
Abstract
Background:
Tetraiodothyroacetic acid (tetrac) is a deaminated analogue of L-thyroxine that blocks the actions of L-thyroxine and 3,5,3′-triiodo-L-thyronine at the cell surface receptor for thyroid hormone on integrin αvβ3. Tetrac blocks the proliferative effects of thyroid hormone on tumor cells and the proangiogenesis actions of the hormone. In the absence of thyroid hormone, tetrac also blocks angiogenesis induced by various growth factors. Covalently linked to poly(lactide-co-glycolide), tetrac nanoparticles (tetrac NP) do not gain access to the cell interior and act exclusively at the integrin receptor. Here, the activity of tetrac and tetrac NP against follicular thyroid carcinoma (FTC)-236 cells was studied in two models: (1) tumor cell implants in the chick chorioallantoic membrane (CAM) system and (2) xenografts in the nude mouse.
Methods:
FTC-236 cells (106) were implanted in the CAM (n = 8 each for control, and for tetrac and tetrac NP, both at 1 μg/CAM) and the actions of tetrac and tetrac NP were determined after 8 days on tumor-related angiogenesis and tumor growth. Xenografts of 107 FTC-236 cells were implanted in nude mice (n = 8 per group). Tetrac or tetrac NP was administered intraperitoneal (1 mg/kg and 1 mg tetrac equivalent/kg, respectively) every other day for 32 days beginning on day 10, when tumor volume was 200–250 mm3. Animals were monitored after discontinuation of treatment up to day 40.
Results:
In the CAM paradigm, tetrac and tetrac NP arrested tumor-related angiogenesis and tumor growth. In the xenograft model, tetrac and tetrac NP promptly and progressively reduced tumor volume (p < 0.01) over 32 days. There was some regrowth of tumor after interruption of tetrac treatment, but at day 40, tumor volume and tumor weight at sacrifice were 45–55% below those of controls (p < 0.01). Animal weight gain was comparable in the control and treatment groups of animals.
Conclusions:
Tetrac and tetrac NP effectively arrest FTC-236 cell tumor growth in the CAM and xenograft models, suggesting its potential utility against FTC.
Introduction
Recent in vitro studies from our laboratories showed that thyroid hormone (L-thyroxine [T4]) or 3,5,3′-triiodo-L-thyronine treatment of human papillary carcinoma and FTC cell lines resulted in enhanced cell proliferation, measured by proliferating cell nuclear antigen. Thyroid hormone also induced activation of the Ras/MAPK (ERK1/2) signal transduction pathway. ERK1/2 activation and cell proliferation caused by thyroid hormone were blocked by an iodothyronine analogue, tetraiodothyroacetic acid (tetrac), which inhibits binding of iodothyronines to the cell surface receptor for thyroid hormone (TR) on integrin αvβ3 (2,3). A MAPK cascade inhibitor at MEK, PD 98059, also blocked hormone-induced cell proliferation. We then assessed the possibility that thyroid hormone is antiapoptotic. Thus, plasma membrane-initiated activation of the MAPK cascade by thyroid hormone promotes papillary carcinoma and FTC cell proliferation and is blocked by tetrac (2).
The aim of this investigation was to test the in vivo antitumor efficacy of the thyroid antagonist tetrac and tetrac immobilized on the surface of poly(lactic-co-glycolic acid) (PLGA) nanoparticles. The thyroid hormone signal at the cell surface receptor is transduced into an angiogenic response (4,5) in endothelial cells and vascular smooth muscle cells that bear the receptor, as well as into a tumor-cell proliferative response in cancer cells (1,6). The proliferative response is generated by L-T4 in physiological concentrations and by 3,5,3′-triiodo-L-thyronine in concentrations that exceed the physiologic (3). The deaminated T4 analogue, tetrac, is an antagonist at the integrin receptor (3), rather than an agonist, and blocks the binding of agonist thyroid hormone analogues to the integrin receptor. Within the cell, however, tetrac can have low-potency thyromimetic activity (7). To limit the action of tetrac to the TR, we synthesized tetrac nanoparticles (tetrac NP), which are excluded from the cell interior. The nanoparticle used in this study is tetrac bound at the outer ring hydroxyl by an ether linkage to PLGA. The average length of the nanoparticle is 200 nm.
