
Abstract
Background:
Patients with Graves' ophthalmopathy (GO) have circulating autoantibodies directed against the thyrotropin receptor (TSHR) and elevated levels of the proinflammatory cytokine interleukin-6 (IL-6) in both serum and orbital tissues. We hypothesized that these autoantibodies might increase IL-6 expression and secretion in preadipocyte fibroblasts and adipocytes from patients with GO, and thus directly impact the clinical activity of the disease.
Methods:
IL-6 mRNA levels were measured in cultures of GO orbital preadipocytes (n = 3) treated during adipocyte differentiation with a monoclonal stimulatory TSHR antibody (M22; 10 ng/mL), IL-6 (1 ng/mL), or TSH (10 U/L). Additionally, levels of IL-6 protein secretion were assessed after adipocyte differentiation in orbital cultures exposed to TSH or M22 for 24 or 48 hours (n = 8). IL-6 mRNA levels were also measured in orbital adipose tissue specimens from well-characterized GO patients (n = 9) and normal individuals (n = 9).
Results:
Treatment of GO orbital preadipocyte cultures with IL-6, TSH, or M22 during adipocyte differentiation resulted in increased IL-6 mRNA levels (3.1-fold, 2.9-fold, and 2.7-fold, respectively; p < 0.05). Treatment of orbital cultures with M22 or TSH after adipocyte differentiation enhanced the release of IL-6 protein into the medium at both 24 and 48 hours for TSH (mean 1.9- and 2.3-fold; p = 0.002 and 0.015, respectively) and at 48 hours for M22 (mean 2.0-fold; p = 0.005). In addition, we found mean IL-6 mRNA levels to be significantly increased in GO orbital adipose tissue specimens (10-fold; p < 0.01), primarily attributable to high levels in three of the four patients with clinical activity scores ≥5.
Conclusions:
Both TSH and M22 increase IL-6 expression in orbital preadipocyte fibroblasts and IL-6 secretion by mature adipocytes. These results suggest that circulating TSHR autoantibodies in GO might play a direct role in the clinical activity of the disease.
Introduction
Levels of the proinflammatory cytokine interleukin-6 (IL-6) are elevated in the sera of patients with Graves' hyperthyroidism, particularly in the subgroup having clinically significant GO (22 –24). In addition, studies have shown that IL-6 is expressed within the GO orbit (25 –28) with the highest levels found in tissues from patients with active GO (20). We have shown that cultured GO orbital fibroblasts increase TSHR expression after IL-6 treatment (29), suggesting that this cytokine within the GO orbit may act to enhance local levels of the autoantigen and thus augment the autoimmune response.
We undertook the current study to better understand mechanisms responsible for the elevated levels of IL-6 in GO orbital tissues. To this end, we assessed the impact of a human monoclonal stimulatory antibody directed against the TSHR (M22), TSH, and IL-6 itself on IL-6 expression in orbital preadipocyte cultures undergoing adipocyte differentiation. In other studies, because mature adipocytes are known to secrete IL-6, we measured the effect of M22 or TSH exposure on the secretion of IL-6 protein in orbital cultures after adipocyte differentiation. In addition, we measured IL-6 mRNA levels in orbital tissue specimens obtained from well-characterized patients with GO.
Materials and Methods
Cell cultures
Orbital tissue samples were obtained during the course of orbital decompression surgery for severe GO. The tissues were transported to the laboratory, minced, and placed directly in plastic culture dishes, allowing preadipocyte fibroblasts to proliferate as described previously (30). Cells were initially propagated in medium 199 containing 20% fetal bovine serum (FBS; HyClone Laboratories, Logan, UT), penicillin (100 U/mL), and gentamicin (20 μg/mL) in a humidified 5% CO2 incubator at 37°C and maintained in 75-mm2 flasks with medium 199 containing 10% FBS and antibiotics. The study was approved by the Mayo Clinic Institutional Review Board.
