
Abstract
Background:
We have shown substantial expression of type 3 deiodinase (D3, a major enzyme involved in the inactivation of thyroid hormone) in infiltrating leukocytes in several models of inflammation. Recently, thyroid hormone has been shown to improve remyelination in experimental autoimmune encephalomyelitis (EAE), an animal model of multiple sclerosis. As induction of D3 may play an important role in decreasing local bioavailability of thyroid hormone at inflammation sites, we hypothesized that D3 is induced in spinal cord inflammatory lesions in EAE.
Methods:
The aim of the study was to evaluate D3 expression in spinal cord inflammatory lesions of EAE Dark Agouti rats and to investigate D3 induction in activated monocytes.
Results:
Here, we show marked expression of D3 by granulocytes and macrophages in spinal cord inflammatory lesions of EAE rats. We further confirm induction of D3 expression in vitro in monocytes that were activated toward proinflammatory or immunomodulatory phenotypes.
Conclusions:
We observed increased D3 expression both in spinal cord inflammatory lesions during EAE and in activated monocytes. Although increased D3 expression theoretically results in decreased triiodothyronine availability, it is unknown at present whether reduced local triiodothyronine concentrations are involved in impaired remyelination as observed during EAE.
Introduction
Recent studies showed that thyroid hormone protects axons against damage induced by inflammatory processes during MS. Thyroid hormone activated OPCs in experimental allergic encephalomyelitis (EAE), an animal model of MS, by increasing myelin-basic protein content in the spinal cord of affected animals (9). Further, the pulsatile administration of thyroxine (T4) during the acute EAE improved remyelination by increasing OPC cell numbers and oligodendrocyte markers, resulting in faster morphological reorganization of myelin sheaths in the white matter during chronic EAE (10). Thyroid hormone has also been shown to improve remyelination in cuprizone-induced demyelination in mice (11). Recently, a study of Fernandez et al. showed that triiodothyronine (T3) completely restores the impaired oligodendrocyte generation observed in neurospheres derived form EAE rats, suggesting that T3 may favor remyelination in demyelinating disorders (12).
Extrathyroidal or peripheral thyroid hormone metabolism is predominantly mediated by deiodinating enzymes, which are organ specific and have different metabolic functions (13). D1 and D2 catalyze conversion of pro-hormone T4 into bioactive hormone T3, whereas type 3 deiodinase (D3) catalyzes inactivation of T4 and T3 into reverse (r)T3 and 3,3-diiodo-thyronine (T2), respectively. We have recently shown that D3 is expressed in infiltrating leukocytes during chemical (14) and bacterial inflammation (15). Substantial D3 expression may play an important role in decreasing local bioavailability of thyroid hormone at the inflammation sites. Since D3 is expressed in infiltrating leukocytes in several models of inflammation and in view of the possible role for thyroid hormone in the repairing capacity of oligodendrocytes, we tested the hypothesis that D3 expression is induced in inflammatory lesions during EAE.
Materials and Methods
Animal manipulations
Female Dark Agouti (DA) rats (Harlan, Boxmeer, the Netherlands), 7–8 weeks old, were immunized as reported earlier (16). DA female rats were immunized with spinal cord homogenate from allogenic healthy DA females matched by age in incomplete Freund's adjuvant in the tail base resulting in protracted, relapsing, and demyelinating EAE. This model is characterized by massive infiltration of blood-borne cells (macrophages, lymphocytes, and granulocytes) into the parenchyma leading to demyelination, axonal suffering, and expression of proinflammatory markers. Clinical disabilities are measured by a clinical score: 1, loss of tail tone; 2, weakness in one or both hind legs or middle ataxia; 3, ataxia or paralysis; 4, severe hind-leg paralysis. All experiments were performed in accordance with European Union (86/609/EEC) and French National Committee (87/848) recommendations (animal experimentation permission, France 33/00055). Immunized rats underwent magnetic resonance imaging (MRI) with intravenously injected ultra-small superparamagnetic iron oxide nanoparticles injection and T2-weighted MRI examination (17) at clinical onset, a protocol that was recently established as prediction marker of disease development, toward either severe relapsing (MRI+ve) or mild monophasic (MRI−ve) EAE (18). Rats were randomly grouped and sacrificed at four time points during EAE development: at disease onset (n = 8), during the first attack (n = 8), during the second attack or equivalent time frame (n = 11), and during recovery phase (n = 6). D3 expression was evaluated in inflammatory spinal cord lesions during these time points. Apart from immunized EAE animals, spinal cords of three vehicle-injected control rats were studied.
