
Abstract
Background:
Orbital fibroblasts are involved in the pathogenesis of Graves' ophthalmopathy (GO) by producing hyaluronan (HA), synthesized by three types of hyaluronan synthases (HAS1, HAS2, and HAS3). Thyrotropin receptors (TSHR) expressed in orbital fibroblasts activate the cyclic adenosine monophosphate (cAMP) pathway. Only sparse data are available at present supporting a role for TSHR activation in the regulation of HA in GO orbital fibroblasts. We hypothesize that TSHR activation, via cAMP signaling, results in induction of HAS1–3 mRNA expression and HA production by nondifferentiated GO orbital fibroblasts.
Methods:
Cultured nondifferentiated orbital fibroblasts obtained during orbital decompression surgery from 15 GO patients were stimulated with recombinant human TSH (rhTSH), TSHR-stimulating Graves' disease immunoglobulin G (GD-IgG) or forskolin (FSK), or interleukin-1β (IL-1β).
Results:
FSK significantly stimulated cAMP production, HAS1 and HAS3 mRNA expression, and HA secretion in orbital fibroblasts. IL-1β slightly induced cAMP production, but induced HAS mRNA expression of all three isoforms and HA secretion. In contrast, the effects of rhTSH and GD-IgG on cAMP were modest and absent, respectively, and on HAS mRNA and HA synthesis were completely absent.
Conclusions:
The strong increase in cAMP synthesis by FSK in nondifferentiated GO orbital fibroblasts results in increased HA synthesis, but TSHR activation by rhTSH or GD-IgG does not result in altered HA synthesis. Our results do not support a predominant role for GD-IgGs in the accumulation of orbital glycosaminoglycans; cytokines like IL-1β seem largely responsible for excessive glycosaminoglycan production by nondifferentiated orbital fibroblasts in early immunopathogenesis of GO.
Introduction
There is increasing evidence that orbital fibroblasts are major participants in the immunopathogenesis of GO (1). They exhibit heterogeneous phenotypes (2); a subpopulation, termed preadipocytes, can differentiate into mature adipocytes under strict culture conditions in vitro and may contribute to the increase in adipose tissue (3 –5).
Orbital fibroblasts in vitro respond to various mediators of inflammation, such as interleukin-1β (IL-1β), by producing excessive amounts of the markedly hydrophilic GAG hyaluronan (HA) (2,6,7), which is the major GAG in orbital tissues of GO patients (8,9). HA can be synthesized at the plasma membrane by three mammalian hyaluronan synthases (HAS); HAS1, HAS2, and HAS3 are differentially inducible in various cell types by several cytokines and growth factors (10,11).
Orbital fibroblasts express a functional thyrotropin receptor (TSHR) (12 –15), the antigen that binds Graves' disease immunoglobulin G (GD-IgG) in thyroid follicular epithelial cells, leading to excessive production of thyroid hormone (16,17). TSHR gene expression is increased in orbital fatty connective tissue of patients with GO in the active stage of the disease compared to patients with inactive GO (18). In vitro functional TSHR expression in orbital fibroblasts is increased upon differentiation into adipocytes (5). Moreover, a correlation between the level of TSHR autoantibodies and the clinical activity of GO has been reported (19 –21). Taken together, these findings suggest that the TSHR is a major autoantigen in the immunopathogenesis of GO.
Activation of the TSHR by TSH or GD-IgG, through G-protein-coupled pathways, results in activation of the adenylyl cyclase/cyclic adenosine monophosphate (cAMP) pathway (22,23). cAMP was described to induce HA synthesis in several cell types, among which orbital fibroblasts (24 –27). Only sparse data have been reported to date on the effects of GD-IgG on HA synthesis in nondifferentiated orbital fibroblasts. Smith and Hao reported GD-IgG to induce HA synthesis, but suggested the insulin-like growth factor-1 receptor (IGF1R) to be involved (28). Zhang et al. (29) did not find altered HA production in GO orbital fibroblasts after incubation with a monoclonal thyroid-stimulating antibody.
There is accumulating evidence that cell-mediated immunity prevails in early disease pathogenesis of GO, as suggested by a predominance of Th1-type T-cells and Th1-type-cell-derived cytokines like IL-1β in early disease (1,30,31). In this study, we hypothesized that nondifferentiated orbital fibroblasts best approach the population present in the orbit during the early stage of GO when adipocytic differentiation with increased TSHR expression has not yet occurred. Therefore, we systematically evaluated TSHR activation by recombinant human TSH (rhTSH) and GD-IgGs and its effects on cAMP signaling and HA synthesis in cultured, nondifferentiated orbital fibroblasts obtained from surgical specimens of several patients with GO.
Materials and Methods
Graves' disease IgG and control IgG isolation
We isolated IgG from sera of two GO patients (GD-IgG) and two healthy control subjects (c-IgG) using protein G Sepharose 4 Fast Flow (ProtG; Amersham Pharmacia Biotech Benelux, Roosendaal, The Netherlands), after obtained informed consent. TSH-binding inhibitory immunoglobulin (TBII) serum levels were determined by a second-generation luminescence receptor assay (DYNOtest TRAK assay; B.R.A.H.M.S Diagnostica, Berlin, Germany) calibrated to the World Health Organization (WHO) standard 90/672. The patients with Graves' disease had been found to have mild (CAS score 3/7) and moderately severe (CAS score 4/7) GO, respectively (32), and had not been treated with steroids. At the time of the blood sampling, the GO was inactive in both patients.
Orbital fibroblast cell culture
Orbital fatty tissue explants were obtained from 15 patients with severe GO undergoing orbital decompression surgery. Three patients received 1 g of corticosteroids on the day or the day previous to surgery. Twelve patients underwent decompression surgery in the inactive stage of the disease and had not received corticosteroids in the previous 6 months. Collection of these tissues was approved by our Institutional Review Board.
