
Abstract
Background:
Thyroid gland development and function are essential for life, and recent findings indicate the presence of stem/progenitor cells within the thyroid gland as a potential source of tissue regeneration and cancer formation.
Summary:
This review summarizes the current knowledge on early differentiation of thyroid cells from embryonic stem cells and highlights exciting concepts and recent novel findings on adult thyroid stem/progenitor cells in the normal thyroid gland and in thyroid cancer. Other potential sources and markers of stem/progenitor cells in the thyroid include bone marrow, microchimerism, and embryological remnant-derived multifocal solid cell nests. Finally, we discuss new therapeutic strategies that target thyroid cancer stem cells.
Conclusions:
Thyroid stem/progenitor cell populations are present in the normal and diseased thyroid gland. Advances in normal and cancer thyroid stem cell biology will be essential for future targeted therapies.
Introduction
Thyroid cancer is the most common carcinoma of endocrine glands, comprising ∼1% of all malignancies, with a 3:1 female/male ratio, and North America has one of the highest incidences of this cancer among all malignancies (5,6). Four types of thyroid carcinoma, namely papillary thyroid carcinoma (PTC), follicular thyroid carcinoma, anaplastic, undifferentiated thyroid carcinoma (UTC), and medullary thyroid carcinoma comprise more than 98% of all thyroid malignancies. Different and sometimes exclusive cytogenetic events and oncogenic mechanisms are effective in thyroid carcinoma of distinct histological types (7 –9).
Here, we review developmental processes in relation to thyroid stem/progenitor differentiation, comment on adult stem cells within the thyroid gland, and provide evidence for the presence of CSC in thyroid cancer.
Differentiation Markers in Thyroid Development
The two distinct endocrine cell populations of the thyroid gland develop from different origins: the thyroid hormone-producing follicular thyroid epithelial cells derive from the endoderm of the pharyngeal floor and the calcitonin-producing interfollicular C cells derive from neural crest cells within the ultimobranchial bodies (UBB) of the fourth pharyngeal arch (10,11). Thyroid organogenesis requires the specification of thyroid precursor cells from endoderm cells of the foregut, the subsequent migration and proliferation of thyroid precursors, the fusion of derivatives of the thyroid primordium and the UBB, and the final stage of functional differentiation (Fig. 1).

Summary of milestones of thyroid organogenesis in the mouse embryo until the beginning of functional differentiation (24,28,136,157). The equivalent human developmental stages according to Carnegie staging (CS) and/or weeks of development are shown in blue (158). Fgfr2, fibroblast growth factor receptor 2; Tg, thyroglobulin; TPO, thyroid peroxidase; TAZ (transcriptional coactivator with PDZ-binding motif ), a transcriptional regulator of the Pax8 and Titf-1 gene (159); TSH, thyroid stimulating hormone; TSHR, TSH receptor; T4, tetraiodothyronine or thyroxine; NIS, sodium/iodide (Na+/I−)-symporter.
In the mouse embryo, definitive endoderm originates as early as E6–E7.5 from the posterior epiblast, where gastrulation begins with the expression of fibroblast growth factor 8 (FGF8), brachyury, Wnt3, and eomesodermin (12 –16). Endoderm development in vertebrates is initiated by the transforming growth factor β (TGFβ)/nodal-related pathway (12,17) and gives rise to the epithelial lining of the gut and the respiratory system and all its derivatives, such as the thyroid, lung, liver, gallbladder, and pancreas. Canonical Wnt signaling was shown to promote the early expression of nodal ligands (12,18,19), and high levels of nodal signaling initiate definitive endoderm formation. Nodal activity in the epiblast is confined spatially and temporarily by expression of the nodal antagonists Lefty and Cer1 (20). In embryoid bodies (EBs), endoderm development can be induced by exposure to high levels of the TGFβ family member activin-A from a brachyury-positive cell population (21).
Among nodal targets, the homeodomain-like protein Mixl-1 and the transcriptional activators, GATA4 and GATA6, are key components in visceral endoderm derivatives. The high-mobility group (HMG)–box transcription factor (TF) Sox 17 and the hepatic nuclear factors Hnf3α (Foxa1) and Hnf3β (Foxa2) are temporarily expressed during thyroid organogenesis.
Thyroid organogenesis begins with the specification of distinct endodermal cells within the floor of the primitive pharynx on E8.5 (mouse) and formation of the thyroid anlage. Thyroid precursor cells are characterized by the unique coexpression of the four TFs, Titf1/Nkx2.1 (TTF1), Pax8, Foxe1 (TTF2), and Hhex, which are expressed simultaneously in thyroid precursor cells and in the differentiated follicular cells (22). The factors initiating thyroid specification of endoderm are unknown (Fig. 2). Although in vitro activation of the thyroid-stimulating hormone (TSH)/TSH receptor (TSHR) pathway regulates the expression of Titf1, Pax8, and Foxe1, their expression in vivo is independent of TSH/TSHR signaling (23,24) as shown in mouse models with impaired TSH signaling, such as Tshrhyt/Tshrhyt mice (25,26) or pitdw/pitdw mice (27). Studies with mouse mutants have shown that the four TFs have distinct roles during early organogenesis: Titf1 and Pax8 are essential for the survival of thyroid precursor cells, Foxe1 promotes migration of thyroid precursor cells, and Hhex may be important as a repressor of differentiation and for the maintenance of gene expression of Ttif1, Foxe1, and Pax8 in the thyroid primordium (Table 1).