Tissue preparation and immunohistochemistry
After sacrifice and intracardiac perfusion with 4% paraformaldehyde, spinal cords were sectioned with a vibratome; free floating coronal sections (30 μm) were stained for D3 expression as described (14). Double labeling was performed by immunofluorescent assay. Subsequent incubations in 1/2000 polyclonal rabbit anti-D3 antiserum 676 (kindly provided by Prof. T.J. Visser; Erasmus MC, Rotterdam, The Netherlands) and 1/1000 mouse monoclonal antibody against rat ED1 antigen (Serotec, Düsseldorf, Germany) were followed by 1/1000 goat anti-rabbit CY3–conjugated antibody (Jackson ImmunoResearch, Newmarket, UK), 1/1000 goat anti-mouse Alexa® 488–conjugated antibody (Molecular Probes [Invitrogen] Cergy-Pontoise, France), and 4′,6-Diamidino-2-phenylindole (DAPI) staining. The analysis was done using light, epifluorescent (Nikon, Champigny sur Marne, France), or confocal microscopy (Leica, Rueil Malmaison, France).
Monocyte culture and activation
Activated macrophages can acquire a plethora of activation phenotypes resulting in either proinflammatory (M1 phenotype) or tissue protecting (M2 phenotypes) properties that are necessary for effective inflammatory reaction and tissue restoration during inflammation (19). To confirm D3 enzyme expression by activated monocyte/macrophages, circulating blood monocytes were tested in vitro under different activation conditions inducing either M1 proinflammatory or M2 immunomodulatory profile compared with nonstimulated M0 type (20,21). Blood was collected from healthy DA female rats by heart puncture, and circulating blood monocytes were isolated by standard Ficoll protocol and adhesion. About 106 Peripheral Blood Mononuclear Cell (PBMC) were plated out, and after 2 hours of culture, nonadherent cells were removed. Adherent cells (∼10%) were cultured for 20 hours in RPMI 1640 medium supplemented with 5% heat-inactivated fetal calf serum, 2% L-glutamine, and 1% penicillin/streptomycin solution (complete medium), in the presence of factors promoting activation toward M1 (10 μg/mL LPS and 20 ng/mL interferon γ) or M2 (20 ng/mL interleukin [IL]-10 and 20 ng/mL IL-13), phenotype or in complete medium alone (M0 activation). M1, M2, and M0 phenotypes were controlled by analysis of activation marker expression with quantitative real-time reverse transcriptase (RT)–polymerase chain reaction (BioRad Laboratories, Marnes-la-Coquette, France): inducible nitric oxide synthase (iNOS) and IL-1β (M1 phenotype) and mannose receptor (MR) and transforming growth factor β (TGFβ) (M2 phenotype). Primers used were GAPDH, 5′TGCTGGTGCTGAGTATGTCGTG and 5′CGGAGATGATGACCCTTTTGG; IL-1β, 5′AATGACCTGTTCTTTGAGGCTGAC and 5′CGAGATGCTGCTGTGAGATTTGAAG; iNOS, 5′GATTTTTCACGACACCCTTCACC and 5′GGTCCTCTGGTCAAACTCTTGGAG; MR, 5′AAGATCTCGTGGGCAACATC and 5′AAAAGACAGGCCAATGCAAC; and TGFβ, 5′CCGCAACAACGCAATCTATG and 5′AGCCCTGTATTCCGTCTCCTT. M1, M2, and M0 cells were fixed with 4% paraformaldehyde and double stained for D3 and ED1 expression with immunofluorescent labeling as described above. Increase of D3 staining intensity in M1 and M2 compared with nonactivated M0 cells was quantified in arbitrary units of immunostaining using ImageJ software. The intensity of staining was normalized in time independently for each marker, and background staining was taken into account as well. We considered M0 nonactivated cells as controls, and the intensity of D3 staining of M1 and M2 cells was normalized to the mean staining value of these control cells.