Orbital fibroblast cell lines were initiated as described previously (6,33). We cultured fibroblast cell lines in six-well plates with medium 199 (BioWhittaker, Lonza, Verviers, Belgium) supplemented with 10 U/mL penicillin, streptomycin, Fungizone, and 10% fetal calf serum (FCS; all from Cambrex, East Rutherford, NJ), and cultures were used between the third and eight passage from initiation.
For each experiment, 5 × 104 orbital fibroblasts/well were grown to confluence in six-well plates containing the culture medium in approximately 48 hours. Cells were incubated with supraphysiological 10 mU/mL rhTSH (Thyrogen®; Genzyme Therapeutics, Cambridge, MA), 1 mg/mL GD-IgG (batch I or II), 1 mg/mL c-IgG (batch I or II), or 50 μM forskolin (FSK; Sigma-Aldrich, Zwijndrecht, The Netherlands), a direct activator of the adenylyl cyclase/cAMP pathway. We also employed incubation with IL-1β (10 ng/mL; Sigma-Aldrich) as a positive control because it has been reported previously that IL-1β induces expression of the three HAS mRNA isoforms and HA production in orbital fibroblasts (6). We chose to perform the experiments with rhTSH, as we preferred to study a homologous system. To evaluate the effect of GD-IgG on orbital fibroblast cell cultures, a dose–response experiment was performed in orbital fibroblasts of two patients with inactive GO. c-IgG and GD-IgG were added in concentrations of 100 ng/mL, 1 μg/mL, 10 μg/mL, 100 μg/mL, and 1 mg/mL; cAMP production, HAS mRNA expression, and HA content in the supernatant were measured, as described below.
cAMP-responsive luciferase assay
Chinese hamster ovary cell line (CHO-K1; ATTC No. CCL-61; LGC Standards, Middlesex, United Kingdom) stably transfected with the human TSHR and a cAMP-responsive (CRE) luciferase gene (CHO.hTSHR.LUC) was generated at Schering-Plough Research Institute and cultured in Dulbecco's Modified Eagle Medium (DMEM)/F12 modified medium (Gibco, Breda, The Netherlands) containing 5% FCS, 100 U/mL penicillin/0.1 mg/mL streptomycin, 0.4 mg/mL hygromycin B (Invitrogen, The Netherlands; Greiner, Alphen a/d Rijn, The Netherlands). The human TSHR complimentary DNA was kindly provided by Prof. E. Milgrom, INSERM (34). Cells were incubated overnight in a humidified atmosphere with 5% CO2 at 37°C. One day before the assay, the culture medium was replaced with serum-free DMEM/F12 supplemented with 1 μg/mL bovine insulin (Sigma-Aldrich), 5 μg/mL human apo-transferrin (Sigma), and 0.8% penicillin/streptomycin. The next day, cells were resuspended in the assay medium (DMEM/F12 modified medium [Gibco], 1 μg/mL bovine insulin [Sigma-Aldrich], 5 μg/mL human apo-transferrin [Sigma-Aldrich], 100 U/mL penicillin, 0.1 mg/mL streptomycin, and 20 μM rolipram [Sigma-Aldrich]). Cells were seeded in a 384-well assay plate (1.10E-6 cells/mL, 10 μL/well).
For the dose–response curves of rhTSH, 5 μL rhTSH in concentrations of 1.00E-10, 3.16E-9, 1.00E-8, 3.16E-8, 1.00E-7, 3.16E-7, 1.00E-6, 1.00E-6, 1.00E-5, and 3.16E-5 g/mL was pipetted in quadruplicate into the 384-well assay plate. For the dose–response curves of c-IgG and GD-IgG, 5 μL IgG in concentrations of 1.00E-2, 3.16E-3, 1.00E-3, 3.16E-4, 1.00E-4, 3.16E-5, 1.00E-5, and 1.00E-6 g/mL was pipetted in quadruplicate into the 384-well assay plate. After adding 5 μL of 4% dimethyl sulfoxide/assay medium, cells were incubated for 4 hours in a humidified atmosphere with 5% CO2 at 37°C. Subsequently, Steadyliteplus solution (PerkinElmer, Groningen, The Netherlands) dissolved in Reconstitution buffer according to the manufacturer's protocol was added, and the cells were lysed for 45–60 minutes in the dark. The CRE-luciferase activity was measured using a Packard Topcount luminescence counter. We tested the IgG batches in two separate runs. In both runs stimulation with TSH resulted in a similar induction in luciferase activity (approximately fivefold stimulation over basal values). The luciferase signal is expressed as percentage of the maximum luciferase activity elicited by rhTSH.
cAMP assay
cAMP measurements were performed in the supernatant of three GO orbital fibroblast cultures. Plates were preincubated in 750 μL of the culture medium containing 0.5 mM IBMX, for 30 minutes at 37°C, 5% CO2. Then, 10 mU/mL rhTSH, 1 mg/mL GD-IgG (batch I or II), 1 mg/mL c-IgG (batch I or II), or 50 μM FSK was added, and cells were incubated for another 6 hours at 37°C, 5% CO2. We used orbital fibroblasts of three GO patients; each incubation was performed in quadruplicate. Culture supernatants were collected and stored in −80°C until use. Cell layers were washed with saline and lysed in distilled water, and the total amount of protein per well was measured using the Bio-Rad protein assay (Bio-Rad Laboratories, Richmond, CA). cAMP concentrations were corrected for total amount of protein. The cAMP concentrations in the supernatants were measured in triplicate, using the Alphascreen cAMP Assay (PerkinElmer) as described previously (35).
