
Abstract
Background:
Estrogen promotes the growth of thyroid cells. Therefore, we analyzed the influence of estrogen and selective estrogen receptor modulators (SERMs) on the expression of vascular endothelial growth factor (VEGF) and nitric oxide synthase III (NOS III) in the thyroid gland of ovariectomized (Ovx) rats.
Methods:
Wistar rats were divided into five groups, and bilateral ovariectomies were performed, except on the Sham-operated controls (Sham). Rats were grouped as follows: Sham; Ovx; and Ovx rats treated with daily subcutaneous injections of estradiol benzoate 3.5 μg/kg, tamoxifen 2.5 mg/kg, or raloxifene 2.5 mg/kg for 50 consecutive days. Control animals received vehicle (propyleneglycol), and at the end of the treatment, rats were sacrificed. The thyroid glands were excised, weighed, and processed for analysis of the expression of VEGF or NOS III by immunohistochemistry. The mean vascular areas were evaluated by immunodetection of α-smooth muscle actin.
Results:
Thyroid weight and mean vascular area were lower in Ovx as compared with Sham, Ovx + estradiol benzoate, Ovx + Tam, or Ovx + Ral (p < 0.01). VEGF (p < 0.01) and NOS III expressions (p < 0.05) were significantly lower in the Ovx group, as compared with Sham, Ovx + estradiol benzoate, Ovx + Tam, and Ovx + Ral. Immunoreactivity for both VEGF and NOS III was mainly detected in the cytoplasm of the follicular epithelial cells.
Conclusions:
Our data suggest that estrogen and SERMs regulate the thyroid gland vascularization and that tamoxifen and raloxifene behave like estrogen does. Estrogen and SERMs upregulate VEGF and NOS III in such a way as to reverse the effects detected on the thyroid microvasculature of the Ovx rats.
Introduction
Selective estrogen receptor modulators (SERMs) are therapeutic agents currently administered to a large number of patients. These compounds are characterized by showing either estrogen agonist or antagonist properties, depending on the specific target tissue. Tamoxifen represents the first generation of SERMs and raloxifene corresponds to a second generation of such compounds (11,12). Tamoxifen is a triphenylethylene derivative widely used for more than three decades to treat both early and advanced breast cancer (13). The efficacy of tamoxifen in preventing recurrence and fatalities from breast cancer when used as an adjuvant treatment has been well established (14). Raloxifene is a benzothiophene, commonly prescribed to postmenopausal women for prevention or treatment of osteoporosis (15). Also, as a prophylactic agent, raloxifene has been shown to be as effective as tamoxifen in reducing the risk of invasive breast cancer (16). SERMs may produce effects on reproductive and nonreproductive peripheral tissue sites (17,18). Tamoxifen is known to be an antagonist of estrogen in breast tissue and a partial estrogen agonist in uterus (11,19). On the other hand, raloxifene behaves as a complete antagonist not only in breast but also in uterus (20,21). Therapy with tamoxifen and raloxifene in postmenopausal women is associated with decreases in both cholesterol levels and bone loss, through their estrogen agonistic activities (22 –24). Additionally, tamoxifen and raloxifene exhibit vascular relaxing properties and behave as estrogen agonists in several vascular beds, inducing vasodilatation (25 –27).
Given the widespread role of estrogen in physiology, it is not surprising that estrogen may also be implicated in other unknown events. However, several aspects of the effects of SERMs on the nonreproductive tissues have not been elucidated, and data on the long-term efficacy and safety are limited (21,28). Biological effects of estrogen and SERMs are mediated by estrogen receptors that are present at organs susceptible to these compounds (1,2,29,30). It is well established that the thyroid gland is influenced by estrogen and SERMs (31 –33); however, the mechanisms underlying the thyroid gland changes by estrogen have not been studied so far. The purpose of this study was to test the hypothesis that these compounds might influence the thyroid gland vascularization. We analyzed the expression of VEGF and NO synthase III (NOS III) and correlated with the microvascular changes in the thyroid gland of Ovx rats treated with estrogen, tamoxifen, or raloxifene.