To initiate adipocyte differentiation, orbital cells were grown to confluence in six-well plates in medium 199 with 10% FBS. Differentiation was carried out over 10 days as reported previously (31). Briefly, confluent cultures were fed serum-free Dulbecco's Modified Eagle's Medium (DMEM)/Ham's F-12 (1:1; Sigma, St. Louis, MO) supplemented with biotin (33 μM), pantothenic acid (17 μM), transferrin (10 μg/mL), T3 (0.2 nM), insulin (1 μM), carbaprostacyclin (cPGI2; 0.2 μM; Calbiochem, La Jolla, CA), and, for the first 4 days only, dexamethasone (1 μM) and isobutylmethylxanthine (0.1 mM). The medium was replaced every 3–4 days during the 10-day differentiation period.
In studies aimed at determining the impact of a human stimulatory monoclonal antibody directed against TSHR [M22; Kronus, Boise, ID; # M22-1b; (32)], IL-6 (R&D Systems, Minneapolis, MN; # 206-IL), or bovine TSH (Sigma Aldrich, St. Louis, MO; # T-8931) on IL-6 mRNA levels during adipocyte differentiation, GO orbital preadipocytes (n = 3) were cultured in differentiation media for 10 days in the presence or absence of M22 (10 ng/mL), IL-6 (1 ng/mL), or bovine TSH (10 U/L). Results were expressed as mean fold elevation of IL-6 mRNA (normalized to 18S rRNA) relative to parallel untreated (control) GO cultures.
In other studies aimed at measuring the effect of M22 or TSH on the secretion of IL-6 protein from mature adipocytes, GO orbital preadipocytes (n = 8) were cultured in differentiation media containing the peroxisome proliferator γ (PPARγ) agonist rosiglitazone (10 μM) to enhance adipogenesis and TSHR expression (33). After 10 days in culture, cells were treated for 24 or 48 hours with M22 (10 ng/mL) or bovine TSH (10 U/L). Supernatants were then aspirated for assessment of IL-6 protein levels as described below. Results were expressed as mean fold elevation of IL-6 protein relative to parallel untreated (control) cultures.
Whole orbital tissue specimens
Orbital adipose/connective tissue specimens were obtained from patients with GO who underwent orbital decompression surgery for severe disease (n = 9; Table 1) and from individuals with no history of Graves' disease (n = 9). All GO patients were euthyroid on thyroid hormone replacement therapy at the time of orbital surgery. The clinical activity score (CAS), a measure of inflammatory disease activity (34), was determined by an ophthalmologist on physical examination before the orbital decompression surgery. We used orbital tissues retrieved at very early autopsy from patients (without GO) whose corneas had been harvested for transplantation as normal control tissue. The early autopsy tissues were retrieved between 6 and 20 hours after death with the average being 12 hours. Each orbital adipose/connective tissue specimen was placed in a sterile container on saline-soaked gauze, and transported at room temperature to the laboratory, where it was either frozen immediately at −70°C or placed in culture. Because tissue samples obtained during orbital decompression surgery are quite small (e.g., approximately 0.3 g), patients whose whole orbital tissues were studied (n = 9) were not the same as those whose orbital preadipocyte fibroblasts were propagated for use in the tissue culture-based studies.
TSI, thyroid-stimulating immunoglobulin index (normal < 1.3); ND, not done; CAS, clinical activity score; GO, Graves' ophthalmopathy.
Real-time reverse transcription-polymerase chain reaction analyses
Total RNA was isolated from whole orbital tissue specimens or orbital cultures using the RNeasy kit (Qiagen, Valencia, CA) according to the manufacturer's protocol. cDNA was synthesized using total RNA (200 ng for orbital tissues and 500 ng for orbital cultures) incubated with random hexamers, followed by a reverse transcription reaction (100 μL) with Multiscribe Reverse Transcriptase (6.25 units; Applied Biosystems, Foster City, CA). Conditions used were 25°C for 10 minutes, 37°C for 60 minutes, and 95°C for 5 minutes. Oligonucleotide primers and TaqMan probes for IL-6, 18S rRNA, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were purchased from Applied Biosystems. Quantitative polymerase chain reactions were done in a 96-well optical reaction plate. Amplification reactions contained cDNA equivalent of 2 ng total RNA for orbital tissues and 35 ng total RNA for orbital cultures, 900 nM of the forward and reverse primers, and 250 nM of the probes in a volume of 25 μL using the Universal TaqMan 2 × polymerase chain reaction mastermix (Applied Biosystems). The thermal cycling conditions used were 2 minutes at 50°C for optimal AmpErase uracil N-glycosylase (UNG) activity and 10 minutes at 95°C to activate Amplitaq Gold DNA Polymerase, followed by 40 cycles at 95°C for 15 seconds and 60°C for 1 minutes. IL-6 rRNA and 18S rRNA were amplified in separate wells. All reactions were performed in duplicate in the ABI Prism® 7700 Sequence Detector (Applied Biosystems), and the data pooled. Normal human adipose tissue was used as the IL-6-positive control. Reaction mixture, without the cDNA, was used as the negative control in each run.