RNA isolation and quantitative real-time polymerase chain reaction
Experiments for mRNA expression were performed in nondifferentiated orbital fibroblasts of five GO patients (six wells/group), except for the experiments with IL-1β, which we conducted in three GO orbital fibroblast cultures. Cells were lysed in 250 μL lysis buffer supplied with the Magna Pure LC RNA Isolation kit–High Performance (Roche Molecular Biochemicals, Mannheim, Germany). Orbital fibroblast RNA was isolated with the relevant Magna Pure kit using the protocol and buffers supplied. We measured RNA amounts using the Nanodrop (Wilmington, DE) to be able to perform cDNA synthesis with equal RNA input for all experiments. cDNA synthesis was performed using the First-Strand cDNA Synthesis kit for real-time (RT)-polymerase chain reaction (PCR) with oligo d(T) primers (Roche Molecular Biochemicals). Real-time PCR was performed using the Lightcycler 480 (Roche Molecular Biochemicals). For all PCRs, the LightCycler 480 SYBR Green I Master kit (Roche Molecular Biochemicals) was used, adding 50 ng (5 μM for HAS1, HAS2, and HAS3) primers (Biolegio, Nijmegen, The Netherlands) each. Primer pairs for HAS1, HAS2, HAS3, TSHR, and IGF1R have been reported previously (18,36,37). Glyceraldehyde 3 phosphate dehydrogenase (GAPDH) was used as housekeeping gene, as described before (38). Fluorescence values of the samples were baseline corrected and individually checked for their PCR efficiency using LC480 Conversion and LinRegPCR software (version 11.x), kindly provided by Dr. J.M. Ruijter (Amsterdam, The Netherlands). The median of the efficiency was calculated for each assay and samples that had a greater difference than 0.05 of the efficiency median value were not taken into account (0–5%). The original amounts of target cDNA were calculated by LinRegPCR software; calculation was based on the mean PCR efficiency of the amplicon (39).
HA assay
HA measurements were performed in culture supernatants of nondifferentiated orbital fibroblasts of three GO patients (four wells/group), except for the experiments with FSK, which we conducted in four GO orbital fibroblast cultures. We quantified the amount of HA with a specific ELISA-kit (Echelon Biosciences, Salt Lake City, UT) according to manufacturer's instructions. Orbital fibroblast cultures were incubated for 48 hours in 750 μL of the culture medium containing 10 mU/mL rhTSH, 1 mg/mL GD-IgG (batch I or II), 1 mg/mL c-IgG (batch I or II), or 50 μM FSK. Supernatants were collected and stored at −20°C until use. Cell layers were rinsed carefully with saline and lysed in distilled water for protein determination using the Bio-Rad protein assay (Bio-Rad Laboratories). We measured HA concentrations in duplicate and concentrations were corrected for total amount of protein per well.
Statistical analysis
Normal distribution of the data was tested using the Kolmogorov–Smirnov test. Student's t-test was used to test statistical significance between groups if data were normally distributed; Mann–Whitney U-test was used when data were not normally distributed. All tests were performed using SPSS (SPSS, Chicago, IL).
Results
c-IgG and GD-IgG
GD-IgG were purified from sera of two patients with inactive GO and from plasma of two healthy control subjects. Both Graves' disease patients had very high levels of TBII (88 and 256 U/L), whereas the healthy control subjects were TBII-seronegative (<1.0 U/L). Purified c-IgG and GD-IgG were tested for CRE-luciferase activity in the CHO.hTSHR cell line to determine the ability of TSHR activation. Figure 1 shows the dose–response curves with increasing concentrations for rhTSH, both c-IgGs and both GD-IgGs. The two GD-IgGs (1 mg/mL) showed activities comparable to the activity induced by, respectively, 10−7 and 10−6 g/mL rhTSH (≈ 0.5–5 mU/mL). The c-IgG batches showed little CRE-luciferase activity. This residual activity is not related to activation of the TSHR because it was also seen, with a similar magnitude, in CHO cells expressing only CRE luciferase (data not shown). We therefore chose to use the dose of 1 mg/mL IgG in subsequent experiments. The dose of 10 mU/mL rhTSH was chosen because this concentration of rhTSH elicits a marked effect on the CRE luciferase activity.