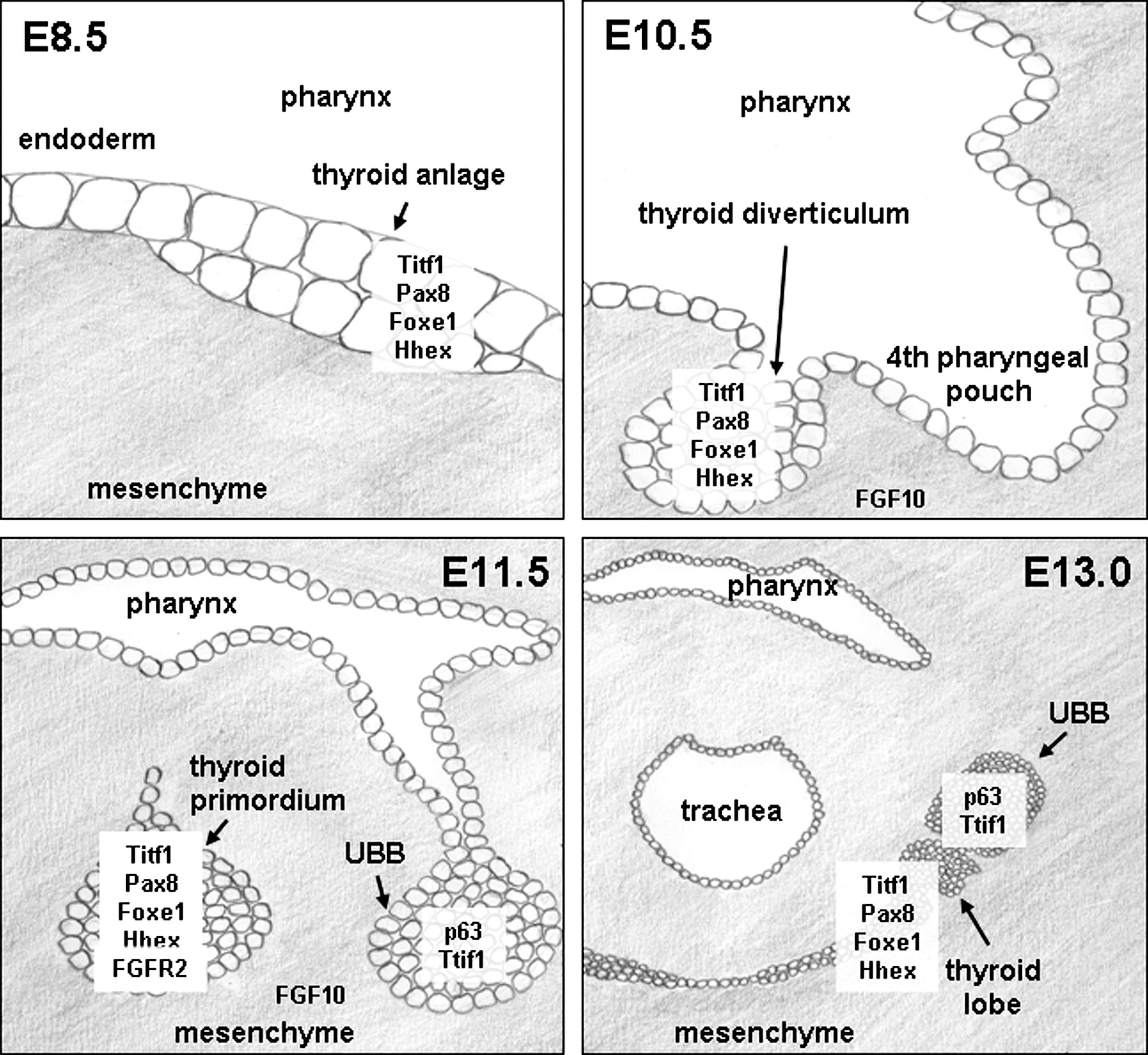
Schematic drawings of early thyroid development. At E8.5, the thyroid anlage is formed as a thickening of the endoderm layer on the bottom of the primitive pharynx by specification of endoderm cells for the thyroid lineage. Factors initiating this specification process are unknown. Thyroid precursor cells coexpress the four transcription factors Titf-1/Nkx2.1, Pax8, Foxe1, and Hhex during development and functional differentiation and in adult thyroid follicular cells. At E10.5, proliferation of thyroid precursor cells has shaped the thyroid diverticulum. Migration of the thyroid precursors away from the endoderm lining temporarily forms the thyroglossal duct, which regresses at E11.5. At E11.5, the thyroid primordium has further descended and the UBB has formed from the fourth pharyngeal arch. At E13.0, the thyroid primordium has formed two lobes, reached the final location in front of the trachea and contacts the UBB. FGF10, fibroblast growth factor 10; FGFR2, FGF receptor 2; UBB, ultimobranchial body.
UBB, ultimobranchial bodies; TPO, thyroid peroxidase; Tg, thyroglobulin.