In addition to its inhibition of the cell surface effects on cell proliferation of thyroid hormone, tetrac is antiangiogenic in the absence of thyroid hormone (8). The thyroid hormone-tetrac receptor is located at the Arg-Gly-Asp (RGD) recognition site on integrin αvβ3 (3,9), which is important for interactions of the integrin with extracellular matrix proteins and growth factors (10). There is evidence for clustering of and crosstalk between integrin and vascular endothelial growth factor receptors (11) and basic fibroblast growth factor receptors (12). Vascular endothelial growth factor and basic fibroblast growth factor are proangiogenic agents whose activity is blocked by RGD peptides and tetrac (8).
In a human breast cancer xenograft model in the nude mouse, tetrac has been shown to inhibit tumor growth (13), presumably by its antiproliferative action. Its antiangiogenic action was not explored in previous studies. In this study, we examined the effects of unmodified tetrac and tetrac NP on the growth of human FTC xenografts and also estimated the vascularity of the treated tumors.
Materials and Methods
Materials
Polyvinyl alcohol, N-(3-dimethylaminopropyl)-N′-ethylcarbodiimide-hydrochloride (EDC), and the dialysis tubing cellulose membrane were purchased from Sigma Aldrich (St. Louis, MO). Poly(
Methods
PLGA nanoparticles were synthesized by modification of a method (single-emulsion solvent diffusion) originally described by Jeffery et al. (14) and Song et al. (15). Amino-functionalized PLGA nanoparticles were obtained by conjugating the PLGA nanoparticles with ethylenediamine, using carbadiimide chemistry. Amino-functionalized PLGA nanoparticles were reacted with tetrac intermediate, supplied by Azopharma Contract Pharmaceutical Services, to obtain tetrac-conjugated PLGA nanoparticles. The custom-made intermediate of tetrac was composed of tetrac conjugated to epibromohydrin through the phenolic-OH group present on it. This epoxy group reacts with the amino group (16,17) present in the modified PLGA nanoparticles in aqueous conditions. In a typical experiment, PLGA nanoparticles were synthesized by adding 200 μL of PLGA (40 mg/mL in dimethyl sulphoxide) to 20 mL of a 1% aqueous solution of polyvinyl alcohol. This mixture was constantly stirred for about 12 h at room temperature to form the nanoparticles. The nanoparticle suspension was then dialyzed for 6 h using a dialysis membrane with a molecular cutoff of 10–12 kDa to remove the impurities as well as the organic solvent. To 18 mL of this void PLGA nanoparticle solution were added 2 mL of phosphate-buffered saline (PBS) (10 ×) and 500 μL of EDC (180 mg/mL in 10 × PBS). The mixture was stirred for about 1 h, followed by another addition of 500 μL of ethylenediamine (280 mg/mL in 10 × PBS) and then magnetic stirring was continued for at least 24 h. The whole solution was then dialyzed (10–12 kDa cutoff) for 10–12 h to eliminate unreacted materials. A stock solution of the custom-made epoxy-activated tetrac (Azopharma Contract Pharmaceutical Services) in anhydrous dimethyl sulphoxide (5 mg/mL) was prepared, and then 100 μL of this activated tetrac solution was added to 10 mL of the above amino-functionalized PLGA nanoparticles and stirred for at least 24 h. The solution was then dialyzed for at least 12 h for purification (12 kDa cutoff) and lyophilized. These lyophilized tetrac-conjugated PLGA nanoparticles were reconstituted in deionized water/PBS and used for the experiments described below.