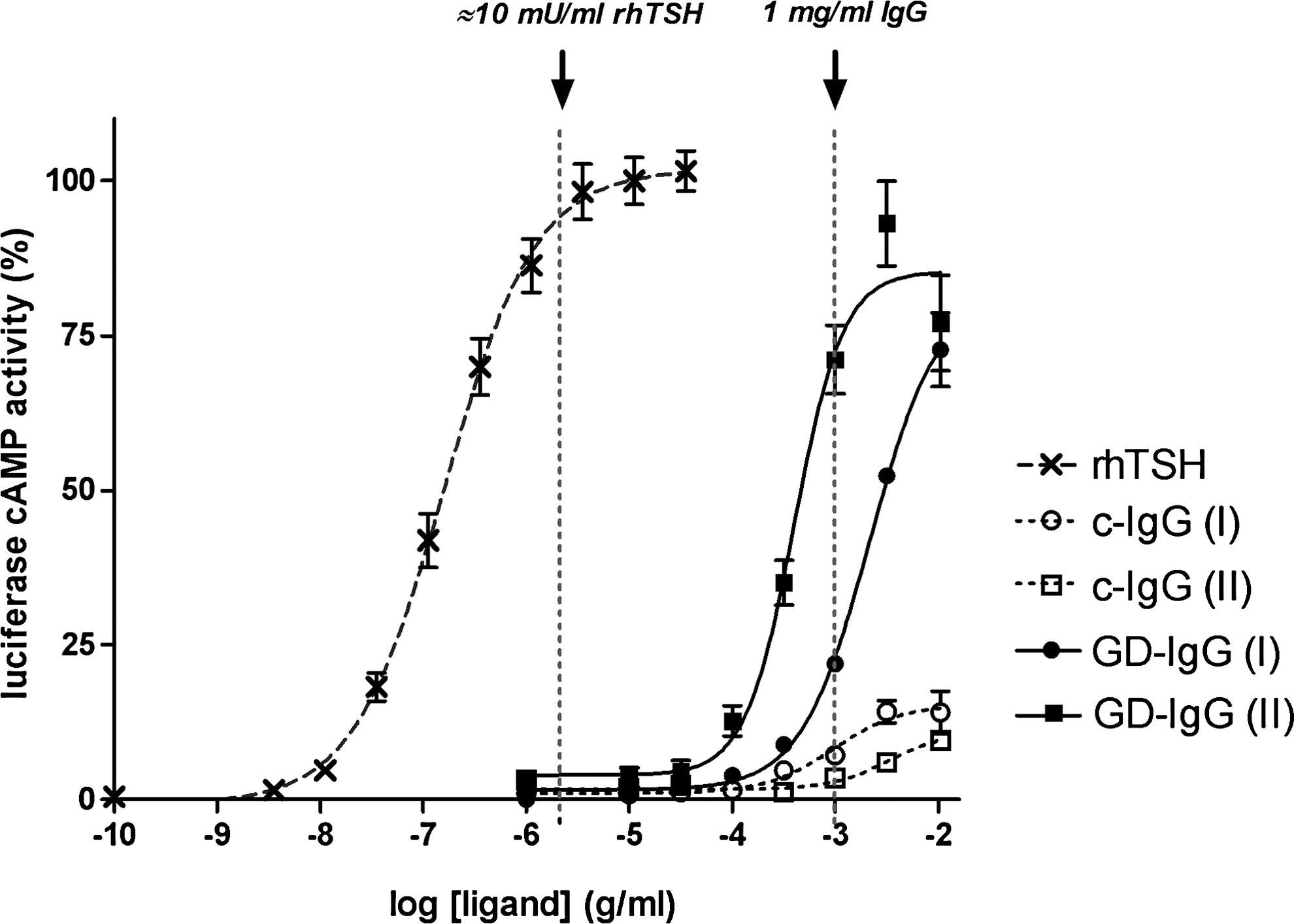
Stimulation of cAMP-responsive luciferase activity by increasing concentrations of rhTSH (X), c-IgG (○, □), and GD-IgG (•, ▪) in a hTSHR-CHO cell line with luciferase activity readout. Data are normalized to the luciferase activity after rhTSH; baseline was set at 0%, and maximal luciferase activation upon rhTSH was set at 100%. Shown are the dose–response curves for rhTSH, the two c-IgG, and the two GD-IgG batches. The dotted vertical line indicates the administration of 1 mg/mL IgG. Mean values of quadruple measurements ± standard deviation are shown. cAMP, cyclic adenosine monophosphate; GD-IgG, Graves' disease immunoglobulin G; rhTSH, recombinant human thyrotropin.
TSHR and HAS mRNA expression in orbital fibroblasts
We measured TSHR mRNA expression in orbital fibroblasts from five patients and all cultures expressed TSHR mRNA under basal conditions; however, expression levels were very low (∼10E-5 times lower) compared to housekeeping gene transcript levels. TSHR mRNA expression was not altered after 8 hours of incubation with 10 mU/mL rhTSH, but was moderately increased after 8 hours incubation with GD-IgG (p < 0.01; Fig. 2). HAS2 mRNA was most abundantly expressed under basal conditions in all orbital fibroblasts cell lines (n = 9) with transcript levels similar to that of the housekeeping gene. HAS1 and HAS3 mRNA expression was less abundant (∼100-fold lower compared to HAS2 mRNA expression), although still detectable in all cultures. HAS mRNA expression levels varied between different orbital fibroblast cell lines (data not shown).
