within the 30-day experiment (93) (Fig. 5B). These findings demonstrate that unliganded TR indeed recruits corepressors to repress gene expression in the premetamorphic tadpole to regulate the timing for the initiation of metamorphosis, thereby ensuring proper growth of the tadpole before its transformation into a frog, an important measure for the growth and survival of the postmetamorphic animal.
Conclusions
Anuran metamorphosis is one of the most dramatic events mediated by TR-dependent gene regulation. It serves as an excellent model to understand the function and molecular mechanism of transcriptional regulation by TR in vivo during postembryonic development in vertebrates. Studies by us and others have provided strong evidence to show that TR is both necessary and sufficient to mediate the metamorphic effects of TH. Furthermore, TR has dual functions during development, as an unliganded repressor to repress gene expression to ensure a proper premetamorphic tadpole growth period and later as a TH-bound activator to induce metamorphosis. These dual roles are exerted by recruiting corepressors and coactivators, respectively. While this basic dual function model is well supported, the detailed molecular pathways controlling metamorphosis in different tissues are likely to be much more complex. For example, the two TRs, TRα and TRβ, are likely to have different functions, in part due to their distinct expression patterns and in part due to the differences in their molecular properties.
Indeed, studies with a TR-specific agonist have provided evidence to support different roles for the receptors during development in X. laevis (94,95). In addition, much remains to be learned about the roles of TH target genes during metamorphosis. Metamorphosis in insects, particularly Drosophila, is also controlled by a member of the nuclear receptor family, the ecdysone receptor. Extensive genetic and molecular analyses have demonstrated that a complex gene regulation cascade regulates the metamorphic transformations and the existence of feedback mechanisms involving transcription factors encoded by ecdysone target genes (96,97). Such feedbacks appear also to exist during amphibian metamorphosis as a recent study has shown that a TH-induced transcription factor can bind to the TH-dependent promoter of the X. laevis TRβA genes to regulate TRβ expression (98). There are a number of other TH-induced transcription factors during Xenopus metamorphosis. It would be interesting to study how they participate in the process of metamorphosis.
While the dual function model has been substantiated during frog development, it is very likely to exist during higher vertebrate development as well. During mammalian development, TRs are expressed prior to the synthesis of endogenous TH (3,11). While some maternal TH is likely to enter the fetus, low levels of TH during early embryogenesis will undoubtedly leave many of the TRs unliganded in many organs/tissues. However, as shown during frog development, the effects of derepression are relatively mild, leading to relatively small increases in gene expression compared to the much bigger contribution from activation through ligand-dependent coactivator recruitment. On the other hand, such mild effects can have an important influence on development, e.g., controlling the timing of metamorphosis. Thus, one would expect that disrupting the repression by unliganded TR may also have relatively mild effects on development in mammals, which is consistent with the weak phenotypes of TR knockout mice (99,100).
Most genetic and molecular studies in mice have focused on the activating effects of liganded TR and demonstrated an essential role of TH in development. The role of unliganded TR in mammalian development has been difficult to ascertain (101). On the other hand, several studies suggest a role of unliganded TR in the development of at least some mouse organs. First, in mouse embryos where TH levels are low, TRα knockout increases the expression of several TH-response genes in the heart, and this is accompanied by a significant increase in heart rate of the embryos (102). In postnatal mice when TH levels are high, the expression of these same TH-response genes are lower in the knockout mice compared to the wild-type siblings and the heart rate is also lower in the knockout animals, opposite of those observed in the embryos. Thus, TRα appears to repress the target genes and to keep the heart rate low in the unliganded state in the embryos and does the opposite when TH levels are high after birth. Similarly, it has been reported that TRβ deletion leads to hearing loss in both mouse and human and causes major defects in the photoreceptor cells in mouse, while mutations in TRβ that result in the formation of dominant negative forms of TRβ have relatively mild effects on the auditory and visual systems (103 –108). These findings suggest that unliganded TRβ is important for the development of the auditory and visual systems in higher vertebrates. In support of this, deleting the gene encoding the TH-inactivating enzyme type 3 deiodinase also leads to auditory deficits (109), suggesting unliganded TRs are important for the development of the auditory system. Thus, like frog development, mammalian development also utilizes the dual functions of TR to coordinate organ development and maturation. It would be of interest in the future to determine what corepressors and coactivators are recruited by TR to endogenous target genes to regulate the developmental processes in different organs/tissues in mammals.
Footnotes
Acknowledgment
I would like to thank Dr. D. Buchholz and current members in my laboratory for comments on the manuscript. The work in the author's laboratory has been supported by the Intramural Research Program of National Institute of Child Health and Human Development, National Institutes of Health.
Disclosure Statement
No competing financial interests exist.