The standard curve method was used to quantify the expression of IL-6 mRNA in each sample, and normalized results were expressed as the ratio of IL-6 mRNA (pg) to 18S or GAPDH mRNA (pg). The relative expression level of IL-6 in GO orbital adipose tissue specimens or cultures was compared with that of tissue specimens from normal individuals or untreated parallel cultures, respectively. The Mann–Whitney Rank Sum test was used to assess statistically significant differences between the groups.
ELISA for IL-6 protein secretion
After adipocyte differentiation in the presence of rosiglitazone, GO orbital cultures were refed differentiation media containing M22 (10 ng/mL) or bovine TSH (10 U/L), and supernatants were aspirated 24 or 48 hours later for determination of IL-6 protein levels using an enzyme immunometric assay (Assay Designs, Ann Harbor, MI; # 900–033). Results were normalized to total cellular protein (as measured by Bradford protein assay) and analyzed by unpaired t-test using Sigma Stat statistical software version 3.5 (Systat Software, San Jose, CA).
Results
Orbital cultures
The first set of studies was performed to assess the effect of M22, IL-6, or TSH treatment on IL-6 gene expression during adipocyte differentiation. For these studies, we used our cell culture model of early GO in which orbital fibroblasts undergo differentiation into mature fat cells (31). Treatment of GO orbital preadipocyte cultures (n = 3) with M22 (10 ng/mL) during differentiation enhanced IL-6 mRNA levels (mean 2.7-fold, range 1.8–3.6, p = 0.03; Fig. 1). Similarly, IL-6 (1 ng/mL) and TSH treatment increased IL-6 mRNA (mean 3.1-fold, range 2.3–3.9, p < 0.05 and mean 2.9-fold, range 2.4–3.6, p = 0.005, respectively; Fig. 1).

Relative level of IL-6 mRNA (mean ± standard error of the mean) in cultures of GO orbital preadipocytes (n = 3) treated with IL-6 (1 ng/mL), bovine thyrotropin (10 U/L), or M22 (10 ng/mL) during differentiation compared with parallel untreated cultures (3.1, 2.9, 2.7-fold; *p < 0.05). Data represent mean fold elevation relative to parallel untreated (control) GO cultures. GO, Graves' ophthalmopathy; IL-6, interleukin-6.
Because mature adipocytes are known to secrete IL-6 protein, other studies were performed to assess the impact of M22 or TSH on IL-6 protein secretion by mature fat cells. Therefore, we treated GO orbital cultures (n = 8) with the PPARγ agonist rosiglitazone (10 μM) for 10 days to enhance adipocyte differentiation. After that, we exposed the cultures to either bovine TSH (10 U/L) or M22 (10 ng/mL) for 24 or 48 hours. We found that TSH increased secretion of IL-6 protein at both 24 and 48 hours (mean 1.9 and 2.3-fold; p = 0.002 and 0.015, respectively) and that M22 significantly increased secretion of IL-6 protein at 48 hours (mean 2.0-fold; p = 0.005; Fig. 2).