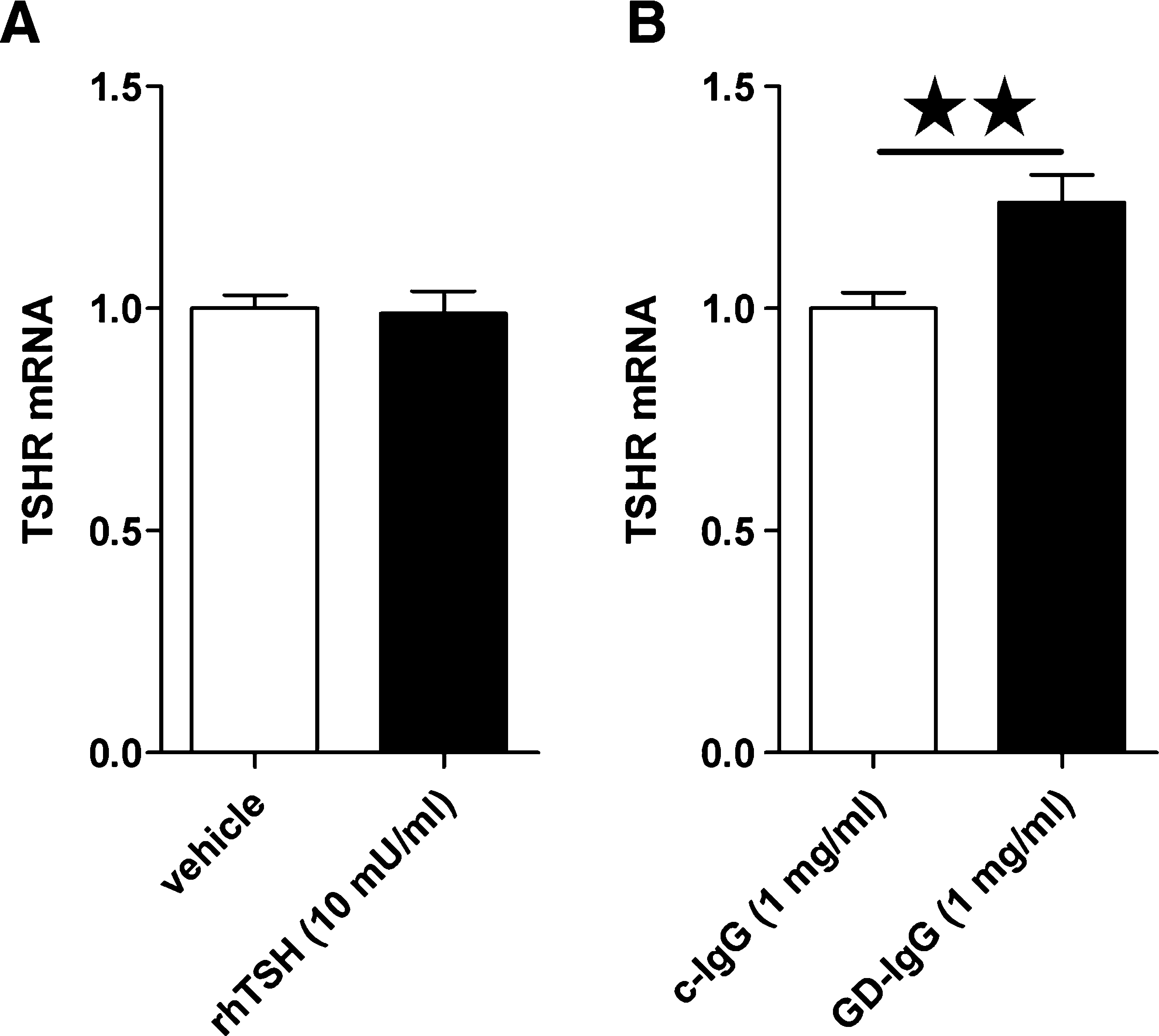
Effects of rhTSH, c-IgG, or GD-IgG on TSHR mRNA expression. TSHR mRNA expression (normalized to GAPDH) after incubation with vehicle or rhTSH (
cAMP production and HA synthesis after IL-1β in GO orbital fibroblasts
Stimulation with 10 ng/mL IL-1β resulted in a twofold induction of cAMP production by orbital fibroblasts (p < 0.01; Fig. 3A). HA concentration in the supernatant was induced twofold (all p < 0.01; Fig. 3B). mRNA expression of all three HAS isoforms was increased after 8 hours of incubation with 10 ng/mL IL-1β; HAS1, HAS2, and HAS3 were induced, respectively, 13-fold, 1.7-fold, and 2-fold (all p < 0.01; Fig. 3C). These findings indicate adequate responsiveness of the orbital fibroblasts used in our experiments.

Effects of IL-1β on cAMP production and HA synthesis. (
cAMP production and HA synthesis after FSK in GO orbital fibroblasts
Stimulation with 50 μM FSK induced a 40-fold induction of cAMP production (p < 0.01; Fig. 4A) in the orbital fibroblasts of all three patients. HA concentrations in the supernatant of the GO orbital fibroblasts (four patients) were 2.5-fold increased upon 48 hours of treatment with FSK (p < 0.01; Fig. 4B). FSK induced HAS1 mRNA expression levels markedly in all cultures (n = 5; p < 0.01; Fig. 4C). HAS3 mRNA expression was also moderately increased; however, HAS2 mRNA expression was unaltered.
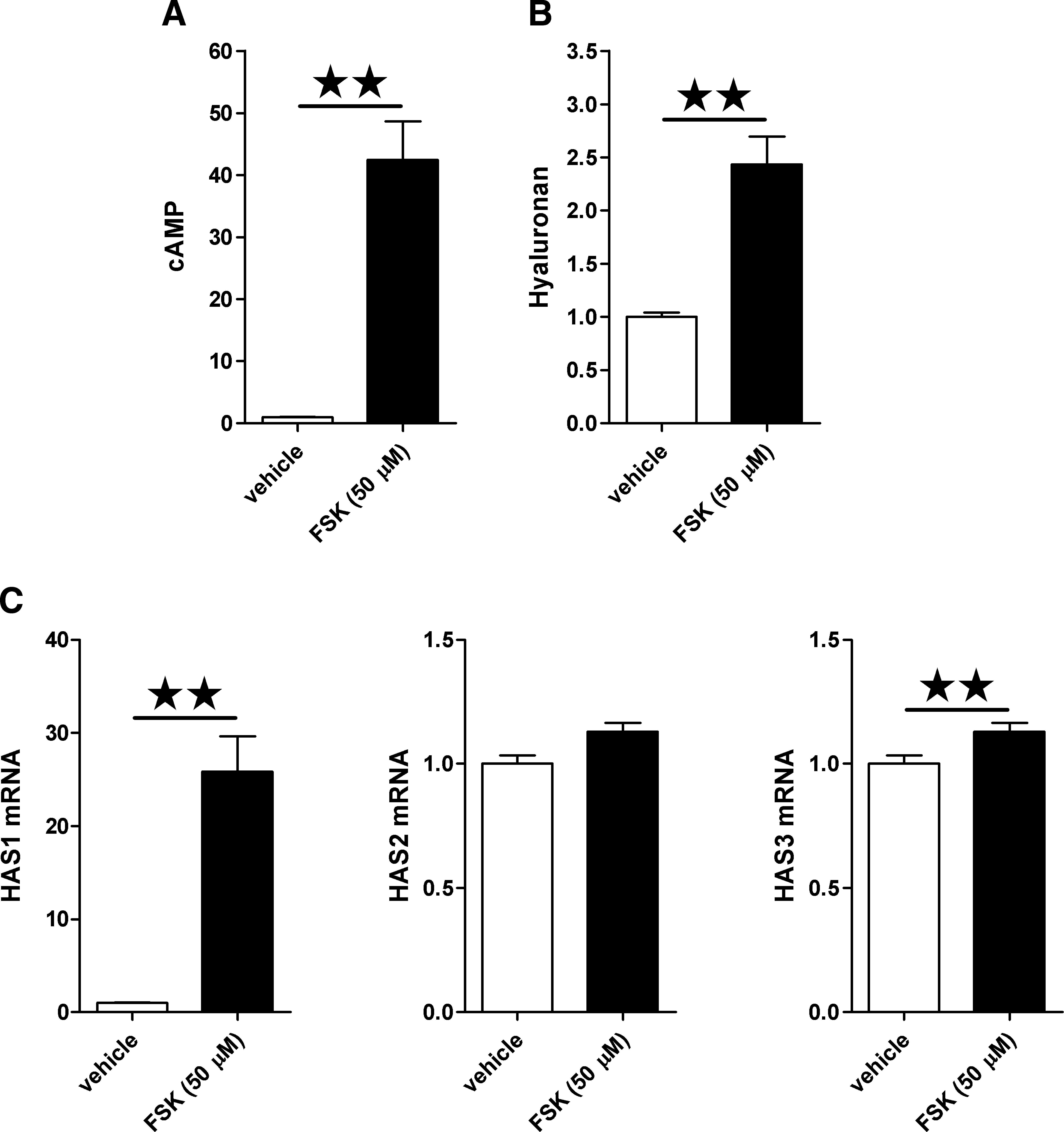
Effects of FSK on cAMP production and HA synthesis. (
cAMP production and HA synthesis after rhTSH in GO orbital fibroblasts
Orbital fibroblasts of all three patients tested showed an increased cAMP production by 10 mU/mL rhTSH. Orbital fibroblasts of one patient showed a 50% cAMP increase compared to 25% in both the other cultures. This strain came from the patient suffering from dysthyroid optic neuropathy due to severe GO not responding to corticosteroid treatment, but it was also the strain with the lowest number of passages (3) compared to the other cultures (both passage 5). In the pooled data, cAMP production was increased after rhTSH treatment (p < 0.01; Fig. 5A). HA concentrations did not differ in supernatants of GO orbital fibroblasts (three patients) after 48 hours of stimulation with 10 mU/mL rhTSH (Fig. 5B). In agreement, HAS 1, 2, and 3 mRNA expression was not altered in GO orbital fibroblasts (five patients) after stimulation with 10 mU/mL rhTSH for 8 hours (Fig. 5C).