Factors derived from the surrounding mesenchyme are essential for thyroid development. The growth factors epidermal growth factor (EGF) and fibroblast growth factor (FGF) promote thyroid cell proliferation and repress differentiation in vitro (28). FGFs upregulate the expression of Titf1 in the early anterior foregut (29) and FGF10 was shown to downregulate Sox2 and p63 in the endoderm of the future esophagus (30). Mouse embryos deficient for the FGF receptor 2 IIIb (31) and FGF10 knockout mice (32) show thyroid agenesis, suggesting that thyroid precursor cells respond to FGF10 from the surrounding mesenchyme (32,33). Interestingly, E-cadherin is expressed during all stages of thyroid organogenesis and N-cadherin is absent at all times (34), indicating that the migration of thyroid precursor cells occur independent of epithelial to mesenchymal transition.
The UBB contains two different cell types: Titf1+ cells (majority of cells) and rare p63+ cells (35). P63+ cells can survive without Titf1 and give rise to vesicular structures lined by p63+ basal-type epithelial cells. Solid cell nests (SCNs) in the mature human thyroid gland are considered developmental remnants of the UBB and contain p63+, Bcl-2+, and telomerase+ main cells (36 –38).
Thyroid Stem/Progenitor Cells from Embryonic Stem Cells
Pluripotent embryonic stem (ES) cells can develop three-dimensional embryoblast-like EBs under suitable culture conditions. Cells within the EB can spontaneously differentiate into more advanced embryonic stages of all lineages (39,40). Apart from three major advantages of the EB model, namely (i) the characterization of precursor cell functions, (ii) the in vitro assessment of the effect of null mutations of specific genes or the testing of lethal gene knockouts on ES cell viability, and (iii) the suitability as a cell source for tissue engineering, this EB model has now also been used to promote early thyroid cell differentiation (41).
CCE ES cells and TSHR+/− ES cells (W9.5) with a green fluorescent protein (GFP)-neomycin cassette in mouse TSHR exon 1 coding sequence resulting in TSHR promoter-mediated GFP expression (GFP+) have been employed in ES studies on thyroid differentiation (41 –43). Generating thyroid-like cells from CCE and TSHR+/− ES cell-derived EBs is a multistep/multifactorial process. It involves initial induction of endoderm differentiation by limited exposure of EBs to serum or activin-A, a TGFβ family member and nodal ligand (21,42 –44). When EBs are allowed to grow out as an adherent cell layer, thyroid-like cells appear and express follicular thyroid markers PAX8, Na+/I− symporter (NIS), functional TSHR, thyroid peroxidase (TPO), and thyroglobulin (Tg).
Activin-A induces human ES cells to undergo differentiation into definitive endoderm cells (21,45). Upon 5 days of exposure to activin-A, EBs derived from TSHR+/− ES cells (W9.5) were reported to show markedly decreased transcript levels for the Oct-4 and Rex-1 stem cell markers and significantly increased gene activity for the endoderm markers Gata-4, CXCR-4, Foxa-2, and α-fetoprotein (44). Similar treatment also resulted in ∼2% of GFP+ and functional TSHR+ ES cells within the EB expressing, identifying activin-A as an inducer of NIS+ , PAX8+ , and TSHR+ thyroid progenitor cells in the absence of TSH (44).
Recently, the importance of step-wise exposure of ES cells to a coordinated sequence of endocrine factors for thyroid-specific determination and thyrocyte maturation has advanced our understanding of in vitro thyroid differentiation from pluripotent ES cells. Although activin-A increased the expression of the endoderm-specific genes Foxa2 and Sox17, its transient effect on TSHR expression at day 5 was extended for an additional day in the presence of TSH, the endocrine factor known to affect TSHR gene regulation during EB differentiation (41,43). TSH-mediated thyroid lineage induction and determination also included a temporary upregulation of NIS in replated cells from EBs under serum-free conditions. However, both activin-A and TSH failed to sustain a stable differentiated thyrocyte phenotype (43). Propagation of EB-derived mature thyrocytes with the expression and partial colocalization of the follicular thyroid markers TSHR (number of GFP+ cells) and NIS and the appearance of Tg required the additional presence of insulin and/or insulin-like growth factor-I at later stages during EB culture (43).