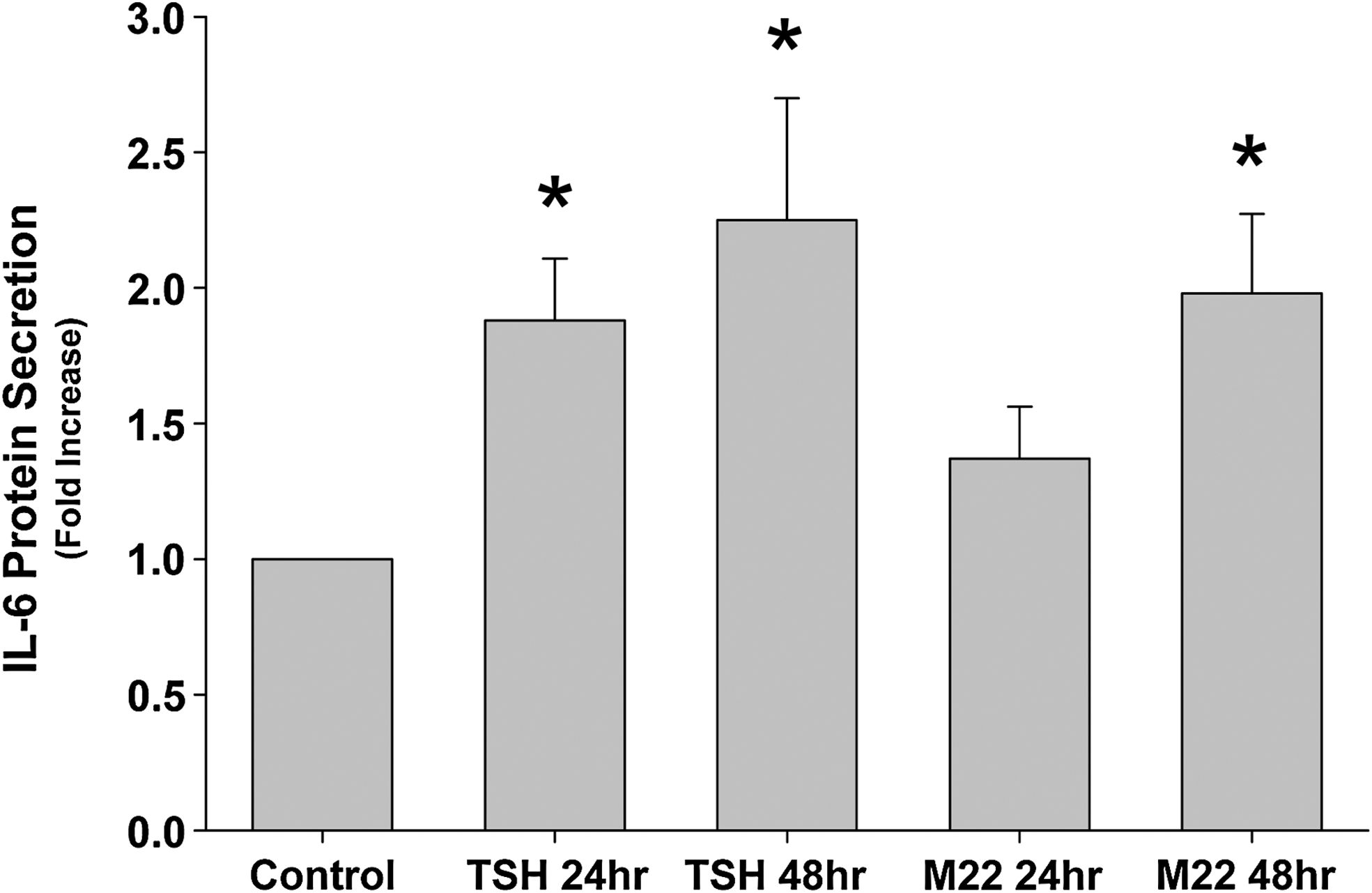
IL-6 protein secretion in cultures (n = 8) treated for 24 or 48 hours with bovine thyrotropin (10 U/L; 1.8-fold; p = 0.002 and 2.3-fold; p = 0.015, respectively) or M22 (10 ng/mL; 1.4-fold; p = 0.076 and 2.0-fold; *p = 0.005, respectively) after a 10-day period of adipocyte differentiation in the presence of rosiglitazone. Data represent mean fold elevation relative to parallel untreated (control) cultures.