Cells and cell culture
Human FTC cells (FTC-236) were kindly provided to the authors (F.B.D., P.J.D.) by Dr. Orlo H. Clark (University of California, San Francisco, CA) and cultured as instructed by Dr. Clark, using a complete growth medium consisting of F-12K medium supplemented with 10% fetal bovine serum. The cells were cultured in 5% CO2 air atmosphere at 37°C to subconfluence and then treated with 0.25% (w/v) trypsin/ethylenediaminetetraacetic acid to affect cell release from the culture vessel. After washing with culture medium, the cells were suspended in DMEM (free of phenol red and fetal calf serum) and counted.
Cellular distribution of tetrac versus tetrac NP in FTC-236 cells
FTC cells were grown in RPMI 1640 and F-12K medium (1:1 ratio) (Invitrogen, Grand Island, NY), plus 10 mU/mL of thyrotropin (Sigma Aldrich), supplemented with 10% fetal calf serum (Atlanta Biologicals, Lawrenceville, GA). Penicillin and streptomycin (1%) were present in the culture media. The cells were trypsinized and centrifuged, and the cell pellet was resuspended in the appropriate media. Then 500 μL of the suspension (∼50,000 cells) were transferred to each well of a four-well glass chamber slide system (Nalge Nunc International, Naperville, IL) and incubated for 24 h at 37°C and 5% CO2 (Forma Series II; Thermo Electron, Waltham, MA). The cells were treated with 20 μL of each free tetrac tagged with Cy3 dye or tetrac-NP tagged with Cy3 and separately incubated (37°C, 5% CO2) for about 2 h. After 2 h of incubation, the plates were taken out and washed several times with PBS and then fixed in 1% formaldehyde (Sigma) and mounted using Vectashield (Vector Laboratories, Burlingame, CA). Confocal images were taken using a Leica TCS SP5 confocal microscope with a 63 × (NA = 1.3 glycerol immersion) objective. A 405 nm laser was used for excitation, and emission was detected between 565 and 688 nm.
Tumor growth in the chorioallantoic membrane cancer implant model
The effect of tetrac versus tetrac NP at 1.0 μg/chorioallantoic membrane (CAM) on tumor angiogenesis (total hemoglobin in tumor matrigel plug) and tumor growth (tumor weight) of 1 × 106 FTC cells implanted in Matrigel in 7-day-old chick eggs 8 days after implantation was determined (18). Matrigel (BD Bioscience, San Jose, CA) was thawed overnight at 4°C and placed on ice. FTC cells in exponential growth phase were harvested using 0.25% trypsin–ethylenediaminetetraacetic acid, washed, and suspended in medium. Only suspensions of single cells with a viability exceeding 95% were used. Approximately 1 × 106 cells in 30 μL of medium were mixed with the same volume (30 μL) of Matrigel and implanted on the CAM. Data represent mean tumor weights (mg) per treatment group and tumor hemoglobin (mg/dL) + standard error of the mean (n = 8 per group).
Cell implantation in nude mice
Female NCr nude homozygous mice aged 5–6 weeks with body weights of 20 g were purchased from Taconic Farms (Hudson, NY). Animals were maintained under specific pathogen-free conditions and housed four animals per cage, under controlled conditions of temperature (20–24°C) and humidity (60–70%) and with a 12 h light/dark cycle. Water and food were provided ad libitum. Xenograft experiments were carried out in the animal research facility of the Veterans Affairs (VA) Medical Center (Albany, NY) and the experimental protocol was approved by the VA-IACUC. Mice were allowed to acclimate for 5 days before the start of treatments. Subcutaneous implant of 1 × 107 FTC cells was carried out in the flank region of each mouse.
Treatment of animals with unmodified or tetrac NP
Tumors were measured daily by calipers and tumor volume was calculated according to a standard formula: W × L 2/2, where W = width and L = length. Tumor volume measurements were followed up after the implant every other day for the first 4 weeks after inoculation. Tumor volumes were measurable at 3 days after implant where the treatment started on day 10. Mice with tumors of 200–250 mm3 in volume proximal to the injection site were randomized into three groups (n = 8): vehicle (PBS, pH 7.8) control, tetrac (1 mg/kg) group, and tetrac NP (1 mg/kg) group. Drugs were administered intraperitoneal (ip) every 2 days for 32 days. Treatment was discontinued on day 32 and tumor volumes were measured for up to 8 days after treatment withdrawal. Mice were weighed every other day and tumor size was measured every other day as well until the end of the experiment.