Methods
This investigation conforms to the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (Publication No. 85-23, revised 1996) and was approved by the Institutional Animal Welfare Committee. Adult female Wistar rats, 2 months old, weighing ∼200 g were kept under a 14:10 hours light:dark cycle (lights on at 08:00 am) at 23°C. The animals had ad libitum access to a standard diet and tap water. The animals were randomly divided into five treatment groups, and bilateral ovariectomies were performed, except on the Sham-operated controls (Sham). The rats were grouped as follows: Sham; Ovx; Ovx rats treated with estradiol benzoate (EB = Ovx + estrogen, 3.5 μg/kg body weight; Sigma, St. Louis, MO); Ovx rats treated with tamoxifen (Tam = Ovx + tamoxifen, 2.5 mg/kg body weight; Sigma); and Ovx rats treated with raloxifene (Ral = Ovx + raloxifene, 2.5 mg/kg body weight; Sigma). The dose of EB used corresponds to an intermediate dose that normally restores the physiologic serum estradiol levels (34 –37), and the doses of tamoxifen and raloxifene have been previously used in some studies (20,34 –39). We have chosen to use subcutaneous injections to be sure about the exact doses administered and delivered to tissues. Treatment began a month after ovariectomy, and the two control groups received vehicle (propyleneglycol). All animals were treated daily by subcutaneous injections for 50 consecutive days. Vaginal smears were daily collected from the sham-operated rats with intact ovaries to identify the phase of estrous cycle.
Tissue collection
At the end of the treatment, rats were weighed and sacrificed by decapitation. Sham were killed only during the estrous phase. Uteri were removed, drained of the luminal fluid, freed from connective and adipose tissues, and weighed to confirm the effects of ovariectomy or drug treatments. The thyroid glands were carefully dissected out and immediately fixed in 10% formalin. Every fixed gland was then dehydrated in graded alcohol, embedded in paraffin, sectioned at 4 μm, and mounted on the glass slides. Sections of the central portion of thyroid, known to be representative of the whole gland, were selected for analysis (40).
Immunohistochemistry
Tissue sections were initially dewaxed and rehydrated by serial passages through xylene and graded alcohols. To block endogenous peroxidase activity, they were placed in 3% H2O2, and to reduce nonspecific protein binding, they were incubated in phosphate-buffered saline supplemented with 5% bovine serum albumin for 15 minutes. Then the slides were incubated with primary antibodies for VEGF at a dilution of 1:200 (monoclonal antibody; Santa Cruz Biotechnology, Santa Cruz, CA), NOS III at a dilution of 1:500 (monoclonal antibody; Santa Cruz Biotechnology), or α-smooth muscle actin (α-SMA) at a dilution of 1:400 (polyclonal antibody; Dako, Carpinteria, CA) overnight at room temperature in humidified chamber. After that the samples were rinsed in phosphate-buffered saline and incubated with biotinylated link antibody and peroxidase-labeled streptavidin, according to the product datasheet instructions (LSAB kit; Dako). The peroxidase activity was revealed by 3-3′diaminobenzidine (Dako). Sections were counterstained with Mayer's hematoxylin, rinsed, and then mounted. For negative controls, incubation of the primary antibody was omitted.
Image analysis
The analyses were performed using digitalized images obtained directly from the light microscope via a video camera and stored in magnetic media. Three sections of the central portion of every thyroid gland were selected for analysis. From each section, three photomicrographs of 1280 × 1024 pixels were obtained from noncoincidental randomly selected fields at a magnification of ×400. Two different observers independently evaluated the images, with similar results. Both of them were blind to the analyzed groups. The images were evaluated using the imaging processing and analysis system Image Pro Plus 4.5.1.29 (Media Cybernetics, Silver Spring, MD,
Statistical analysis
Data are reported as mean ± standard error of mean. Statistical analysis was performed by the one-way analysis of variance followed by the Newman–Keuls multiple comparison test using the software GraphPad Prism version 4.00 for Windows (GraphPad Software, San Diego, CA,
Results
Uterine weight in Sham (456 ± 30.3 mg) was greater as compared to the other groups (p < 0.001). Treatment with estrogen increased uterine weight to 50% of Sham levels (243 ± 12.07 mg), which was significantly greater than in the Tam, Ral, and Ovx groups (p < 0.001). Uterine weight in tamoxifen-treated rats (92 ± 2.3 mg) was enhanced when compared with Ral and Ovx (p < 0.05), but not with EB and Sham groups (p < 0.001). Tam uterine weight corresponded to 20% of the Sham levels. Ral (44 ± 2.5 mg) and Ovx (34.7 ± 3.6 mg) uterine weights represent 10% and 8% of Sham levels, respectively (p < 0.001), and no significant differences in uterine weight were noted between these two groups (Fig. 1). Thyroid weight was lower in Ovx (12 ± 0.6 mg) as compared with Sham (15 ± 0.6 mg), EB (15 ± 0.7 mg), Tam (15 ± 0.3 mg), or Ral (15 ± 0.6 mg) (p < 0.01).

