
Abstract
Background:
Both expression of the thyrotropin receptor (TSHR) and the production of hyaluronan (HA) by orbital fibroblasts (OF) have been proposed to be implicated in the pathogenesis of Graves' ophthalmopathy (GO). HA is synthesized by three types of HA synthase. We hypothesized that TSHR activation by recombinant human TSH (rhTSH) and TSHR-stimulating Graves' disease immunoglobulins (GD-IgGs) via induced cyclic adenosine monophosphate (cAMP) signaling increases HA synthesis in differentiated OF from GO patients.
Methods:
Cultured human OF, obtained during decompression surgery from 17 patients with severe GO, were stimulated in vitro to differentiate into adipocytes. Differentiation was evaluated by phase-contrast microscopy. The differentiated OF were stimulated by rhTSH or by TSHR-stimulating GD-IgG. We measured cAMP using a biochemical assay, HA synthase mRNA expression by quantitative polymerase chain reaction, and HA in the supernatant by enzyme-linked immunosorbent assay.
Results:
All differentiated OF cultures expressed higher levels of TSHR mRNA than nondifferentiated OF cultures. Stimulation by rhTSH induced a marked cAMP response in 11 of 12 differentiated OF cultures, but no measurable HA response in all but one differentiated OF cultures. By contrast, stimulation by GD-IgG induced a moderate cAMP response in a number of differentiated OF cultures, but a marked HA response in the majority of differentiated OF cultures.
Conclusion:
Stimulation of differentiated OF by GD-IgG, but not by rhTSH, induces HA synthesis in the majority of patients, suggesting that in most patients TSHR-mediated cAMP signaling does not play a pivotal role in GD-IgG-induced HA synthesis in differentiated OF cultures.
Introduction
Inflammation may drive fibroblasts to differentiate into adipocytes in various tissues (2,8). Under appropriate culture conditions, a subgroup of OF, termed preadipocytes or lipofibroblasts, has been shown to differentiate to adipocytes in vitro. This process may contribute in vivo to the accumulation of excessive adipose tissue in the orbit frequently seen in GO, and the differentiated OF may play a distinct role in the immunopathogenesis of GO (8 –12).
Functional thyrotropin receptor (TSHR) expression increases in OF after adipocytic differentiation (8,10,13). The TSHR is known to be the antigen that binds Graves' disease immunoglobulins (GD-IgGs) in thyroid follicular cells, leading to Graves' hyperthyroidism (14,15). TSHR gene expression was shown to increase in orbital tissue specimens of patients with GO in the active stage of the disease. As a correlation between TSHR autoantibody levels and clinical activity score of GO has been reported and in view of the prognostic value of TSHR antibody levels for the severity and the outcome of GO, the TSHR is considered a major autoantigen in the immunopathogenesis of GO (16 –18).
Binding of the TSHR by TSH or by GD-IgG results in activation of the adenylyl cyclase/cyclic adenosine monophosphate (cAMP) signaling pathway via G-protein-coupled pathways (19). In turn, cAMP induces HA synthesis in several cell types, among which OF (20,21). We and others have recently reported that strong activation of the cAMP pathway increases HA synthesis, mainly via upregulation of HAS1, in nondifferentiated GO OF, whereas TSH-mediated cAMP induction does not induce HA synthesis (21,22). In addition, we found that TSHR-stimulating GD-IgG did not increase cAMP production or HA synthesis in nondifferentiated OF. These findings suggested that orbital cytokines, such as IL-1β, play a more prominent role than GD-IgG to increase HA synthesis in the early stages of GO.
The aim of the present study was to evaluate the effect of recombinant human TSH (rhTSH) and GD-IgG on cAMP signaling and HA synthesis in differentiated OF from GO patients.
Materials and Methods
GD-IgG and control IgG isolation
As reported previously (22), we isolated IgG from sera of two GO patients (GD-IgG, batch I and batch II) and two healthy controls (c-IgG, batch I and batch II) using protein G Sepharose 4 Fast Flow (ProtG; Amersham Pharmacia Biotech Benelux, Roosendaal, The Netherlands), after obtaining informed consent. The GO patients had TSH-binding inhibitory IgG serum levels of 88 and 256 U/L, respectively. They had been found to have mild (clinical activity score [CAS] score 3/7) and moderately severe (CAS score 4/7) GO, respectively (23), and had not been treated with corticosteroids. At the time of the blood sampling, the GO was inactive in both patients. The healthy controls were TSH-binding inhibitory IgG-seronegative (<1.0 U/L).