Thyroid Stem/Progenitor Cells in the Adult Thyroid Gland
Resident thyroid stem/progenitor cell populations
Although the ES/EB system is a suitable in vitro model to recapitulate steps in follicular thyrocyte differentiation and despite sophisticated new culture conditions, like the activin-A–TSH–insulin/insulin-like growth factor-I treatment regime, the thyroid-like cell progenitors generated are usually transient, variable, and too low in cell number (1–2%) for additional functional studies. The adult thyroid gland has been implicated as another source of stem/progenitor cells. Being a low-proliferation tissue (46,47), the high frequency of benign and malignant nodular disease in the thyroid gland is not sufficiently and convincingly explained by genetic predisposition, iodine deficiency, mutagenesis, perturbed endocrine actions, and/or ligand–receptor deregulation events alone (48,49). Earlier results from transplantation experiments in nude mice also suggest the presence within nodular goiter tissues of autonomously growing thyroid-derived cells (50). Recently, first evidence for the presence of Oct4+ adult stem cells and precursor cells of endoderm origin with an Oct4+, GATA-4+, HNF4a+, PAX8+, and Tg− phenotype was obtained in tissues and cultured cells derived from adult human goiter (51). Based on the expression of a functionally active ABCG2, fluorescent-activated cell sorting (FACS) revealed a small side population (SP; 0.1% of total cells) in cultured thyrocytes from human goiter tissues with a high nuclear/cytoplasmic ratio and an ABCG2+, Oct4+ phenotype. This SP cell fraction failed to express the endoderm markers GATA-4, PAX8, and HNF4a, or the typical thyroid differentiation markers TSHR, TPO, NIS, and Tg which were all detected in the non-SP fraction (52). The SP cell population remained viable in monolayer cultures for 14 days, but only in nonadherent suspension cultures and in the presence of EGF and FGF did they create spheres. These thyrospheres were composed of up to 5% of a proliferative ABCG2+, Oct4+ (SP) or GATA-4+, HNF4a+ (non-SP) thyroid stem/progenitor cell population (52). TSH completely inhibited sphere formation. The addition of serum-induced TSHR gene activity and subsequent dual action of serum plus TSH over a 3-week period further enhanced TSHR expression and caused the sphere-derived cells to differentiate into adult PAX8+, TSHR+, NIS+, TPO+, and Tg+ thyrocytes with loss of stem and endoderm markers. Importantly, these cells showed the hallmarks of differentiated thyrocytes by being capable of generating thyroid follicle-like structures and showing TSH-dependent iodine uptake (52).
Clusters of clonally derived human Oct4+, Nanog+ cells with large nuclei and small cytoplasm similar to thyroid stem cells described by Lan et al. (52) were recently also reported to generate spheres in the presence of basic FGF and EGF but in the absence of serum and TSH (53). When these thyrospheres were seeded into a three-dimensional collagen matrix adopted for differentiated thyrocytes (54,55), ∼50% of spheres created T4-secreting follicle-like structures of Tg+ cells lining the lumen and peripherally located TPO+ cells, but TSHR was absent from these follicles (53). The formation of follicles in the presence of serum was TSH independent but required the absence of EGF and FGF. Spheres cocultured with the neuroblastoma cell line LAN5 (56) displayed larger morphology, showed expression of the neuronal marker β-tubulin III, and also initiated adipocyte differentiation, indicating a high level of plasticity of the cells within the thyrospheres (53). When injected into SCID mice, thyrocytes failed to generate tumors (57).
In the adult mouse thyroid, stem cells expressing the ES cell genes Oct4 and nucleostemin also reside in the SP which makes up 0.3–1.4% of the total population of thyroid cells (58 –60). Similar to the human thyroid SP, mouse ABCG2+ cells represent a heterogeneous population with ∼50% expressing Sca1+ (61,62) thyroid stem/progenitor cell marker of an otherwise ABCG2+, Oct4+, nucleostemin+, c-kit−/CD117−, and CD45− phenotype. Similar to the human thyroid SP, mouse thyroid SP cells were devoid of transcripts encoding for PAX8, TPO, and Tg (58). However, when the SP was further subdivided into SP1 and SP2 fractions resembling FACS-gated areas of higher and lower Hochst33342 efflux, respectively, the SP2 of ABCG2+, Oct4+ cells was weakly positive for TSHR and Titf1, suggesting a thyroid progenitor lineage to be present in SP2 (58). Small numbers of ABCG2+ cells (∼1%) were localized to the interfollicular space and coexpressed Sca1 or the mesenchymal marker vimentin. Interestingly, a few calcitonin-positive C cells were also ABCG2+, suggesting the presence of a C-cell progenitor lineage among the ABCG2+ thyroid cells in the mouse (58).
Other Sources of Stem/Progenitor Cells in the Thyroid
Bone marrow
In many high- and low-proliferating organs, stem/progenitor cells are replenished by the bone marrow (63,64). Patients who had received hematopoietic stem cell transplantation have an increased risk for thyroid carcinoma (65,66). Young age at transplantation was the strongest risk factor, followed by irradiation, female sex, and graft-versus-host disease. The lack of CD45 and/or CD117 hematopoietic marker expression in SP cells derived from the human (53) and mouse adult thyroid (58), respectively, may indicate either of two things: (i) stem cells in the thyroid are not replenished by the bone marrow, or (ii) as a result of low cell turnover rates in the thyroid (46,47), human CD45+ and mouse CD45+, CD117+ stem cells derived from bone marrow lose the expression of both markers while residing in the thyroid for longer times.
Microchimerism
Microchimerism describes the bidirectional trafficking and persistence of a small number of allogeneic cells in a genetically different organism (67). This fetal–maternal exchange of cells is a common phenomenon in likely all placental species (68 –72). The detection of allogeneic fetal cells in maternal tissues, called fetal microchimerism, is frequently observed as early as 4–6 weeks of gestation (71,73,74) and at term virtually all pregnant women contain low numbers of circulating fetal cells (75 –77). Long-term survival of fetal cells, for example, CD34+ hematopoietic stem/progenitor cells, CD45+ cells, and desmin+ mesenchymal stem cells, has been reported in maternal tissues (77 –80). Male fetal cells were detected in follicular adenoma and the normal thyroid gland of mothers (81 –84) and these cells frequently populate the thyroid tissues of autoimmune Graves' disease and Hashimoto thyroiditis (85). A subset population of human CD34+, CD45− stem/progenitor cells (3–19%) with high plasticity was recently detected in seven spheroid lines generated from the normal thyroid and perinodular normal thyroid tissues (53). Of these cell lines, three were tested Tg+ and two were Oct4+ and Nanog+ (53), but the potential fetal microchimeric origin of these stem/progenitor thyroid cells was not reported.