Uterine weight (mg) in Sham, Ovx rats that received vehicle or were treated with estrogen (EB), tamoxifen (Tam), or raloxifene (Ral). Columns with different letter designations correspond to the significantly different means (p < 0.05). Data are expressed as mean ± standard error of mean. Sham, Sham-operated controls; Ovx, ovariectomized.
Immunoreactivity for both VEGF and NOS III was mainly localized to the cytoplasm of the follicular epithelial cell, and extracellular, in the thyroid stroma tissue, in association with the extracellular matrix (Figs. 2 and 3). VEGF staining was also detected in smooth muscle cells of blood vessels. Endothelial cells were negative for both VEGF and NOS III. The PAI of VEGF in the thyroid stroma varied from 0.20% to 5.80% in different groups. VEGF was less expressed in the Ovx group (0.20 ± 0.01%), as compared with Sham (3.86 ± 0.90%), EB (5.32 ± 0.81%), Tam (4.27 ± 0.67%), and Ral (5.80 ± 1.42%) (p < 0.01) (Fig. 4A). The PAI of NOS III varied from 0.22% to 5.87% in different groups and was also present in the thyroid stroma tissue. Immunoexpression of NOS III in Ovx (0.22 ± 0.04%) was lower than in Sham (3.03 ± 0.97%), EB (5.34 ± 0.44%), Tam (4.97 ± 1.44%), and Ral (5.87 ± 0.28%) (p < 0.05) (Fig. 4B). Mean vascular area varied from 0.33% to 3.80% in different groups (Fig. 4C). Ovx rats showed decreased vascular tissue area (0.33 ± 0.07%) in comparison to Sham (3.30 ± 0.88%), EB (3.80 ± 0.51%), Tam (3.59 ± 0.22%), and Ral (2.44 ± 0.33%) (p < 0.01).
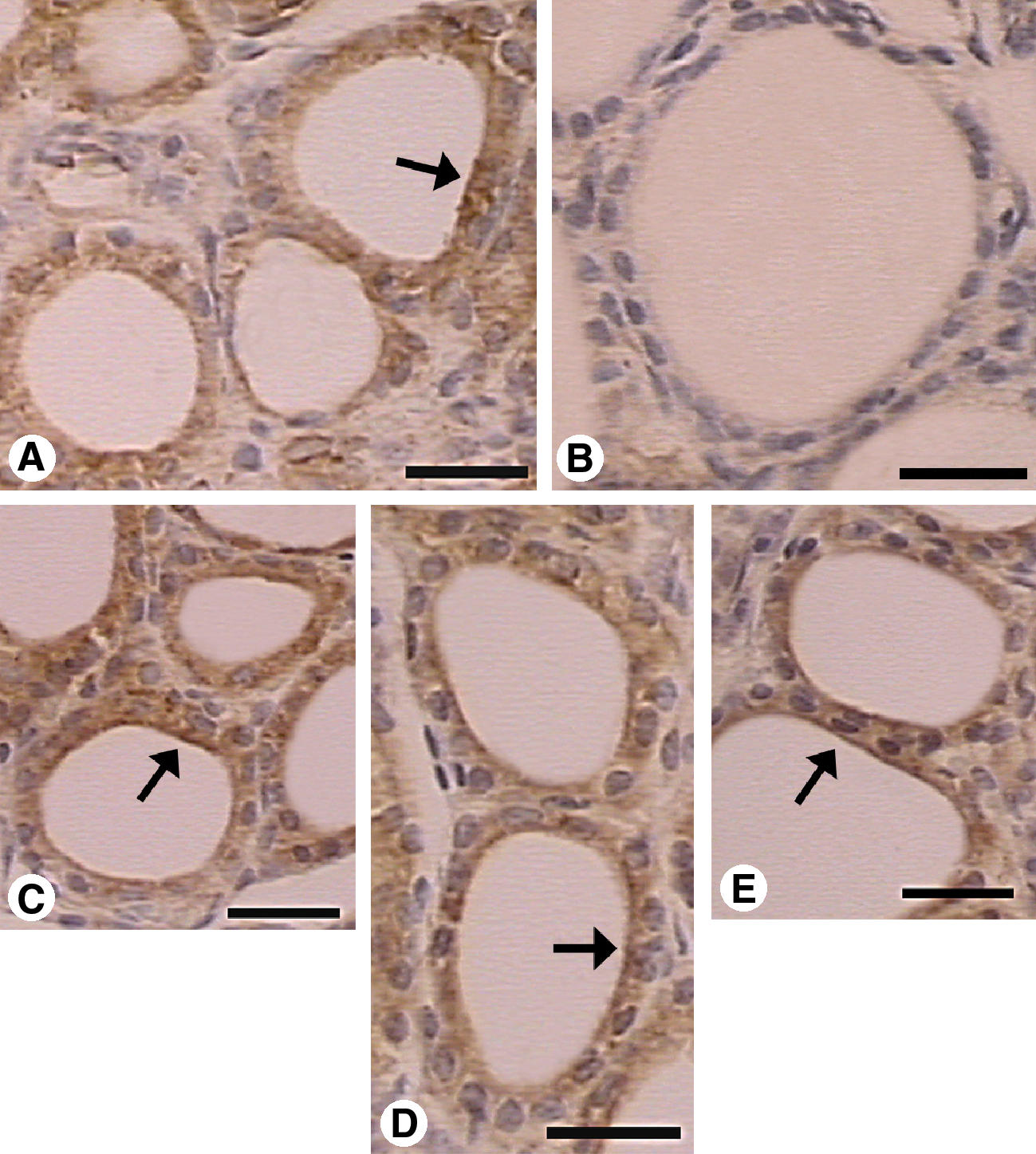