To determine the ability of TSHR activation, purified c-IgG and GD-IgG were tested in a cAMP responsive (CRE) luciferase assay, a Chinese hamster ovary cell line stably transfected with the human TSHR, and a CRE luciferase gene as reported before (22). Briefly, the two GD-IgGs (1 mg/mL) showed activities comparable to the activity induced by 10−7 and 10−6 g/mL rhTSH (∼0.5–5 mU/mL), respectively. The c-IgG batches showed no or little CRE-luciferase activity. We chose to use the dose of 10 mU/mL rhTSH and 1 mg/mL IgG in subsequent experiments, as this concentration of rhTSH and GD-IgG elicited a marked effect on the CRE-luciferase activity in the TSHR-expressing Chinese hamster ovary cell line.
OF cell cultures and adipocytic differentiation
Orbital fat tissue explants were obtained from 17 patients (pat1–pat17) with severe GO undergoing orbital decompression surgery. Three patients (pat3, pat7, and pat11) received 1 g of corticosteroids within 24 hours previous to surgery. Fourteen patients underwent decompression surgery in the inactive stage of the disease and had not received corticosteroids or orbital irradiation in the previous 6 months. Collection of these tissues was approved by our Institutional Review Board.
OF cell lines were initiated as described previously (5,22,24) and cultured in six-wells plates in the culture medium (199 [BioWhittaker, Lonza, Verviers, Belgium] supplemented with 10 U/mL penicillin, streptomycin, fungizone, and 10% fetal calf serum [all from Cambrex, East Rutherford, NJ]). Fibroblast cultures were used between the third and ninth passage from initiation; therefore, not all cultures were subjected to all experiments.
For each experiment, 5 × 104 fibroblasts per well were grown to 90% confluence in six-well plates in ∼48 hours. Then adipocytic differentiation was carried out as reported previously (12). The differentiation protocol was continued for 2 weeks, replacing the medium every 3–4 days. Adipocytic differentiation was evaluated using phase-contrast microscopy, using an Olympus IX71 microscope equipped with a DP20 digital camera with a U-TV1X-2 camera adapter (all Olympus Life Science Europe GmbH, Hamburg, Germany). OF from six GO patients (pat2, pat3, pat4, pat5, pat16, and pat17) were also cultured in six-well plates in the culture medium to be able to evaluate TSHR mRNA expression in nondifferentiated and differentiated OF under basal conditions in these patients.
After adipocytic differentiation cells were incubated with 10 mU/mL rhTSH (Thyrogen®; Genzyme Therapeutics, Cambridge, MA), 1 mg/mL GD-IgG (batch I or II), or 1 mg/mL c-IgG (batch I or II), we also employed incubation with 50 μM forskolin (FSK; Sigma-Aldrich, Zwijndrecht, The Netherlands), a direct activator of the adenylyl cyclase/cAMP pathway, to evaluate the cAMP-production. In addition, we incubated the cells with IL-1β (10 ng/mL; Sigma-Aldrich Inc., St Louis, MO) as a positive control for HA synthesis, as IL-1β induces expression of the three HAS mRNA isoforms and HA production in nondifferentiated OF (5).
Oil Red O staining
After adipocytic differentiation of the OF according to the protocol described above, we performed Oil Red O staining in differentiated OF of four GO patients (pat2, pat3, pat4, and pat5). Most of the medium was removed and cells were incubated with 10% formalin for 5 minutes. Formalin was discarded and cells were incubated with fresh 10% formalin for at least 1 hour. The OF were washed with 60% isopropanol, dried completely, and then stained for 10 minutes with Oil Red O solution (2 mg/mL; Sigma-Aldrich, Zwijndrecht, The Netherlands). After rinsing with tap water, cells were observed by phase-contrast microscopy.
cAMP assay
cAMP measurements were performed in the supernatant of differentiated GO OF cultures (experiments with rhTSH: n = 12; GD-IgG: n = 12; IL-1β: n = 5; FSK: n = 9). Plates were preincubated in 750 μL of the differentiation medium containing 0.5 mM 3-isobutyl-1-methylxanthine (IBMX), for 30 minutes at 37°C, 5% CO2. Then, 10 ng/mL IL-1β, 50 μM FSK, 10 mU/mL rhTSH, 1 mg/mL c-IgG (batch I or II), or 1 mg/mL GD-IgG (batch I or II) was added to the IBMX-containing medium and cells were incubated for another 6 hours at 37°C, 5% CO2; each incubation was performed in three to six wells. We measured cAMP concentrations in culture supernatants in triplicate using the Alphascreen cAMP Assay (PerkinElmer, Groningen, The Netherlands) (25) and corrected for total amount of protein as described previously (22).