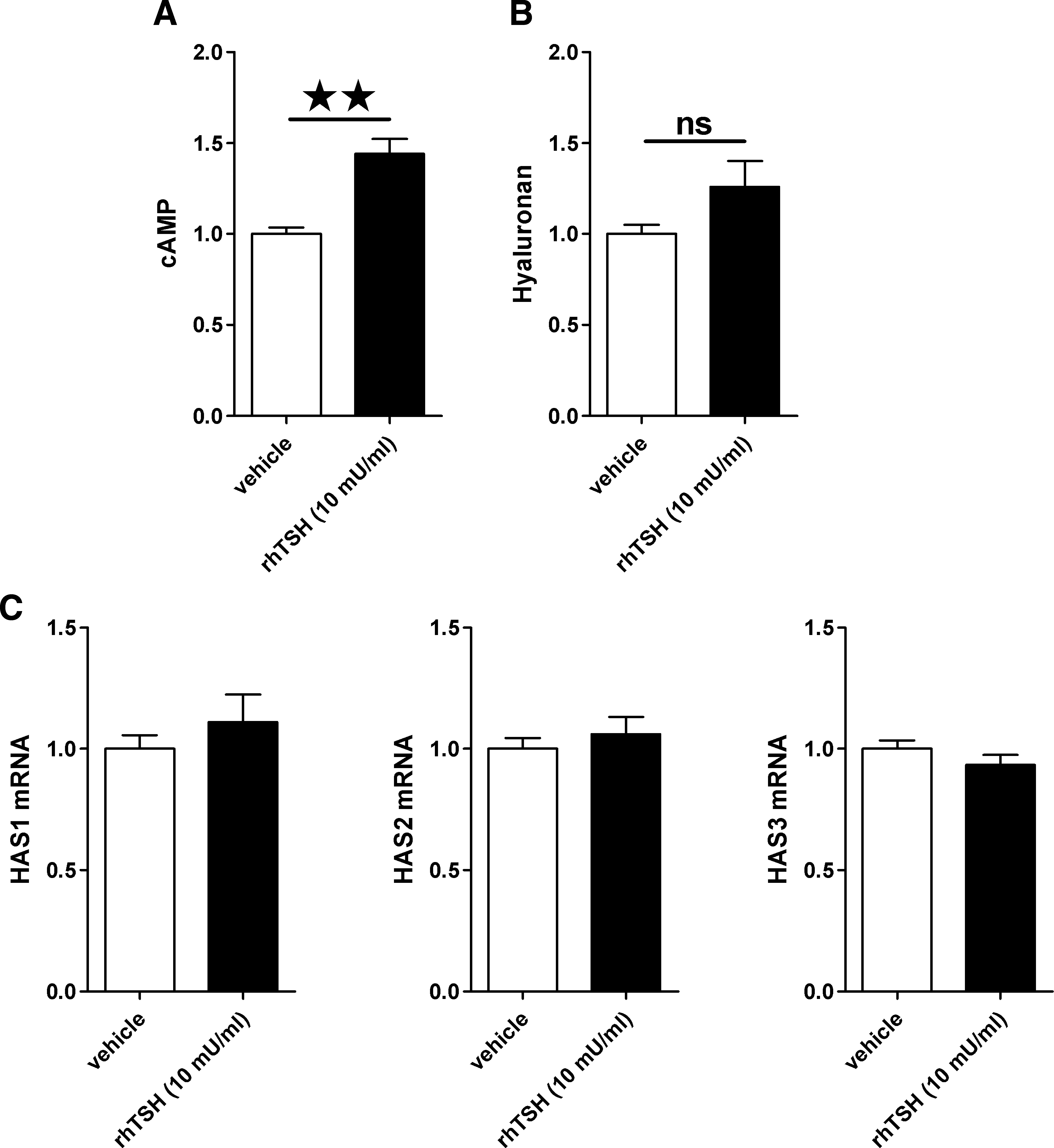
Effects of rhTSH on cAMP production and HA synthesis. (
cAMP production and HA synthesis after GD-IgG in GO orbital fibroblasts
Experiments with c-IgG and GD-IgG were performed with two different batches of c-IgG and GD-IgG. No differences between c-IgG batches I or II or GD-IgG batches I or II were found in the effects on cAMP synthesis, HA production, or HAS1–3 and TSHR mRNA expression in the nondifferentiated orbital fibroblasts (data not shown). GD-IgG (1 mg/mL) had no measurable stimulatory effect on cAMP production compared to c-IgG (1 mg/mL) in any of the three orbital fibroblast cultures (Fig. 6A). Stimulation of GO orbital fibroblasts (six patients) with 1 mg/mL GD-IgG for 48 hours did not result in increased HA concentrations in the supernatant of GO orbital fibroblast cultures (Fig. 6B). In agreement with unchanged HA secretion, HAS1, 2, or 3 mRNA expression was unaltered in GO orbital fibroblast cultures (n = 4 patients) after 8 hours of incubation with 1 mg/mL GD-IgG (Fig. 6C). Additionally, in the performed dose–response experiment no differences between c-IgG and GD-IgG were observed on cAMP production, HAS mRNA expression, or HA synthesis after administration of 100 ng/mL, 1 μg/mL, 10 μg/mL, 100 μg/mL, and 1 mg/mL GD-IgG in the two orbital fibroblast cell lines (data not shown).

Effects of GD-IgG on cAMP production and HA synthesis. (
Discussion
In this study, we evaluated the effects of rhTSH and GD-IgG purified from patients with Graves' disease, on the cAMP signaling pathway in nondifferentiated orbital fibroblasts of several GO patients and examined the effect on HAS mRNA expression and HA synthesis.
We propose that nondifferentiated GO orbital fibroblasts best approach the fibroblast population in the early stage of GO. We are aware that the orbital fibroblast cell lines used in the present study are derived from patients with inactive GO and thus after the inflammation was extinguished (and in three patients after corticosteroid treatment). Naturally, the most accurate representation of the orbital fibroblast population present in the orbit during the early stage of GO in vitro would be derived from orbital fatty tissue obtained immediately after initiation of the autoimmune process. Unfortunately, this is impracticable.