Immunohistochemical expression of VEGF in the thyroid gland of Sham and Ovx rats treated or not with estrogen, tamoxifen, and raloxifene (scale bars, 25 μm). Immunostaining for VEGF was detected in all experimental groups. In Sham (
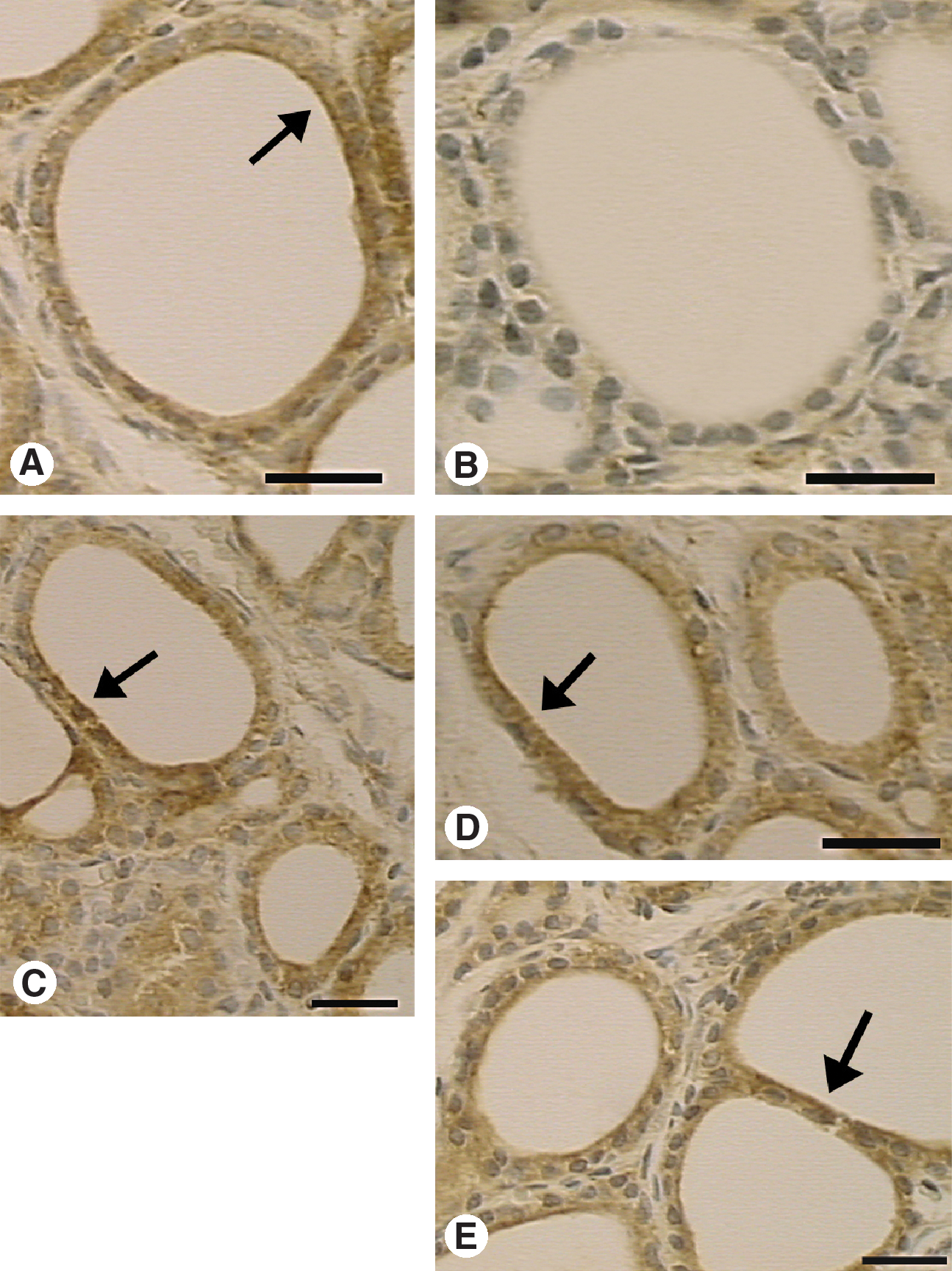
Immunohistochemical expression of NOS III in the thyroid gland of Sham and Ovx rats treated or not with estrogen, tamoxifen, and raloxifene (scale bars, 25 μm). Immunostaining of NOS III was detected in all experimental groups. In Sham (

Percentual area of immunoexpression of VEGF (
Discussion
Mammalian thyroid glands consist of clustered follicular units surrounded by a closely adjacent microvascular bed. Vascular supply is a requirement for cellular inflow of nutrients and provides a pathway for specific glandular secretory products. Modifications in vasculature generally represent a critical process not only in pathogenesis but also in normal thyroid conditions, because adequate blood supply is essential for metabolic and endocrine functions (43,44). Thus, the development of follicular microvasculature may be an important prerequisite for sustaining the thyroid function status, as well as nodules that might develop (45). However, mechanisms mediating the thyroid microvascular changes are not well understood. It has been well established that each follicular unit has its own local vascular regulatory system, and each one of them is responsible to regulate its own capillary network, according to the individual functional status (46). Previous studies provided evidence that thyrocytes produce a number of growth and vasoactive factors. Therefore, they