RNA isolation and quantitative reverse transcriptase-polymerase chain reaction
Experiments for determination of mRNA expression levels were performed in differentiated OF of nine GO patients (six wells/group), except for the experiments with IL-1β, which we conducted in five differentiated GO OF cultures. Cells were incubated with 10 ng/mL IL-1β, 50 μM FSK, 10 mU/mL rhTSH, 1 mg/mL c-IgG (batch I or II), or 1 mg/mL GD-IgG (batch I or II) for 8 hours. RNA isolation, cDNA synthesis, and polymerase chain reaction was performed and analyzed as reported previously (22).
HA assay
HA measurements were performed in culture supernatants of differentiated OF of GO patients (four wells/group) after stimulation with IL-1β (n = 8), FSK (n = 6), rhTSH (n = 7), and c-IgG and GD-IgG (n = 5) for 48 hours. At the start of the experiment, the differentiation medium was refreshed and cells were treated in quadruplicate. After 48 hours, supernatants were collected and stored at −20°C until use. The HA content was quantified in duplicate with a specific enzyme-linked immunosorbent assay kit (Echelon Biosciences, Salt Lake City, UT) per manufacturer's instructions, and data were corrected for total amount of protein per well, as described previously (22).
Statistical analysis
Normal distribution of the data was tested using the Kolmogorov–Smirnov test. To test statistical significance (p < 0.05) between groups of individual patients, we used Student's t-test if data were normally distributed or Mann–Whitney U test when data were not normally distributed. The pooled data were statistically analyzed using the nonparametric Wilcoxon Signed Rank Test for related samples. All tests were performed using SPSS (SPSS, Chicago, IL).
Results
Adipocytic differentiation
We subjected OF cultures obtained during consecutive orbital decompression surgery from 17 patients with GO to the differentiation protocol during 14 days and examined the differentiated cultures using phase-contrast microscopy. During the process of differentiation, the OF lost their elongated appearance and became rounded. In all differentiated cultures, a subset of OF showed macroscopic adipogenesis and contained numerous lipid droplets (Fig. 1A and B) as judged from Oil Red O staining (n = 4; data not shown). A considerable variation in the number and size of lipid droplet containing cells between the cultures was observed; the number of adipocytes ranged between ∼5% and 10% among patients.
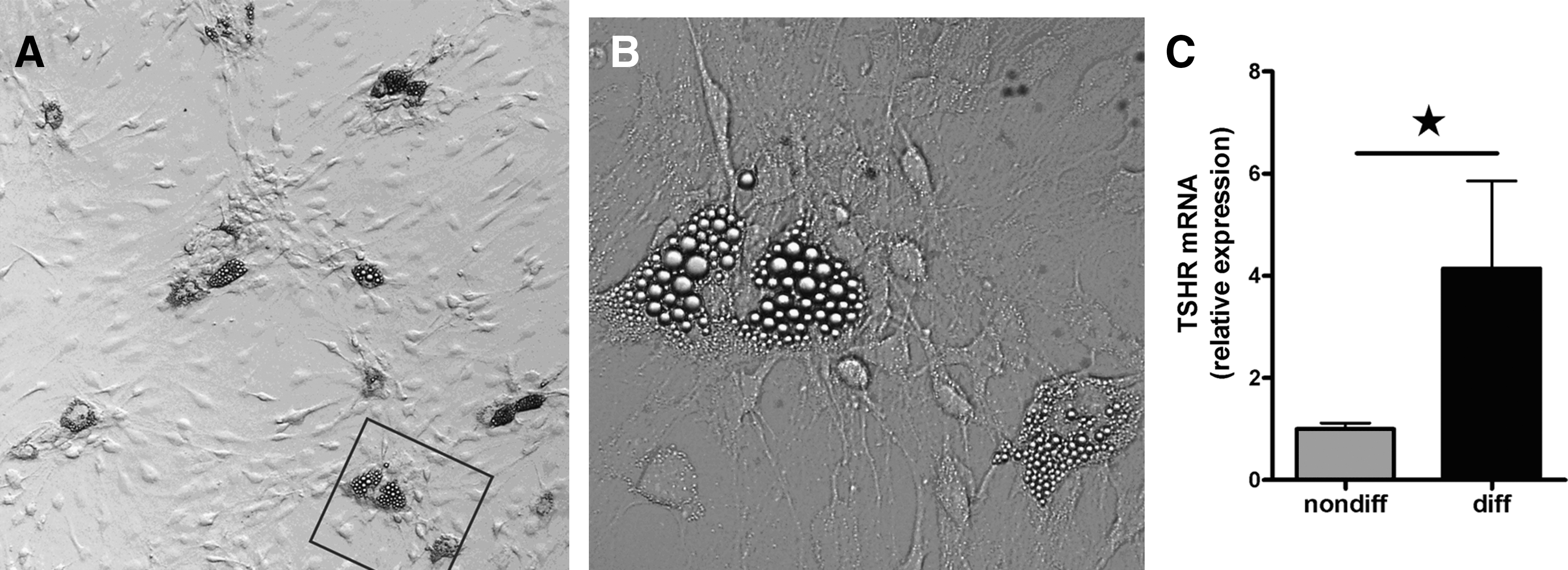
Cultured human orbital fibroblasts after adipocytic differentiation.