Embryological remnant-derived SCNs
Nonfollicular, multicellular, single, or multifocal SCNs are frequently (61–89%) detected in the thyroid and were postulated to be embryological remnants of the UBB and the thyroid diverticulum (36,86 –88). SCN, thyroglossal duct cysts, and lingual thyroid remnants are believed to harbor pluripotent stem cells (36,86,87). SCNs are composed of two distinct cell types, main cells and C cells, and display a wide variety of histopathological features making it sometimes difficult to distinguish SCNs from primary, secondary, or metastasizing squamous metaplasia or carcinoma, thyroglossal cust, PTC, medullary microcarcinoma, and C-cell hyperplasia (36,86). Separate from SCNs but often (81%) admixed with SCNs are mixed follicles which consist of main-like cells and regular follicular cells lining a follicular lumen (36,89). Two recent reports highlighted the importance of the basal/stem cell marker p63 as a suitable marker for main cells of SCN, main-like cells within mixed follicles, and a rare cell population in thyroglossal duct cyst and thyroid isthmus, the latter being a derivative of the thyroid diverticulum/thyroglossal duct (37,88). The nuclear TF p63 shares structural and functional similarities with the tumor suppressor p53. Alternative promoter usage and splicing events generate multiple p63 isotypes with N- and C-terminal alterations, respectively, some of which are known to counteract the actions of p53 (90,91). P63 is a marker of basal and progenitor epithelial cell layers in numerous tissues (91,92). The major defects in limb and craniofacial development and deficient epithelial regeneration due to depletion of a basal progenitor cell population in mice deficient in p63 also point toward a role for p63 in stem cell maintenance and the initiation of squamous cell differentiation (93,94). P63 and the cytokeratin (CK) 34βE12 were identified as specific markers for main cells of SCNs and within mixed follicles, but these cells lacked the thyroid markers Titf-1, Tg, calcitonin, or calcitonin-related peptide (CRP) (38,86). By contrast, C cells of SCNs displayed a P63−, Galectin-3+, Titf-1+, calcitonin+, CRP+, and chromogranin+ phenotype which is thought to reflect progressive parafollicular C-cell differentiation (37). Galectin-3 was identified as a suitable marker to distinguish SCN from normal parafollicular C cells (95).
Thyroid Cancer Stem/Progenitor Cells
Emerging concepts in thyroid carcinogenesis
Thyroid cancer is a diverse entity with respect to causative factors, thyroid cell types involved, histopathological phenotypes at different frequencies, and clinical outcome. Based on epidemiological, genetic, and animal data, a multistep, progressive thyroid carcinogenesis model suggested an orchestrated step-wise dedifferentiation from benign thyroid disease to differentiated thyroid carcinoma to anaplastic thyroid cancer as a result of accumulating somatic mutations in mature thyroid cells (8). This multistep thyroid carcinogenesis hypothesis has been challenged for a number of reasons: (i) a low turnover rate of the thyroid gland of 1 per 5–10 years or six to eight renewals per lifetime limits the accumulation of multiple mutations (46,47); (ii) unique genetic alterations specifically found only in a certain type of thyroid cancer question the multistep carcinogenesis model (96); (iii) transgenic rearranged during transfection/papillary thyroid carcinoma (RET/PTC) mice develop PTC but mature thyrocytes fail to transform when overexpressing the same fusion protein (97) (98); (iv) the monoclonal origin of thyroid cancer cells combined with strikingly different histomorphology may reflect distinct developmental morphogenesis programs to be active in specific thyroid cancer cells (99); (v) specific differentiation marker profiles, like oncofetal fibronectin, in PTC and UTC suggest common fetal ancestry (100 –103); (vi) the frequent presence of SCN within the thyroid points toward an involvement of developmental factors in thyroid cancer (86,88). The fetal cell thyroid carcinogenesis hypothesis postulates a hierarchy of fetal cell remnants within the adult thyroid which may act as a source for specific thyroid CSC generating distinct thyroid cancer phenotypes (104,105) (Fig. 3). Such CSC and specific CSC markers have been identified for different cancers (4,106). Carcinogenic thyroid stem cells and more differentiated thyroblasts are postulated to generate UTC and PTC, respectively, whereas follicular thyroid carcinoma is proposed to be generated from both stem cell populations and further advanced prothyrocytes (104,107). The degree of malignancy in thyroid cancer is likely higher with CSC derived from more immature stem cell populations (107). A modified hierarchical scheme of thyroid stem cell development incorporates SCN-derived embryonic remnant cells (p63+, CEA+, and CK34βE12+) and, supported by histopathological findings, postulates a bipotential Titf1+ and TSHR+ precursor as the cellular source for specific progenitors of the follicular (Titf1+ and PAX8+) and C-cell lineage (P63− but possibly Galectin-3+ and Titf-1+) (86,88,102).