may regulate changes in the thyroid gland microvasculature and blood flow through paracrine exchanges with their neighbor capillary bed (47). This phenomenon involves complex regulatory networks in which local secretion of VEGF and NO is clearly involved (44,47).
Our data suggest that estrogen influences the thyroid gland vasculature. Regressive changes in the thyroid gland microvasculature were found as a consequence of steroid hormone depletion after ovariectomy, because estradiol or SERM treatment reversed these effects. The mean vascular area in the thyroid gland was decreased in Ovx rats that received only vehicle, as compared with Sham (intact ovaries) and Ovx rats treated with estrogen, tamoxifen, and raloxifene. Also thyroid weight was decreased in Ovx compared with EB, Tam, Ral, and Sham rats. These findings are in accordance with the previous studies (48) and may occur as a consequence of the reduced blood flow in the thyroid gland of Ovx animals.
In fact, estrogen deprivation is associated with the development of long-term modifications in target organs in females (49). Previous reports postulated that morphological and functional status of the thyroid gland in women may be affected by aging, in correlation with the progressive changes in the expression of functional proteins, such as VEGF and NOS III (50). Experimental studies provided evidence that aging (51) and ovariectomy (52) may induce morphological changes in the thyroid gland in rodents, compatible with the hypofunctioning status in correlation with the decreases in the thyroid function profiles.