We studied TSHR mRNA expression in paired nondifferentiated and differentiated OF from six GO patients. TSHR mRNA expression significantly increased after adipocytic differentiation (p ≤ 0.05; mean ± standard error of the mean: nondifferentiated 1.0 ± 0.1; differentiated 4.1 ± 1.7; Fig. 1C).
Effect of rhTSH and GD-IgG on cAMP production in differentiated GO OF
To evaluate the effect of 10 mU/mL rhTSH on cAMP in differentiated OF, we first performed a dose–response experiment in differentiated cultures of three patients, adding a range of 0, 0.1, 1.0, 10, 100, and 316 mU/mL rhTSH. In all three cultures the EC80 was ∼10 mU/mL rhTSH, confirming our findings for rhTSH in the CRE luciferase assay (data not shown). In subsequent experiments, we therefore stimulated the differentiated OF with 10 mU/mL rhTSH and observed a marked variation in cAMP response (Table 1): the cAMP concentration significantly (p ≤ 0.05) increased in 10 of 12 differentiated OF cultures (the increase ranging from 1.3- to 7.8-fold), while cAMP concentration after rhTSH stimulation was similar to vehicle in two patients (Table 1 and Fig. 2a).
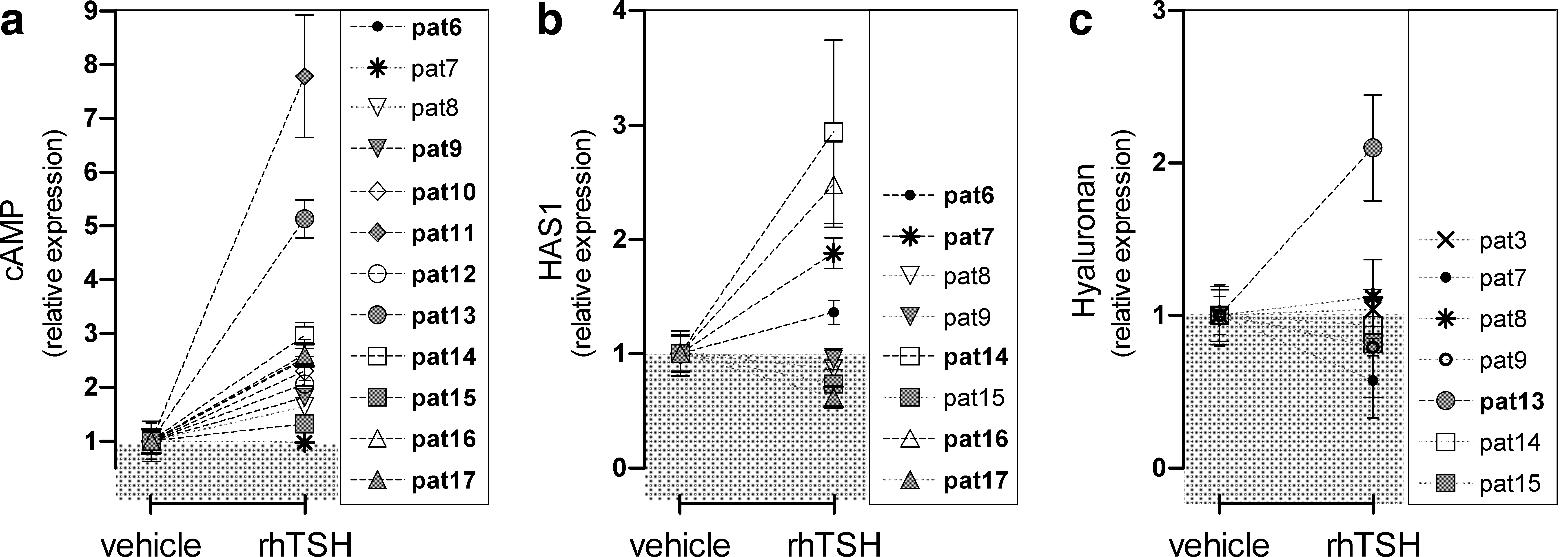
Effect of stimulation with recombinant human TSH (rhTSH) on cyclic adenosine monophosphate (cAMP) production and hyaluronan (HA) synthesis in differentiated GO orbital fibroblasts.