The results of our study suggest that activation of the cAMP signaling pathway plays a role in HA induction by GO orbital fibroblasts, as strong activation of this signaling pathway by FSK markedly induces HAS1 mRNA expression in GO orbital fibroblasts and increases HA content in the supernatant. However, mild activation of the cAMP signaling pathway by rhTSH does not result in HA synthesis in these cells; further, TSHR-stimulating GD-IgG do not induce cAMP production nor HA synthesis in GO orbital fibroblasts. Changes in HA content were in all cases in line with changes in HAS mRNA expression.
Our findings are in keeping with the observation that incubation with dibutyryl cAMP (Bt2 cAMP) increased HA content in a dose-dependent manner in orbital fibroblasts, not showing any effect on HA synthesis after low concentrations Bt2 cAMP (25). Moreover, Zhang et al. (29) very recently demonstrated that incubation with TSH or a monoclonal thyroid-stimulating antibody (IRI-SAb2) did not result in altered HA in the culture medium of orbital fibroblasts from GO patients, which is in perfect agreement with our study. In the same study, HA was modestly increased after similar treatments in normal orbital fibroblasts, implying a slight disparity between GO orbital fibroblasts and naive non-GO orbital fibroblasts in vitro.
The GO orbital fibroblast cultures used in the present study were capable to induce HA synthesis in response to IL-1β via upregulated mRNA expression of all three mammalian HAS isoforms as described previously (6), and all GO orbital fibroblasts expressed TSHR mRNA. Although TSHR expression levels were low as reported before (40), stimulation of the TSHR by rhTSH induced cAMP production, demonstrating functionality. TSHR activation by rhTSH (10 mU/mL) did, however, not result in increased HA synthesis, which is in agreement with previous findings of Smith and Hoa for 1 mU/mL rhTSH (28).