Whole tissue specimens
IL-6 mRNA was measurable in each of the GO (n = 9) and normal (n = 9) orbital adipose tissue specimens studied with the mean expression levels in the GO tissues being 10-fold higher than in the normal tissues (mean = 12.05 and 1.19 IL-6/18S, respectively; p < 0.01; Fig. 3). This increase was attributable to high levels in 3/9 GO patients (9.8, 27, and 56.9 IL-6/18 S) above those measured in any normal tissue (range 0.18–3.2 IL-6/18 S). To confirm that the normal (i.e., cadaveric) tissues had not undergone significant RNA degradation, we compared the absolute values of the GAPDH expression between the GO and cadaveric specimens. We found that the mean Ct of the GAPDH in the cadaveric tissues was 18.35 cycles (standard deviation 0.933), whereas the mean Ct of the GAPDH in the GO specimens was 18.13 cycles (standard deviation 1.14), a result suggesting that the cadaveric tissue RNA was indeed intact.

Relative expression of IL-6 mRNA (normalized to 18S rRNA) in orbital adipose tissue specimens from patients with GO (n = 9) and normal individuals (n = 9). The mean IL-6 level in the GO specimens was 10-fold above the mean of the normal group (*p < 0.01) and was attributable to high levels in 3/9 GO patients.
The patient with the highest expression of IL-6 (Table 1; patient #1) was very severely affected, having the longest duration of GO, a CAS of 6, proptosis measurements of 34/35 mm, and bilateral optic neuropathy. The other two patients with elevated IL-6 mRNA levels (Table 1; patients #2 and #3) also had active and severe disease with CAS of 5 and optic neuropathy. However, another patient with very active and severe disease (Table 1; patient #8) had one of the lowest orbital tissue IL-6 mRNA levels. We did not find a significant correlation between IL-6 expression and CAS. Although we similarly found no correlation between IL-6 expression levels and thyroid-stimulating immunoglobulin levels, the validity of this result is questionable as thyroid-stimulating immunoglobulin was measured at the time of decompression surgery in only 4/9 GO patients.
Discussion
IL-6 is a proinflammatory cytokine synthesized by many different cell types, including thyrocytes, monocytes, fibroblasts, macrophages, preadipocytes, and mature adipocytes (35 –38). This cytokine plays important roles in autoimmune and chronic inflammatory disorders and also functions as a stimulator of B cell differentiation (36,37). We have shown in gene array studies that the expression of various genes encoding adipocyte-related proteins, including IL-6, adiponectin, and PPARγ, is significantly elevated within the GO orbit (6,39). These findings suggest that de novo adipogenesis is active within these tissues and that the adipocytes themselves may be a primary source of orbital IL-6 in GO. Additionally, this cytokine may be macrophage derived, as a host of cytokines produced by macrophages have been shown to be elevated in GO orbital tissues (10,12,25,26,40).
Orbital adipocytes have been shown to express TSHR (1 –5,31), and levels of circulating autoantibodies targeting the TSHR have been shown to correlate with the CAS in patients with GO (41). To explain this association, we hypothesized that TSHR autoantibodies in GO might act to stimulate IL-6 production and secretion by mature adipocytes within the orbit. Our finding that treatment with the TSHR antibody M22 enhanced both IL-6 mRNA levels during adipocyte differentiation (Fig. 1) and IL-6 protein secretion by mature adipocytes (i.e., after the 10-day differentiation period; Fig. 2) is consistent with this hypothesis. It may be that the close temporal association between Graves' hyperthyroidism and the onset of the signs and symptoms of GO (42) may be in part related to TSHR-autoantibody-mediated stimulation of IL-6 secretion within the orbit. Because levels of IL-6 produced by mature adipocytes correlate with the size of the fat cells (43), it is possible that the high IL-6 levels found in active orbital tissues reflect the de novo adipogenesis considered to be an important component of the disease process (6,18,19). Whether TSHR autoantibodies in the sera of patients with active GO are directly responsible for the stimulation of new fat cell development within the orbit is an area of active investigation. Our results are in some respects similar to those of Janson and colleagues, who demonstrated that the lipolytic effect of TSH on lipolysis in infant adipocytes can be mimicked by stimulatory TSHR autoantibodies and blocked by inhibitory autoantibodies in IgG from Graves' patients (44). These authors suggested that the action of TSHR autoantibodies in fat cells may be coupled to the adenylate cyclase system. It is possible that the stimulation of IL-6 secretion in our cultures (and, potentially, the stimulation of adipogenesis) by TSHR stimulatory antibodies is also mediated through this signaling system, as agents that increase intracellular cAMP are known to induce adipocyte differentiation (45).