Results
Both EAE and control rats showed regular patterns of dispersed physiologic D3 immunostaining in both gray and white matter in spinal cords. Figure 1a displays a representative positive D3 staining in the white matter tissue of a control rat, whereas Figure 1b displays intense D3 staining in infiltrating inflammatory cells around blood vessels in the white matter of an MRI+ve EAE rat. In contrast, MRI−ve rats with predicted and confirmed monophasic EAE showed hardly any D3+ve immunoreactivity mostly dispersed in the parenchyma, as did control rats. Incubation of slices with D3 preimmune serum did not result in any immunostaining (Fig. 1c).

Type 3 deiodinase (D3) immunostaining in inflamed experimental autoimmune encephalomyelitis (EAE) spinal cord lesions. (
The intensity of D3 staining in inflammatory infiltrates was scored to give a semiquantitative magnitude of the D3 expression. Table 1 represents D3 intensity in inflammatory infiltrate between 0 (absent) and 3 (very intense) in MRI−ve and MRI+ve rats. No D3+ve staining at infiltration sites was observed in MRI−ve rats, whereas intense D3 expression was present in MRI+ve rats. However, no quantitative expression differences were observed at different time points.
D3 staining was scored using the following grades. 0, no staining present; 1, hardly any staining; 2, clear staining; 3, strong staining.
D3, type 3 deiodinase; MRI, magnetic resonance imaging; SD, standard deviation.
We attempted to further characterize the D3+ve cells in the perivascular inflammation sites by confocal immunofluorescence analysis. As blood-borne macrophages represent a major cellular compound in EAE and MS lesions, we analyzed D3 expression in combination with the ED1 macrophage marker. Although ED1+ve macrophages were massively present in infiltrates, only a minority (less than 10% in perivascular infiltrates) expressed D3. Detailed analysis by confocal microscopy confirmed colabeling of D3+ve and ED1 +ve cells (Fig. 2a, b). Further, D3 +ED1−ve cells were identified as granulocytes by morphological analysis of cell nuclei (Fig. 2a, c).