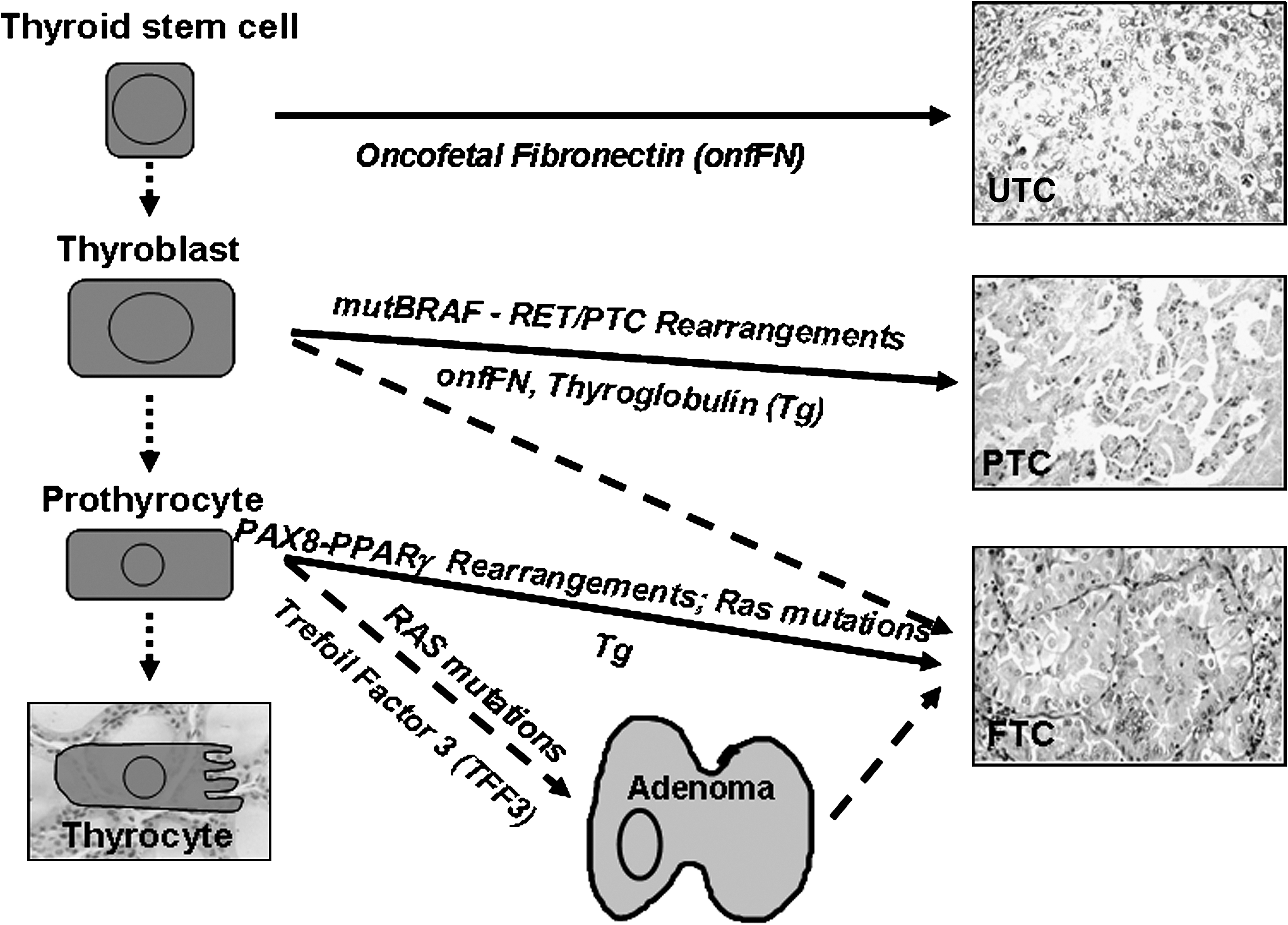
Schematic representation showing a developmental hierarchy of fetal thyroid stem/progenitor cells as a potential cause of specific follicular thyroid cancer types [modified according to Takano (104,107)]. Examples of mutations associated with and factors expressed in certain thyroid cancer types are depicted. Trefoil factor 3 (TFF3) may be used to distinguish foci of thyroid TTF3+ adenoma from TTF3weak/− FTC (160). FTC, follicular thyroid carcinoma; PTC, papillary thyroid carcinoma; UTC, undifferentiated anaplastic thyroid carcinoma.