Estimation of tumor response to tetrac or tetrac NP
At the conclusion of experiments, all animals were sacrificed in a CO2 chamber and tumor masses were collected and weighed.
Statistical analysis
Statistical analysis was performed by one-way analysis of variance, using Statview software (Adept Scientific, Acton, MA), and the mean ± standard deviation from each experimental group was compared with its respective control group. Statistical significance was defined as p < 0.05.
Results
Cellular distribution of tetrac versus tetrac NP in FTC-236 cells
Tetrac labeled with Cy3 showed cellular uptake and cytoplasmic and nuclear distribution in FTC-236 cells, in contrast to the restricted membrane distribution of tetrac NP (Fig. 1).
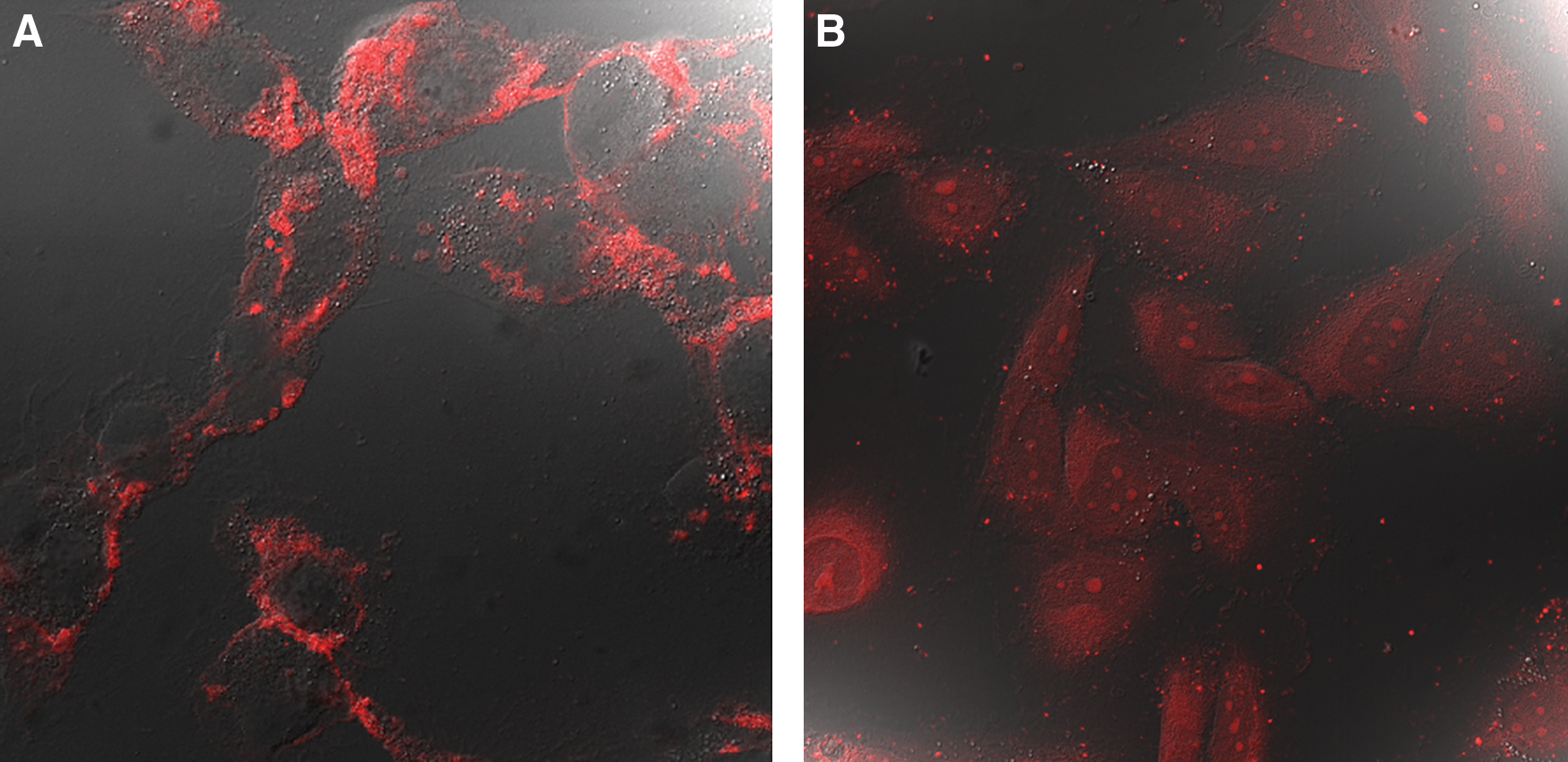
Representative confocal images illustrating the differences between tetrac and tetrac NP distribution in FTC cells. (
Effect of tetrac and tetrac NP on tumor angiogenesis and tumor growth in the CAM model
Cancer cells derived from a human FTC (FTC-236) were implanted at a density of 1 × 106 in the CAM (n = 8 per group) and the effect of tetrac and tetrac NP at 1 μg/CAM on tumor angiogenesis and tumor growth 8 days after implantation in 7-day-old chick eggs was determined. Both tetrac and tetrac NP resulted