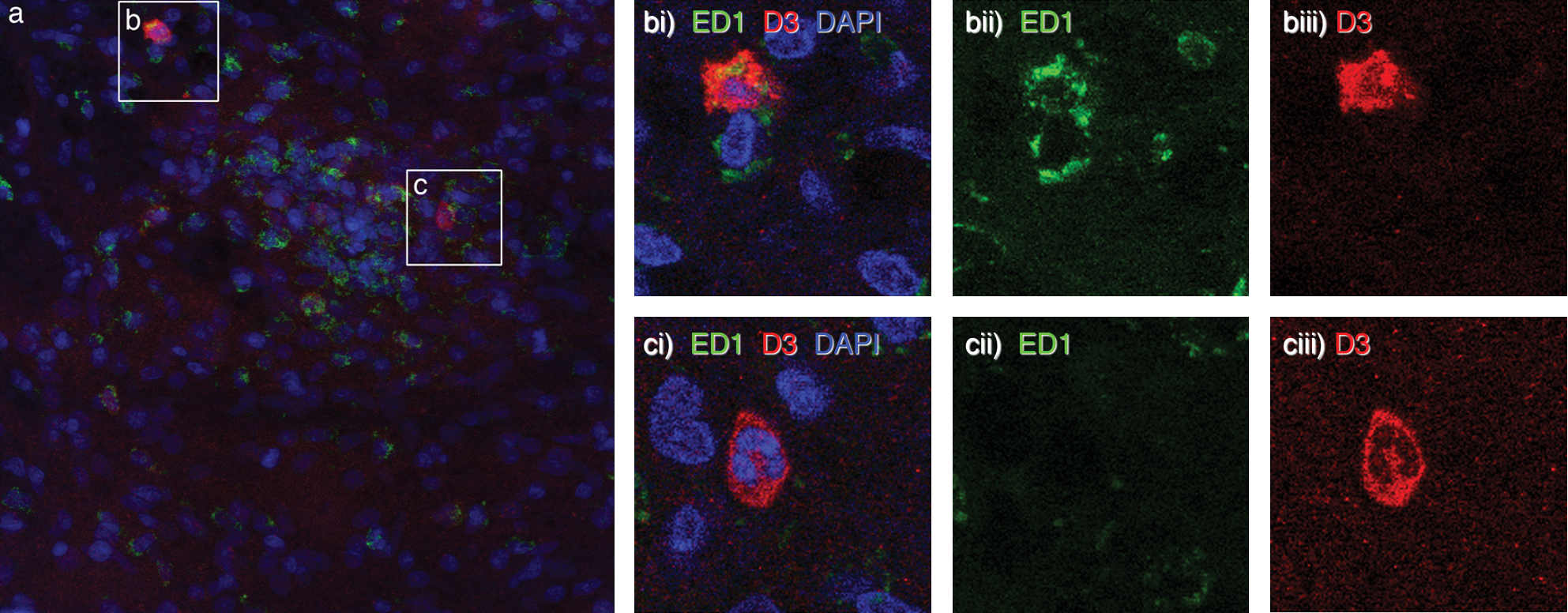
(a) D3+ve macrophages and granulocytes in perivascular infiltrates in spinal cord sections of EAE rat during relapse. Fluorescent triple staining for D3 protein (red), the macrophages/microglia marker ED1 (green) and nuclear labelling with DAPI dye (blue) revealed that a small population of ED1+ve cells expresses D3 enzyme (
Since we detected in inflammatory lesion D3+ expression in a subpopulation of macrophages, we further asked whether this phenomenon is preferentially associated to one of the activation types. Blood monocytes were in vitro activated toward M1 or M2 phenotypes. In agreement with M2 phenotype in MS lesions (22), preliminary data from our laboratory revealed both M1 and M2 activated macrophages besides M0 macrophages in CNS lesions (J. Mikita, N. Dubourdieu-Cassagno, M.S.A. Deloire, M. Biran, G. Raffard, B. Brochet, M.H. Canron, J.M. Franconi, C. Boiziau, K.G. Petry.). The activation state of cultured monocytes was analyzed by real-time RT–polymerase chain reaction, and expression of marker genes for M1 and M2 phenotypes was measured: iNOS and IL-1β (M1 phenotype) and MR and TGFβ (M2 phenotype). Compared with nonactivated controls, expression of iNOS and IL-1β in M1-activated cells increased or remained stable, respectively, as in M2-activated cells iNOS and IL-1β mRNA expression levels were diminished. On the contrary, expression levels of M2 marker genes (MR and TGFβ) were decreased in M1-activated monocytes and augmented in M2-activated cells (Fig. 3). Activated monocytes were stained by anti-ED1 antibody, whereas anti-D3 antibody revealed D3 +ve cells. Analysis of fluorescent staining intensity revealed D3 overexpression due to activation, as 80% of M1-activated (n = 214) and 95% of M2-activated (n = 308) cells increased D3 expression compared with control M0 cells (n = 219). However, no difference between M1 proinflammatory and M2 immunomodulating macrophages was observed (Fig. 4).