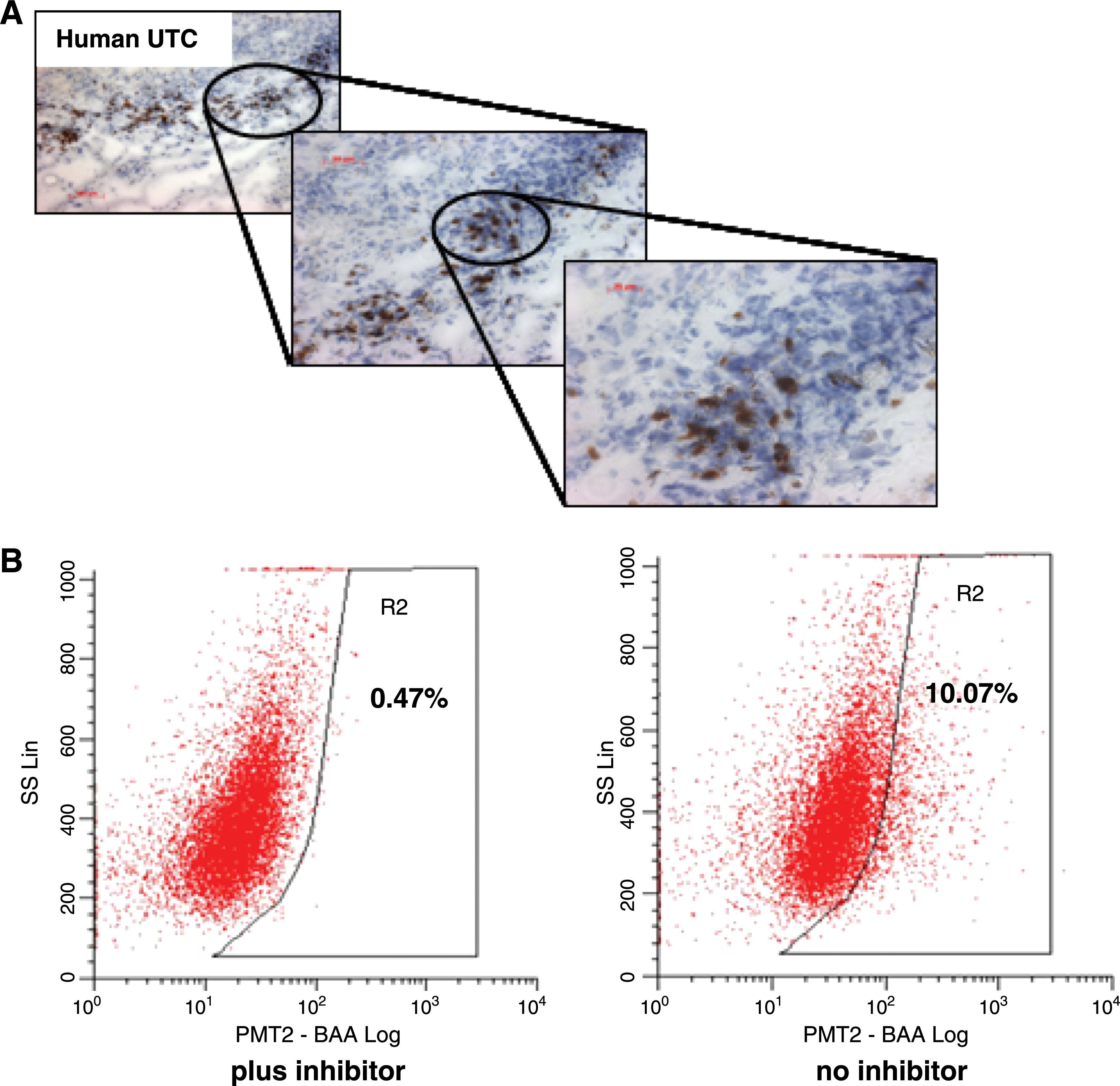
No CSCs have so far been isolated from thyroid cancer tissues, but we were able to detect Oct4+ cells within human UTC tissues (Fig. 4A). The isolation of CSC-like cells enriched in ABCG2+ SP or CD133+ cell populations has recently been reported using cultured human thyroid cancer cell lines. Comprehensive microarray chip analysis revealed genes related to the Wnt and Notch signaling pathways and known to play important roles in normal stem cell development to be upregulated in the SP fraction of ARO cells (4,108). However, both SP and non-SP fractions generated nude mouse tumors, indicating that the non-SP fraction still contained CSCs (108). Recently, CD133 (prominin-1) has been implicated as a specific marker of CSCs in the human anaplastic thyroid cancer cell lines ARO, KAT-4, and FRO (109,110). In comparison to CD133− ARO, the CD133+ ARO population showed increased expression of Oct-4 and Titf1 and significantly higher resistance to chemotherapy-induced apoptosis against chemotherapeutics used to treat UTC, such as cisplatin, doxorubicin, and etoposide (109). Increased Oct-4 expression in the CD133+ ARO was confirmed in a recent study which also showed a strong increase in TSHR mRNA and further enhancement in Oct-4 and TSHR gene activities and presence of CD133 upon TSH-induced proliferation of CD133+ ARO (110). CD133+/high ARO subpopulation revealed higher Oct-4 and TSHR expression levels than CD133+/low or CD133− ARO. When injected subcutaneously into NOD/SCID mice, CD133+/high ARO repeatedly formed aggressive tumors at 10-fold lower cell numbers than CD133+/low ARO, with CD133− ARO being unable to generate any tumor xenografts (110). Strong immunoreactivity for CD133 specific for human UTC tissues suggests an in vivo role for this five-transmembrane domain glycoprotein (110).
(
Although the reported results on UTC-derived CSC are interesting, it should be noted that in all three studies the main experimental work was done with the ARO81-1 cell line (108 –110). A recent DNA profiling analysis of 40 human thyroid cancer cell lines has raised concerns about the ARO cell line as a valid thyroid cell model. ARO81-1 and KAT-4, also used in one of the studies (109), were reported to be likely of nonthyroid origin and a result from a cross-contamination with the human colon cancer cell line HT-29 (111). For ARO81-1, this conclusion was drawn based on the results from two independent labs (111).