Data are normalized to the mean value of the control group, which was set at 1.
Fold changes compared to vehicle.
Fold changes compared to c-IgG.
Significantly increased cAMP production.
Significantly decreased cAMP production.
c-IgG, control immunoglobulin; cAMP, cyclic adenosine monophosphate; GD-IgG, Graves' disease immunoglobulin; rhTSH, recombinant human thyrotropin; SD, standard deviation.
Effects on cAMP concentration after stimulation with 1 mg/mL GD-IgG (batch I or II) were evaluated in the differentiated OF of 12 GO patients. Stimulation with GD-IgG resulted in a significant (p ≤ 0.05) cAMP increase in 4 of 12 differentiated OF cultures and in a significant cAMP decrease in 2 cultures (Table 1 and Fig. 3a). Stimulation of the differentiated OF of pat11 with rhTSH or GD-IgG resulted in the highest cAMP levels. This patient suffered from dysthyroid optic neuropathy and had received 1 g of corticosteroids the day previous to surgery.
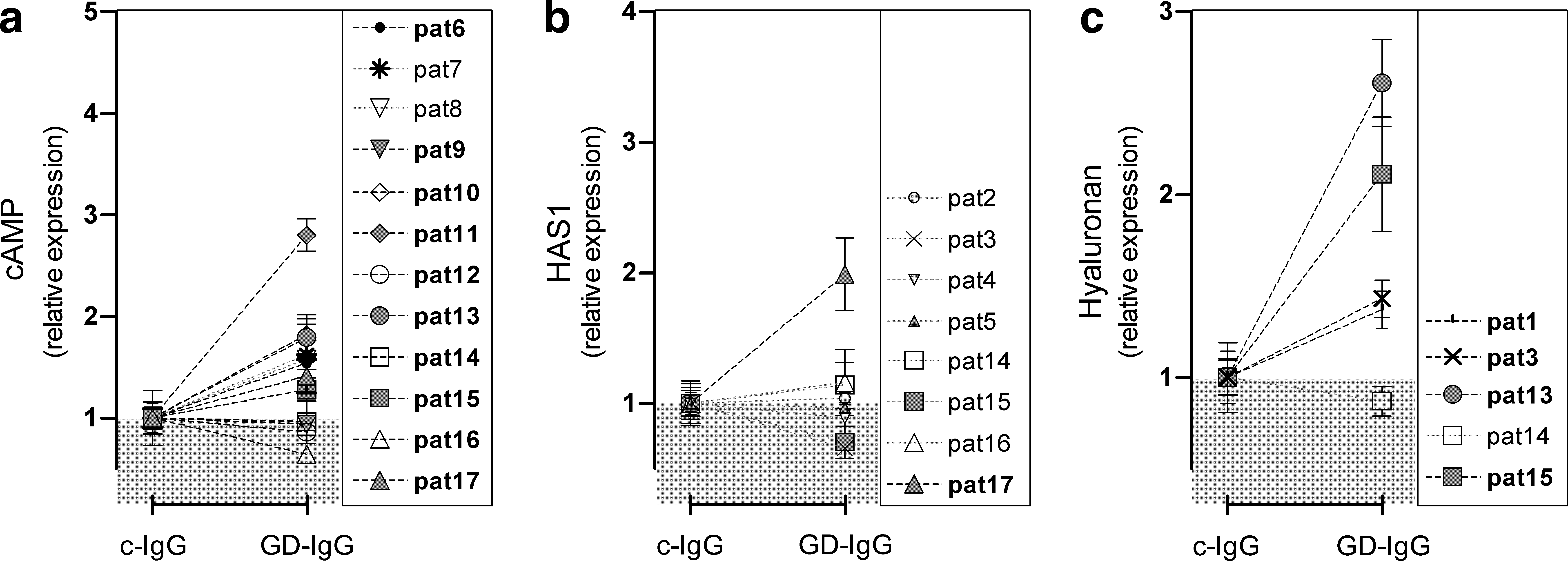
Effect of stimulation with Graves' disease immunoglobulin (GD-IgG) (compared to control IgG [c-IgG]) on cAMP production and HA synthesis in differentiated GO orbital fibroblasts.