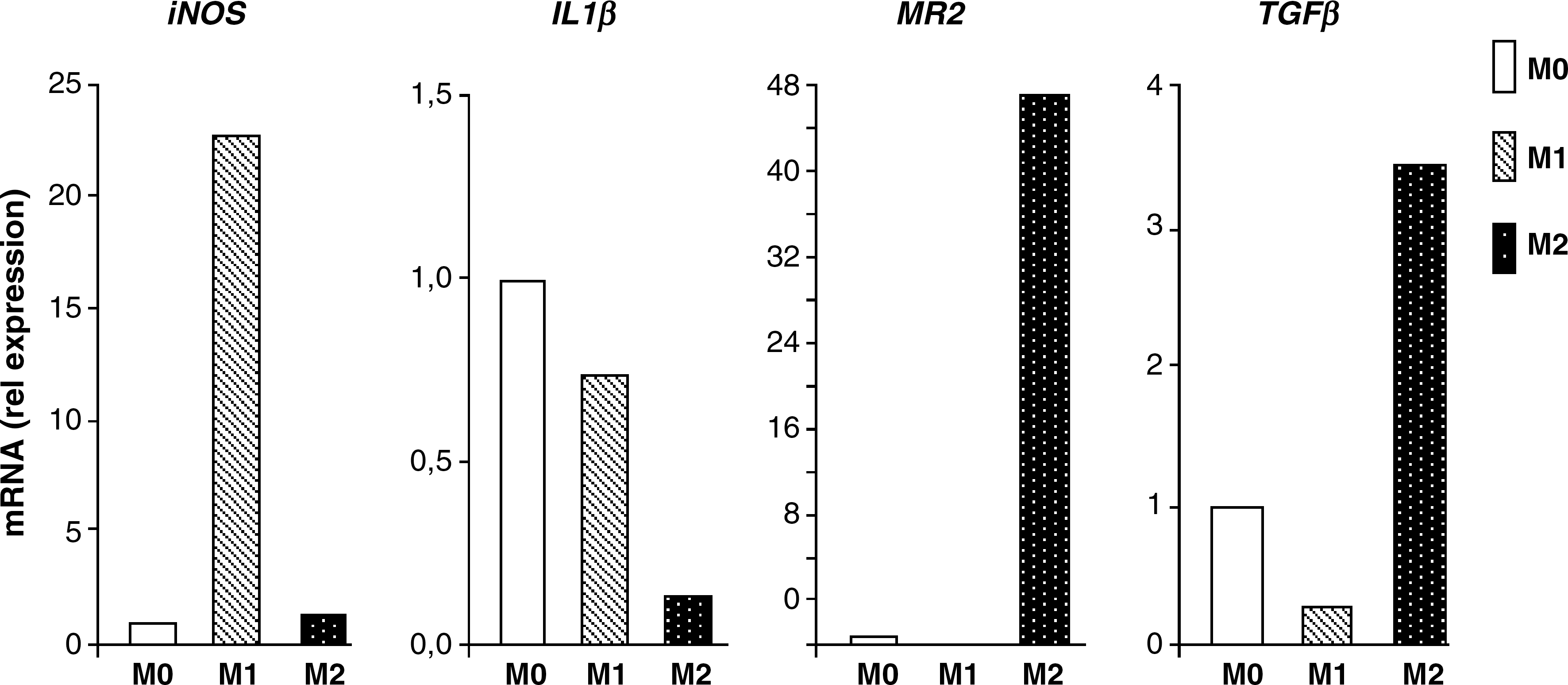
Profiles of activated monocytes expressing D3: blood monocytes were cultured in the presence of factors promoting activation toward M1 or M2 profile compared with nonactivated cells (M0). Expression levels of marker genes were quantified by real-time reverse transcriptase (RT)–polymerase chain reaction and normalized to reference gene expression level (GAPDH).