We used a different FACS-based functional approach to identify CSCs within the human UTC-8505C cell line, which is considered unique among the 40 thyroid cell lines tested (111). The ALDEFLUOR assay is based on the fact that human normal and cancer mammary cells with increased aldehyde dehydrogenase (ALDH1) activity were found to have stem/progenitor properties (112). Stem cell populations in multiple myeloma and acute leukemia also show increased ALDH1 activity (113,114). The ALDH1 substrate BAAA is specifically metabolized by cells harboring ALDH1 to create a fluorescent dye which remains trapped within the cells, thus facilitating positive selection in FACS. The specificity of the enzymatic reaction is determined using the specific ALDH1 inhibitor diethyl-aminobenzaldehyde. Employing this method, we could demonstrate a 17–38% ALDH1+ cell fraction in UTC-8505C which was almost completely inhibited in the presence of diethyl-aminobenzaldehyde, suggesting the presence of a resident stem/progenitor population in human UTC-8505C (Fig. 4B).
Other cell types involved in thyroid cancer
Recent evidence shows an inverse association between pregnancy, microchimerism, and cancer (72,115). Parous women with fetal microchimerism are significantly less likely to develop cancer than parous women not harboring fetal cells (116); survival times and therapeutic response rates are better in microchimerism-positive patients (117). Fetal cells acquired during pregnancy and potentially of stem/progenitor phenotype have been identified in thyroid cancer tissues (82,118). Although their function is not yet known, evidence from skin, breast, and thyroid cancer favors the involvement of these fetal cells in repair mechanisms rather than a role as CSC in cancer propagation (118 –120). Nevertheless, as recently shown for Kaposi sarcoma and evidenced by the blood vessel-forming capacity of fetal progenitor cells, a role for microchimeric cells in tumor formation and/or tumor propagation should also be considered (121).
The persistence within the adult thyroid of embryological remnants forming SCNs and mixed follicles and the pluripotent undifferentiated p63+ stem/progenitor cells they harbor have been implicated in the pathogenesis of Hashimoto autoimmune thyroiditis and thyroid cancer (88,122,123).
New Therapeutic Strategies for Thyroid Cancer
There is no treatment that effectively targets the CSC compartment in thyroid cancer. Current antithyroid therapies, such as surgery, chemotherapy, and radiation, destroy rapidly growing differentiated thyroid tumor cells, thus reducing tumor mass, but leave behind cancer initiator cells. An ideal treatment regime should kill differentiated cancer cells, and at the same time, specifically, selectively, and quickly destroy thyroid CSCs to avoid toxic side effects to other cell types, and counteract the mutagenic evasive potential of these CSCs. In SP the presence of active transmembrane ABC transporter family members, multidrug resistance transporter 1 and breast cancer resistance protein 1/ABCG2/MRP, not only facilitates the efflux of DNA-binding dyes like Hoechst 33342 but can also promote chemoresistance through efflux of anticancer drugs, such as mitoxanthrone, gemcitabine, doxorubicin, and 5-fluorouracil, from cells with (cancer) stem cell activity (124,125). Such SPs have been identified in numerous cancers, including normal and neoplastic thyroid cells (52,108). Targeted inactivation of the ABCG2 may increase resistance of CSCs through upregulation of the transporter as shown for neuroblastoma and CD133+ brain tumor stem cells (125,126). The Wnt/β-catenin pathway has important roles in stem cell renewal and chemoresistance of CSCs. Deregulation of this pathway is associated with thyroid cancer (127). Shown to target β-catenin and cause reduced invasiveness and proliferation of thyroid cancer cells (128), the selective tyrosine kinase inhibitor imatinib mesylate (Gleevec, Novartis, NY; STI571) was ineffective as monotherapy in UTC patients (129). Future advances in targeting CSCs critically depend on new stem cell isolation protocols to identify suitable markers for thyroid CSCs, and such cells isolated from existing thyroid cancer mouse models may provide important clues as to the potential CSC subtypes (Table 2). This will aid in the discovery of novel therapeutic agents and strategies that effectively target thyroid cancer stem/progenitor populations. At the same time, new antagonists directed at epidermal growth factor receptor (EGFR), vascular endothelial growth factor receptor (VEGFR), v-raf murine sarcoma viral oncogene homologue B1 (BRAF), rearranged during transfection (RET), cyclooxygenase 2 (COX-2), or vascular endothelial (VE)-cadherin and novel, more targeted delivery systems possibly involving coated multipurpose nanoparticles will be useful to block the growth of differentiated tumor cells (130,131).
PTC, papillary thyroid carcinoma.
Conclusions
The recent exciting discoveries in thyroid stem cell and thyroid CSCs research mark the dawn of a new era with potentially widespread impact on our understanding of thyroid pathophysiology, carcinogenesis, and treatment. Understanding the complexity of stem cells and their malignant CSC counterpart will undoubtedly be an essential step in the development of more targeted, selective, and individualized treatment regimes for thyroid cancer.
Footnotes
Acknowledgment
The authors express their gratitude to the following grant agencies: Natural Science and Engineering Research Council of Canada and Manitoba Health Research Council (to T.K. and S.H.K.) and Deutsche Forschungsgemeinschaft (to C.H.V.).
Disclosure Statement
The authors declare that no competing financial interests exist.