In our experiments, VEGF and NOS III were overexpressed in the thyroid gland of drug-treated Ovx animals, when compared with Ovx. VEGF and NOS III staining was specially localized in follicular cells, and the presence of estrogen receptors in the thyroid gland suggests a direct action of estrogen on thyrocytes (31,33,53). Indeed, previous studies have demonstrated a definite role of estrogen not only in thyroid pathogenesis and in thyrocytes proliferation but also in the normal development and growth of the thyroid gland (32,54,55). Experimental data have already suggested that sex steroids act directly on thyrocytes to stimulate cell proliferation and thyroid growth (33,52). In addition, evidence for a direct stimulatory effect of estradiol on the thyroid cell has been reported in Fischer rat-derived thyroid cell line (FRTL-5) rat thyroid cell line that expresses functional estrogen receptors (56).
Our data corroborate the concept that thyrocytes play a critical role in the regulation of thyroid microvasculature because staining for VEGF and NOS III was localized in follicular epithelium. In addition, these findings suggest a paracrine role for VEGF and NO in microvascular regulatory system, because VEGF and NOS III overexpressions were related to the increases in the thyroid microvascular area. Also, our results indicate that VEGF and NOS III actions in the thyroid gland are coordinated. In fact, we analyzed the microvascular area in different groups through immunostaining for α-SMA, and antibodies for α-SMA are known to label reactive vessels (arterioles).
Further, according to our results, VEGF and NO may be considered major factors in the regulation of thyroid vasculature. VEGF is a polypeptide specially known as a proangiogenic factor, with a central role in function, development, and growth of blood vessels (57). Interestingly, this cytokine may exhibit other regulatory functions. Experimental data suggested that VEGF is synthesized and secreted by normal thyroid epithelium (58) and may contribute to the regulation of development and function of follicular cells through autocrine and paracrine effects (59). There is evidence that the function of other paracrine factors may depend on VEGF expression in follicular cells (57). It has been reported that thyrocytes produce VEGF mainly in response to the thyroid-stimulating hormone (TSH), among other signals (60,61). Our findings show that estrogen is another signal that regulates the synthesis of VEGF in the thyroid gland. Experimental evidence suggests that the effects of TSH on VEGF production by the thyroid cells may be variable in relation to different experimental conditions (62). Whether or not estrogen modulates thyrocyte response to TSH stimulus for the synthesis of VEGF remains to be clarified. In the thyroid gland, the widespread distribution of VEGF in normal tissues corroborates the idea that it is primarily a regulator of normal function, rather than a mediator of pathological processes (57). In fact, VEGF shows vascular permeability-enhancing properties, and it is known to induce enhanced vascular permeability in different conditions (63). As an endocrine organ, the thyroid gland is richly vascularized with a fenestrated epithelium lining the blood vessels, which may under adequate conditions allow transfer of substances across the permeable vessel wall (64). Therefore, one of the physiological roles of VEGF in the thyroid gland may be to regulate the vascular permeability and facilitate the action of other paracrine factors. Likewise, VEGF-binding sites are consistently found in association with the vasculature of these areas (65). These findings raise the possibility that the tonic presence of the growth factor may be required to maintain the differentiated state of those vessels, which otherwise might undergo involution. It is therefore tempting to speculate that the suppressed expression of VEGF may be responsible, at least partially, for the regressive phenomenon that is detected in the thyroid of Ovx rats (57).
NO is known to act as a vasodilator and may influence vascular tone and blood flow in different tissues. Immunodetection of NOS III, one of the three NOS isoforms, is a commonly used method to evaluate synthesis of NO. Although it has initially been described that NOS III is expressed exclusively in endothelial cells, it is now clearly established that the enzyme is present in many different cell types (66). In this study, we found NOS III expression in follicular cells, and this finding is in agreement with the previous investigations that have shown the expression of NOS III in the normal and pathological thyroid gland (44,46).
In our study, the findings for uterine weight are consistent with the previously described experiments and therefore confirmed the efficacy of ovariectomy and drug treatments (11,20). Moreover, we demonstrated that estrogen as well as tamoxifen and raloxifene affect the thyroid gland microvasculature, and no significant differences were found between the treated Ovx rats and the control groups. These findings are compatible with an estrogen agonistic action of tamoxifen and raloxifene on the thyroid gland. Obviously, the extrapolation of these results to humans must be carefully interpreted. It is reasonable that other experimental studies in animals should be developed, allowing a better understanding of the complex regulatory system of the thyroid gland microvasculature. Further investigations are also necessary to elucidate possible implications of therapy with estrogen and SERMs on physiological or pathological processes of the thyroid gland.
Footnotes
Disclosure Statement
The authors declare that no competing financial interests exist.