We then studied the effect of GD-IgG on cAMP production and HA synthesis in GO orbital fibroblasts. GD-IgGs selected to use in this study stimulated the production of cAMP at a concentration of 1 mg/mL in CHO cells expressing the human TSHR, demonstrating a marked ability of these heterogeneous GD-IgG preparations on TSHR activation. This is important because sera of patients with Graves' disease consist of functionally heterogeneous TSHR antibodies with both TSHR-blocking and TSHR-stimulating activities (16,41,42). Stimulation of the TSHR in GO orbital fibroblasts with 1 mg/mL GD-IgG did not induce cAMP production or HA synthesis; no differences in HAS1, HAS2, and HAS3 mRNA expression or secreted HA were observed.
Smith and Hoa (28) showed increased HA concentration in supernatants of cultured orbital fibroblasts of unspecified passage number of a single patient with GO after incubation with 100 ng/mL GD-IgG. The same group also showed upregulation of two T cell chemoattractants, the C-C chemokine regulated upon activation normal T cell expressed and secreted (RANTES) and IL-16 in supernatant of GO orbital fibroblasts after incubation with 100 ng/mL GD-IgG (28,43). In contrast, in our experimental setting, stimulation with 100 ng/mL GD-IgG did not induce CRE-luciferase production in the hTSHR-CHO cell line. Moreover, in the performed dose–response experiment with c-IgG and GD-IgG in two orbital fibroblast cell lines, no alterations in cAMP production, HAS mRNA expression, and HA synthesis were observed after 100 ng/mL, 1 μg/mL, 10 μg/mL, 100 μg/mL, and 1 mg/mL IgG. It is possible that the discrepancy between the two studies is caused by differences in the GD-IgG preparations. The GD-IgGs used in this study were IgGs purified from patients with Graves' disease with exceptionally high-serum TSHR-antibody titers (88 and 256 U/L) and selected on the basis of their potency to activate TSHR in the hTSHR-CHO cell line. We purified GD-IgG using protein G, whereas Smith and Hoa used protein A. However, proteins A and G have approximately the same binding activity to most human IgG subclasses, although protein G binds more strongly to human IgG3, and protein A binds somewhat better to IgG1, IgG2, and IgG4 λ (44).
It is known that excessive or persistent TSHR stimulation by TSH or TSHR-stimulating antibodies may lead to a decrease in TSHR signaling (45), either by desensitization or downregulation of TSHR. GD-IgG has been described to desensitize, but not downregulate, TSHR on thyroid cells in vitro through excessive TSHR stimulation (22). It has therefore been suggested that a concentration of TSHR-stimulating antibodies in sera of Graves' disease patients of 100 ng/mL or even lower are optimal to maintain hyperthyroidism, because then desensitization might fail to occur (46). However, in our TSHR-expressing CHO cell line we found increased cAMP production after 6 hours of stimulation with 1 mg/mL GD-IgG, but not with 100 ng/mL GD-IgG. Moreover, TSHR mRNA expression in the orbital fibroblasts was moderately upregulated after 8 hours of stimulation with 1 mg/mL GD-IgG, suggesting that 1 mg/mL GD-IgG in vitro does not desensitize or downregulate the TSHR in GO orbital fibroblasts.
Another pathway that has been implicated to play a role in HA synthesis in orbital fibroblasts by GD-IgG is via activation of the IGF1R (28). Our orbital fibroblast cell lines expressed IGF1R mRNA (data not shown); however, our finding that HA production was not altered upon GD-IgG indirectly indicates that the GD-IgG did not activate the IGF1R, or that IGF1R activation did not result in HA synthesis.
Orbital fibroblasts in vitro are reported to produce excessive amounts of HA after stimulation with several cytokines, such as IL-1β, interferon gamma, leukoregulin, and tumor necrosis factor-α (7,47 –49), which supports the idea that cytokines are involved in orbital tissue remodeling by orbital fibroblasts in GO (49). Our findings suggest that cytokines play a more dominant role in HA synthesis by orbital fibroblasts of patients with GO than GD-IgG. The activation of the cAMP pathway may play a role in this process, as IL-1β increases cAMP production in orbital fibroblasts. However, other pathways are likely to be involved, because IL-1β increases mRNA expression of all three HAS isoforms, whereas FSK does not increase HAS2 mRNA expression.
TSHR expression is upregulated after adipocytic differentiation of orbital fibroblasts in vitro (5,12). It is plausible that in these cells, rhTSH and GD-IgG have a greater effect on cAMP production. It would be interesting to study whether increased activation of the cAMP signaling pathway by rhTSH and GD-IgG in differentiated GO orbital fibroblasts would be followed by increased HA synthesis.
We conclude that strong activation of the cAMP signaling pathway by FSK in nondifferentiated orbital fibroblasts of GO patients results in increased HA synthesis. However, GD-IgG with TSHR-stimulating activity did not induce a measurable cAMP response or HA synthesis in nondifferentiated orbital fibroblasts, which do express, though at a low level, functional TSHR. Our in vitro results based on this experimental cellular system do not support a predominant role for TSHR-stimulating GD-IgG in the accumulation of orbital GAGs, but lend support for the assumption that orbital cytokines play an important role in the increased HA synthesis by orbital fibroblasts, at least in the early stages of GO.
Footnotes
Acknowledgments
We thank Dr. J.M. Ruijter (Department of Anatomy and Embryology, Academic Medical Center, Amsterdam) for his expert advice on PCR quantification; Ing. J. Kwakkel (Department of Endocrinology and Metabolism, Academic Medical Center, Amsterdam) for her expert technical assistance; and J. Brands and T. van Achterberg (Schering-Plough Research Institute, Oss, The Netherlands) for performing the cAMP measurements.
Disclosure Statement
C.J.J.Z., O.V.S., and A.B. have nothing to disclose. This work was supported by a grant from Schering-Plough (to E.F. and W.M.W.). C.J.K., M.E.G., M.P.M., and A.M.M.M. are employees of Schering-Plough Research Institute, Oss, The Netherlands.