We studied the impact of TSH treatment on orbital cultures and found that this hormone (also targeting TSHR) acts similarly to the TSHR antibody (M22) in that it increases IL-6 expression when cultures are treated during differentiation (Fig. 1) and also significantly enhances the secretion of IL-6 protein from mature adipocytes (Fig. 2). This finding is consistent with a previous report showing that TSH stimulates IL-6 in cultured 3T3-L1 and human abdominal adipose cells (38,46). It is possible that the increase in CAS observed in some GO patients who experience hypothyroidism with elevated TSH levels after radioiodine ablation for hyperthyroidism (47) may be in part owing to this direct effect of TSH on orbital adipocytes. Additionally, orbital IL-6 levels in this setting may be further elevated by the sustained increase in TSHR autoantibody levels that has been documented after radioiodine therapy (48).
We found particularly elevated levels of IL-6 in 3/9 GO orbital tissue specimens that were higher than any found in normal specimens (Fig. 3). Each of these three patients had active disease with CAS of 5–6. However, a single patient with a comparable CAS had an IL-6 level that was only minimally above the mean of the normal tissues (Table 1; patient #8). A study by Wakelkamp and colleagues found IL-6 mRNA in orbital tissue from only 1 of 6 patients with active GO to be higher than levels found in 11 patients with inactive disease (CAS < 3) (20). These findings differ from ours, as we were able to detect IL-6 mRNA in all normal and GO tissues. We did however find a trend toward higher levels in tissues from patients having the more active disease (data not shown). IL-6 is a regulator of inflammation and of the acute-phase response (36). No patient in our study had early GO at the time of orbital decompression surgery. In fact, the patients with the shortest time since GO diagnosis (6 months) had one of the lowest IL-6 tissue levels. Therefore, our study suggests that this cytokine may not function primarily as an acute phase reactant in GO, but may serve as a mediator of inflammation in later disease. In fact, chronic inflammation itself is known to induce IL-6 production, which is related to polyclonal B cell activation and antibody production (36). It is possible, that the extremely high IL-6 levels found in these three patients are a result of the chronic inflammation present in longstanding GO. This cytokine may also play a role in B cell differentiation and local TSHR antibody production.
A potential limitation to this portion of our study is that the normal control orbital tissue was obtained at very early autopsy from patients (without GO) whose corneas had been harvested for transplantation. We choose not to use tissue from orbital enucleation surgeries as our control because we obtain the majority of those from patients undergoing surgery for orbital malignancies. Since these conditions are associated with elevated cytokine levels, we felt that these specimens would not serve as an appropriate negative control. To demonstrate that it is unlikely that the differences in IL-6 mRNA levels between the GO and cadaveric tissue specimens are due to RNA degradation in the latter, we demonstrated that the mean Ct values for the housekeeping gene 18S in the GO and normal samples were essentially the same (see Results), supporting that the cadaveric RNA was intact.
In summary, we demonstrated that IL-6, TSH, and a stimulatory TSHR antibody (M22) increase IL-6 expression in cultured orbital preadipocytes and that both TSH and M22 stimulate IL-6 protein secretion by mature orbital adipocytes. These findings suggest that circulating autoantibodies directed against the TSHR in Graves' disease may act to increase IL-6 levels and secretion within the orbit and that IL-6 in turn may act locally to stimulate its own production. In addition, the B cell differentiating effects of IL-6 may lead to further production of TSHR autoantibodies within the orbit in GO. We found particularly high levels of IL-6 mRNA within the orbits of some patients with active disease of medium to long standing, suggesting that this cytokine may play a role in GO disease activity and propagation. These results suggest that agents inhibiting IL-6 action, such as the novel humanized monoclonal antibody tocilizumab, may prove useful in some GO patients with especially active disease (49).
Footnotes
Acknowledgment
Supported in part by Grant DK77814 from the National Institute of Diabetes, Digestive, and Kidney Diseases.
Disclosure Statement
None of the authors has any commercial associations that might create a conflict of interest in connection with this article.