When correlating TSH-mediated cAMP production and GD-IgG-mediated cAMP production in the 12 differentiated OF cultures, we observed a Pearson's correlation value of 0.727 (p = 0.007).
Effect of rhTSH and GD-IgG on HA synthesis in differentiated GO OF
rhTSH induced HAS1 mRNA expression in four of eight evaluated cultures (pat6, pat7, pat14, and pat16) and decreased HAS1 mRNA expression in three of eight cultures (pat8, pat15, and pat17) (all p ≤ 0.05; Fig. 2b). HAS2 and HAS3 mRNA expression remained unaltered after rhTSH (data not shown). HA content was increased twofold in the supernatant of one of seven differentiated GO OF cultures (pat13, p ≤ 0.05; Fig. 2c) after 48 hours of stimulation with 10 mU/mL rhTSH; unfortunately, we did not evaluate HAS transcript levels in this culture. In the other six differentiated OF cultures, rhTSH did not alter HA content in the supernatant (Fig. 3c).
Stimulation with GD-IgG increased HAS1 mRNA expression in one of eight differentiated OF cultures (pat17; Fig. 3b). HAS2 and HAS3 transcript levels did not alter after stimulation with GD-IgG (data not shown). The HA content in the supernatant increased in four of the five evaluated differentiated OF cultures after GD-IgG (pat1, pat3, pat14, pat15; all p ≤ 0.05; Fig. 3c).
Effect of IL-1β on cAMP production and HA synthesis in differentiated GO OF
To evaluate the HA synthesis, we stimulated our cultures with IL-1β, a very potent inducer of HA synthesis. Differentiated OF of five GO patients were stimulated with 10 ng/mL IL-1β, which resulted in a moderate cAMP increase (1.5-fold, p ≤ 0.05; Fig. 4a). HAS1 mRNA expression increased markedly (50-fold), HAS2 mRNA expression increased threefold, and HAS3 expression increased fivefold after stimulation with 10 ng/mL IL-1β (all p ≤ 0.05; n = 8 cultures; data not shown). Stimulation of differentiated GO OF with IL-1β for 48 hours resulted in a significant increase of HA concentration in the supernatant (3.3-fold; n = 8; p ≤ 0.01; Fig. 4b).

cAMP production and HA synthesis after interleukin-1β (IL-1β) in differentiated GO orbital fibroblasts.
Effect of FSK on cAMP production and HA synthesis in differentiated GO OF
To evaluate the effect of strong activation of the cAMP signaling pathway on HA synthesis, we stimulated our cultures with FSK. Stimulation of differentiated OF cultures of nine GO patients with 50 μM FSK resulted in markedly increased cAMP levels (200-fold; p ≤ 0.01; Fig. 5a). HAS1, HAS2, and HAS3 mRNA expression increased after 50 μM FSK (HAS1: 400-fold; HAS2: 5-fold; HAS3: 3-fold; all p ≤ 0.05; data not shown). Stimulation of differentiated GO OF with FSK resulted in a significant increase of HA concentration in the supernatant after 48 hours (11-fold; n = 6; p ≤ 0.05; Fig. 5b).