in significant inhibition (p < 0.001) of tumor-mediated angiogenesis (Fig. 2A) when compared with vehicle-treated controls. Additionally, both tetrac and tetrac NP treatment resulted in effective inhibition (p < 0.01) of tumor growth (Fig. 2B).

Effect of tetrac and tetrac NP on FTC-mediated angiogenesis (
Response of tumor implants to tetrac administration
Daily treatment of xenografted animals with tetrac (1 mg/kg; ip) or tetrac NP (1 mg/kg; ip) resulted in an immediate suppression in tumor volume (Fig. 3). Repeated administration of tetrac (1 mg/kg, ip) and tetrac NP (1 mg/kg, ip) resulted in sustained inhibition of tumor growth up to 30–32 days (Fig. 3) when compared with control (vehicle-treated) animals. In preliminary experiments, there were no differences in tumor growth between animals treated with vehicle alone and void tetrac NP (nanoparticles without tetrac) (data not shown). Tumor regrowth was observed after discontinuation of treatment, but was still below control levels at 14 days (Fig. 4A). At the end of the study, tumor weight was directly measured in the control, tetrac, and tetrac NP groups. Both tetrac (1 mg/kg, ip) and tetrac NP (1 mg/kg, ip) treatment followed by withdrawal for 14 days resulted in 30–40% inhibition of tumor mass (p < 0.01; Fig. 4B).