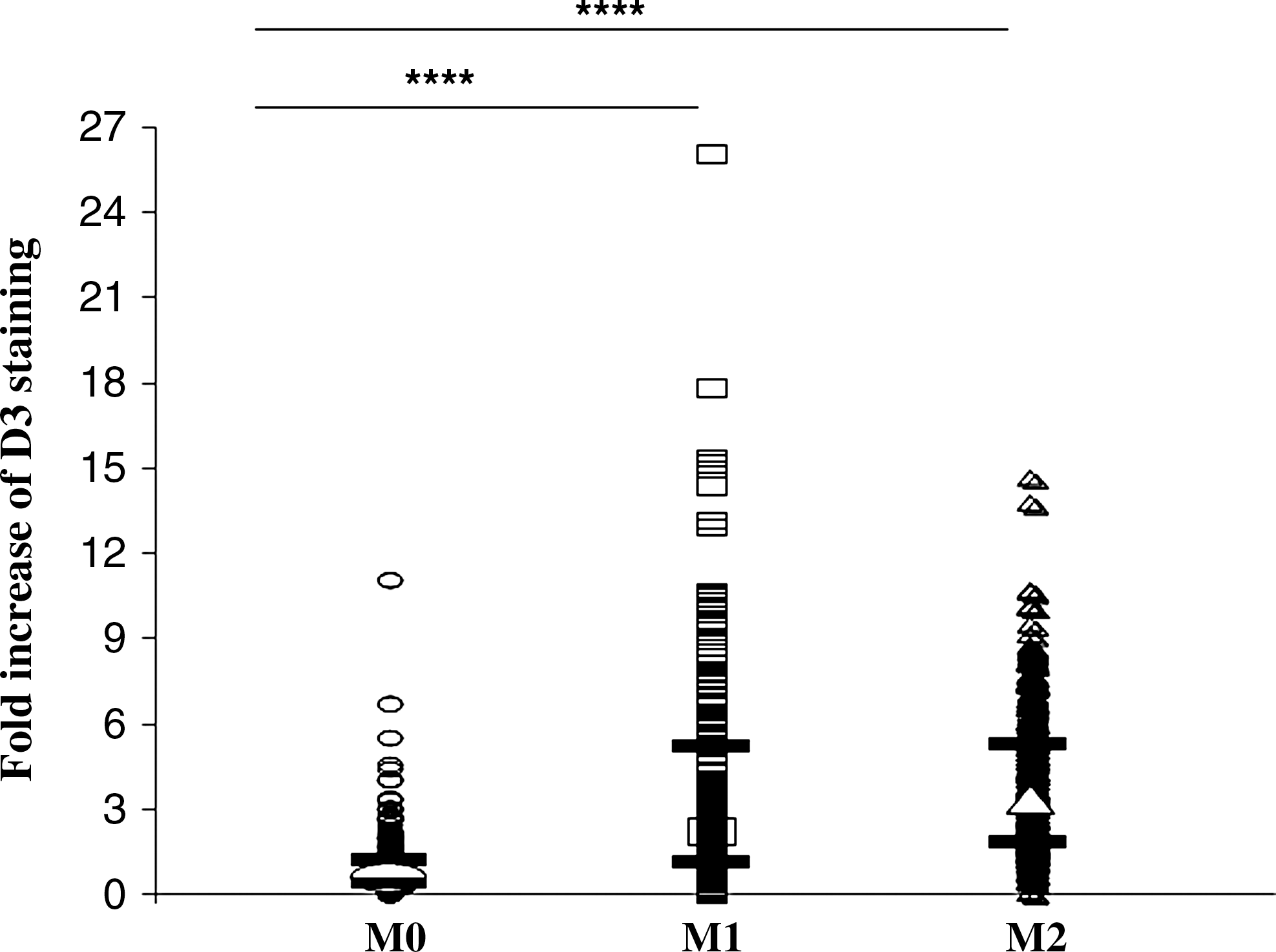
Fold increase of D3 expression (arbitrary units) in M1-activated proinflammatory (n = 214) and M2-activated immunomodulating rat monocytes (n = 308) compared with nonstimulated control monocytes (M0, n = 219). Median values (open circle, square, and triangle for M0, M1, and M2 cells, respectively) and first and third percentile values (black lines) are depicted; differences between groups were analyzed with Kruskall–Wallis (comparing three groups) or Mann–Whitney tests (comparing two groups) using Statview software. ****p < 0.0001.