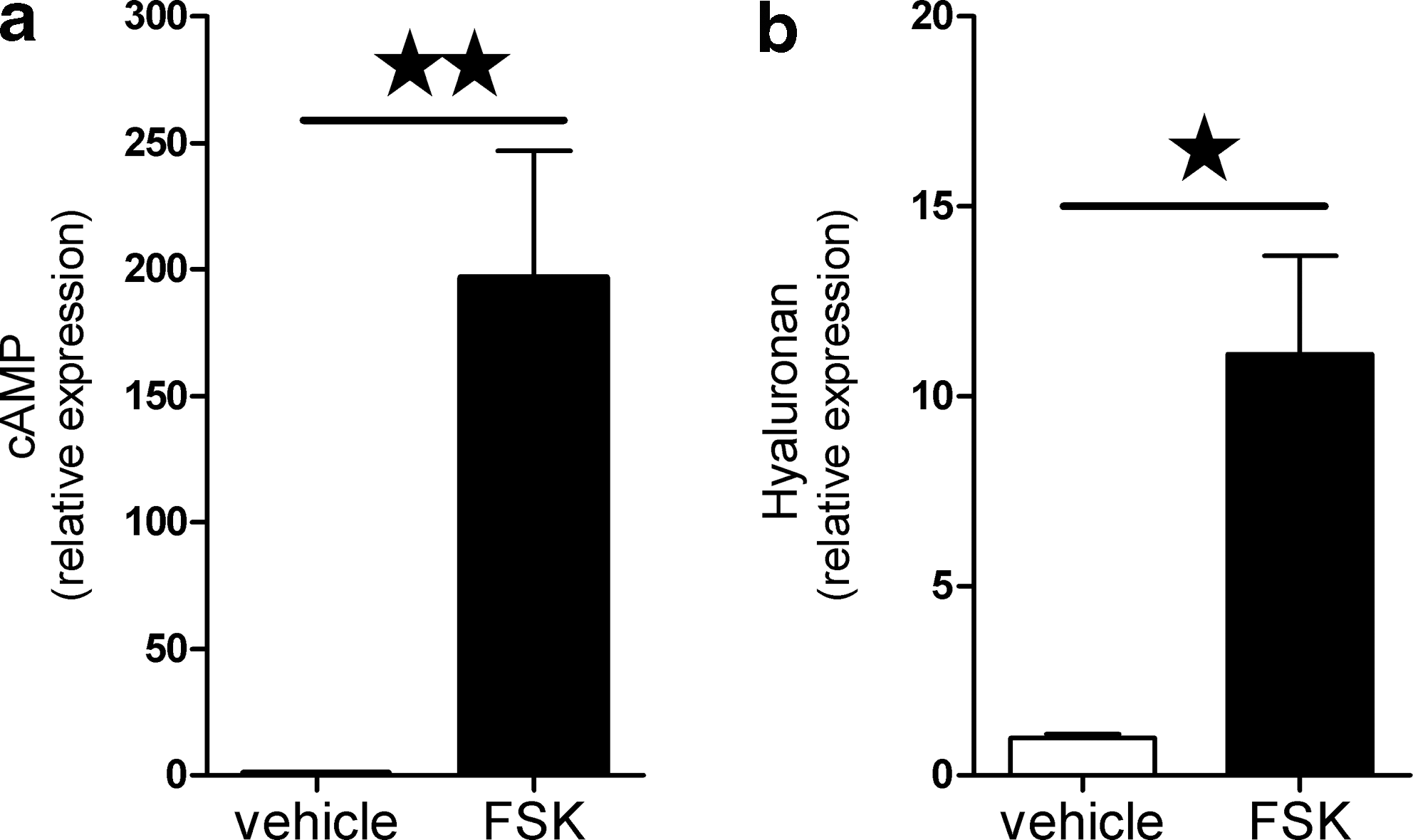
cAMP production and HA synthesis after forskolin (FSK) in differentiated GO orbital fibroblasts.
Discussion
Accumulation of adipose tissue is frequently seen in GO and there is increasing evidence that de novo adipogenesis may contribute to the pathogenesis of GO (8 –12,26). Feldon et al. demonstrated in vitro that activated human T lymphocytes can drive OF to differentiate to adipocytes, via activation of peroxisome proliferator-activated receptor-gamma (8). Functional TSHR expression is increased in OF after adipocytic differentiation (8,12,13,27). Moreover, Kumar et al. recently reported that TSHR activation in differentiated OF, by TSH or a human monoclonal TSHR-stimulating antibody (M22), resulted in increased expression of the adipocyte-related proinflammatory cytokine IL-6 (28). As IL-6 is involved in B-cell activation, this finding points to a distinct role for differentiated OF in the immunopathogenesis of GO.
In the present study, we evaluated the effects of rhTSH and TSHR-stimulating GD-IgG on the cAMP signaling pathway in differentiated OF. The effects of rhTSH and GD-IgG on HA synthesis in differentiated OF were evaluated for the first time.
The human OF cultures that were subjected to the adipocytic differentiation protocol (12) showed lipid droplet containing cells, confirming the presence of triacylglycerol. Variations in the size and the number of adipocytes (range 5%–10%) between the cultures were observed, a phenomenon reported previously by others (12). Stimulation with IL-1β resulted in induction of HA synthesis via upregulation of the three HAS isoforms. In addition, stimulation of differentiated OF with FSK resulted in marked cAMP production followed by increased HA synthesis, as previously observed in nondifferentiated OF (22). The present study confirms induction of functional TSHR in GO OF after adipocytic differentiation as TSHR mRNA expression was fourfold higher in differentiated OF compared to paired nondifferentiated cultures. This observation is in agreement with earlier reports (12,13). We observed marked differences in TSHR mRNA expression between the cultures, but our protocol did not allow us to correlate TSHR mRNA expression with the extent of adipocytic differentiation.