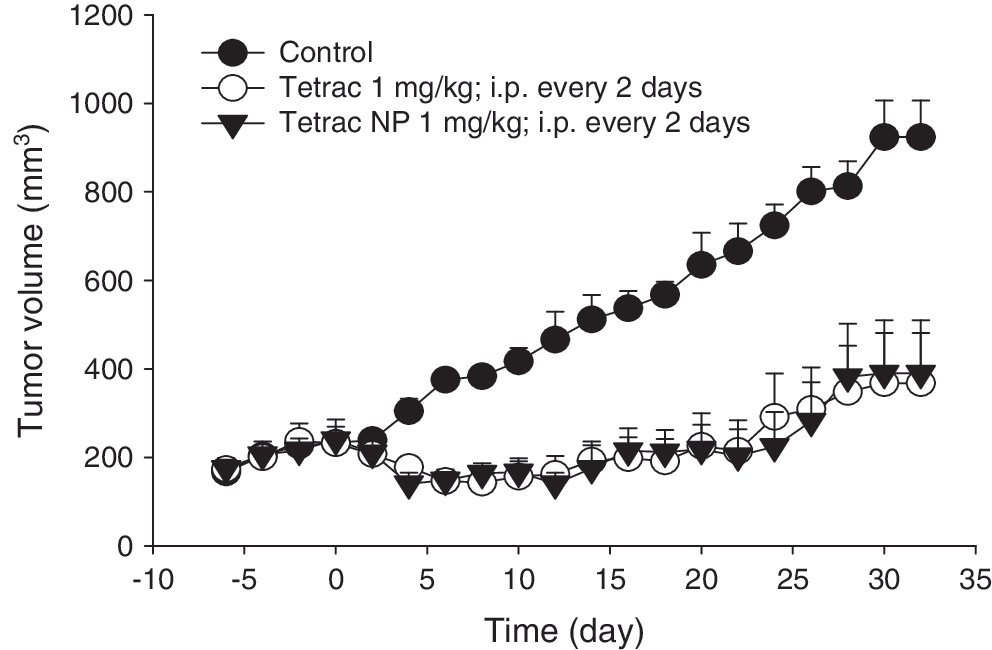
Effect of tetrac and tetrac NP treatment on tumor volume during the treatment period as compared with control. Data represent mean tumor volumes (mm3) ± SEM; n = 8 per group; p < 0.01 for all timepoints from 5–32 days. Treatment period = 32 days administered every other day.
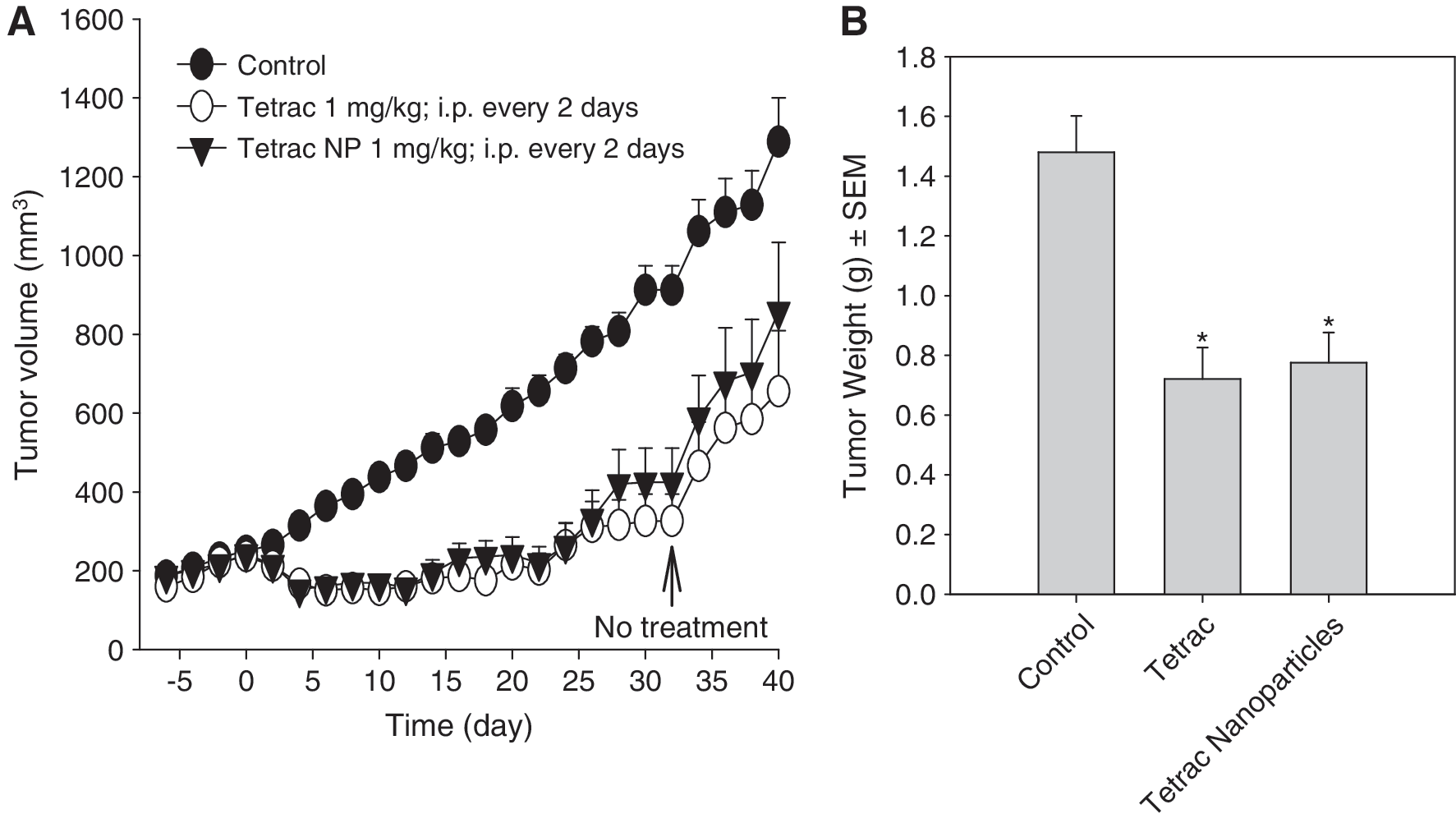
Effect of tetrac and tetrac NP on tumor volume during the treatment period (32 days administered every other day) and following withdrawal of treatment and follow-up for up to day 40 (
Discussion
We showed here that unmodified tetrac and tetrac NP are effective inhibitors of the growth of human FTC xenografts in the nude mouse. Administered parenterally, the agents promptly decreased tumor volume, and over the course of 32 days given every other day, tumor size was significantly reduced. Acting at its integrin receptor, thyroid hormone has been shown to be a proliferative factor for a variety of tumor cells (4,5,19) and to be proangiogenic (1 –3,20,21). Tetrac blocks these effects (3 –5). The pituitary–thyroid axis is intact in the nude mouse (22) and it is assumed that endogenous thyroid hormone is one of the host factors that permits the growth of tumor xenografts, such as those of FTC cells in this study.
Tetrac NP do not gain access to the cell interior and thus tetrac ether bonded to PLGA particles via the outer ring hydroxyl can act only at the integrin receptor—where it is exclusively an antagonist—and not at the nuclear TR. Unmodified tetrac does gain access to the cell interior and can interact with TR, where it is a low-potency agonist (thyromimetic) (7). This study also showed that the antiangiogenic activity of tetrac, which has been shown