Discussion
Our study of CNS inflammation demonstrates for the first time abundant D3 immunoreactivity in inflammatory spinal cord lesions of severely affected EAE rats. Strong demyelination in inflammatory lesions was observed earlier in this EAE model (10,16,18). Upon morphological and immunolabeling criteria, granulocytes and macrophages were, respectively, identified as cell types expressing D3. In vitro activation of monocytes confirmed D3 overexpression regardless of either proinflammatory or immunomodulatory activation. Considering proinflammatory and immunoprotective functions of M1 and M2 cells, respectively, D3 activity may thus be implicated in various pathophysiological mechanisms.
A major function of D3 in tissues is probably protecting them from excessive concentrations of thyroid hormones by inactivating them. In the human hypothalamic neurons, the main iodothyronine deiodinase expressed is D3 (23); it is also expressed in primary cultures of rat astrocytes (24). In addition to this homeostatic expression that we also observed in control rats, increased D3 expression was also observed in cells and tissues under pathological conditions of oxidative stress and ischemic hypoxia, suggesting regulation of metabolic tissue functions by decrease of T3-dependent oxygen consumption (25,26). In peripheral nervous system, sciatic nerve section was followed by prolonged D3 expression (27). In our experimental conditions, D3 expression by M2-activated monocytes in culture and by ED1+ve macrophages/microglia in inflammatory lesions might contribute to protective mechanisms active during tissue injury. Tissue injury and inflammation are associated with induction of a variety of growth factors, cytokines, and chemokines. A few in vitro studies describe the stimulating effects of epidermal and fibroblast growth factors on D3 mRNA expression and activity in primary cultures of astrocytes (28,29). These growth factors induce, via their cellular receptors, several intracellular downstream signaling cascades, one of them being the Raf/MEK/ERK signaling cascade (29). The MEK/ERK cascade is also activated by IL-1β (30) and GM-CSF (31). Because macrophages and granulocytes derive from the same progenitor cell, one could imagine that both cell types express D3 when appropriate stimulated.
D3 is also implicated in mechanisms of acute inflammatory response, which could explain its expression in in vitro stimulated M1 proinflammatory macrophages. Recently, we observed D3 expression in infiltrating granulocytes during chemical (14) and bacterial peripheral inflammation (15). D3 activity results in generation of iodide, thus providing a substrate necessary for appropriate function of the myeloperoxidase (MPO) system. MPO is one of microbial killing mechanisms observed in granulocytes, suggesting that D3 induction during acute inflammation might be an adaptative mechanism in inflammatory cells. Indeed, active MPO is present in monocytes, macrophages, and microglia (32). Its role in MS, however, remains to be established (33).
Thyroid hormones are known to be involved in remyelination, as they reduce EAE-associated axonal damage by stimulation of OPC differentiation and myelin production during acute and chronic EAE. Immediate remyelination activity was observed when T4 was administered during the acute inflammatory episode due to mobilization and extensive proliferation activity of OPCs (9 –11). In demyelination models of cuprizone-induced death of oligodendrocytes, T3 treatment resulted in marked improvement of remyelination, at least partly mediated by sonic hedgehog expression activation (11). Very recently, Fernandez et al. showed a prominent role of T3 in restoration of impaired oligodendrocyte generation observed in neurospheres derived form EAE rats (12). Increased D3 expression in tissues theoretically results in decreased T3 availability, which fits in the concept that administration of thyroid hormones improves efficient remyelination during EAE (34). It is, however, unknown at present whether reduced local T3 concentrations are causal in impaired remyelination as observed during EAE. Unfortunately, there are no specific D3 inhibitors available at this time to test hypotheses regarding the role of D3. Tissue examination of patients suffering from Devic's neuromyelitis optica pathology demonstrated profound demyelination typical for lesions with abundant granulocytes (35). Whether lack of remyelination in active MS lesions is associated with similar mechanisms and D3 expression by infiltrating blood-borne cells remains to be elucidated.
Footnotes
Acknowledgments
JM was a recipient of a Ph.D. fellowship of the French Minister of Technology and Research. This work was partially supported by ANR-TecScan 06/15 and CR Aquitane (to KGP).
Disclosure Statement
The authors declare that no competing financial interests exist.