As reported by others, activation of the TSHR in differentiated OF by TSH resulted in increased cAMP production, also compared to nondifferentiated OF, demonstrating preserved functionality of the receptor (12,13,21,22,27,29). Moreover, we observed an effect of TSHR-stimulating GD-IgG on cAMP production in a number of GO OF cell lines, which does not occur in nondifferentiated OF (22). Notably, GD-IgG did not induce cAMP production in all differentiated cultures and we noted marked variation in cAMP concentration after stimulation with rhTSH or GD-IgG (Table 1). However, despite the limited number of differentiated cultures we found a significant correlation (R = 0.727, p = 0.007) between rhTSH- and GD-IgG-mediated cAMP production, suggesting that GD-IgG acted through the TSHR in the cultures that did increase cAMP production after stimulation with GD-IgG.
Stimulation with rhTSH did not result in altered HA synthesis in differentiated OF except for one culture (pat13), despite induced HAS1 mRNA expression in some cultures. By contrast, GD-IgG significantly induced HA synthesis in four out of five differentiated OF cultures, which was not associated with a cAMP response. We did not observe altered mRNA expression of any of the three HAS genes after stimulation with GD-IgG. As OF cultures do not have hyaluronidase activity, and are thus unable to degrade HA, the increase in HA concentration in the supernatant of differentiated OF must be preceded by increased HAS expression (30,31). We studied HAS mRNA expression after 8 hours as IL-1β, a potent inducer of HA synthesis, increased mRNA expression of all three HAS isoforms at this time point, but we cannot exclude that GD-IgG might have induced HAS mRNA expression at an earlier or later time point. We have previously reported that GD-IgG does not alter HA synthesis in nondifferentiated OF (22), which is in contrast to the responsiveness of differentiated OF to GD-IgG in the present study. At this time point we cannot exclude completely that the differences between nondifferentiated and differentiated OF are related to culture conditions, cell density, and proliferation, as these factors have been shown to influence HA synthesis (32,33).
Nevertheless, the finding that GD-IgG induces HA synthesis, despite lower GD-IgG-induced cAMP production compared to rhTSH-induced cAMP production, suggests that the TSHR-mediated cAMP response does not play a major role in GD-IgG-induced HA synthesis in differentiated GO OF. Hence, additional signaling pathways must be involved in GD-IgG-induced HA synthesis. It is possible that increased expression of TSHR in differentiated cultures is responsible for the effect of TSHR-stimulating GD-IgG on HA synthesis, via activation of alternative downstream signaling pathways. In agreement with this notion is a recent study by Morshed et al. showing that three potent stimulating TSHR-antibodies influence the intracellular extracellular-signal-regulated kinases (ERK1/2) pathway in primary thyrocytes (19). It is also possible that activation of the insulin-like growth factor-I receptor by GD-IgG is involved in GD-IgG-induced HA synthesis, as suggested by Smith et al. for nondifferentiated OF (34,35).
We studied GO OF cultures that were obtained during consecutive orbital decompression surgical procedures and present data of all performed experiments. We could not perform all analyses in all cultures; however, our findings clearly demonstrate that the differentiation of OF did not result in homogeneous cell populations, despite the fact that all differentiated cultures were subjected to the same differentiation protocol. Marked variations between the patients were observed in the number of lipid droplet containing cells, the increase in cAMP upon rhTSH, and the cAMP and HA induction by GD-IgG, illustrating that it is crucial to perform in vitro studies with OF from a variety of patients. Although the in vitro differentiation of OF cannot be compared directly to the in vivo situation in the orbit of patients with GO, it is tempting to speculate that in vivo OF are driven to differentiate to adipocytes by activated T cells and that differentiated OF may have a distinct role in the pathogenesis of GO.
In conclusion, the results of the present study suggest that TSHR-mediated cAMP signaling does not play a pivotal role in GD-IgG-induced HA synthesis in differentiated OF cultures. In addition, our findings suggest that GD-IgG induce HA synthesis in these cells via other signaling pathways, that is, through alternative signaling upon TSHR activation or via the insulin-like growth factor-I receptor.
Footnotes
Acknowledgments
We thank Jolanda Brands and Tanja van Achterberg (Merck Research Laboratories, Oss, The Netherlands) for excellent assistance in IgG purification and performing the cAMP measurements. This work was supported by a grant from MSD (Oss, The Netherlands) (to E.F. and W.M.W.).
Disclosure Statement
C.J.J.Z., E.F., O.V.S., M.P.M., W.M.W., and A.B. have nothing to disclose. C.J.K., M.E.G., and A.M.M.M. are employees of MSD.