in the CAM model (4) and in the microvascular endothelial cell-Matrigel assay (20,21), is also apparent in the CAM tumor implant model. Both unmodified tetrac and tetrac NP reduced the vascularity of FTC cell implants in our study. Antiangiogenic agents clinically directed at a specific vascular growth factor have come to be regarded as adjuncts to standard chemotherapy, rather than as primary anticancer modalities (23). We might therefore conclude that the effectiveness of tetrac against FTC secondarily involves antiangiogenesis. On the other hand, tetrac blocks the effects of more than one vascular growth factor, for example, vascular endothelial growth factor and basic fibroblast growth factor, and thus antiangiogenesis mediated by tetrac may be more important than the adjunctive effect obtained clinically with agents that target a single specific vascular growth factor or vascular growth factor receptor.
The action of tetrac on FTC volume was comprised of two effects: a relatively acute effect that was apparent within 3 days of the initiation of tetrac treatment (Figs. 3 and 4A) and a progressive effect. We have ascribed conventional proapoptotic activity to tetrac (13) that would account for the progressive decrease in tumor volume that occurred over 3 weeks of treatment. On the other hand, targeting an integrin with tetrac may induce loss of cell anchorage and a form of apoptosis termed anoikis (24). In this study, the data clearly demonstrate that tetrac and tetrac NP effectively arrest FTC growth in various cancer models and suggest its potential utility in the management of FTC.
Footnotes
Disclosure Statement
The authors declare that they have no competing financial interests in this study.