
Abstract
Background:
Dendritic cells (DCs) are the most effective antigen-presenting cells and key regulators of immune response. The immunoregulatory properties of DCs strongly depend on the microenvironment in which DCs have been matured and activated. Thyroid hormones are an important part of this environment and regulate many vital processes including growth and cellular metabolism. The aim of the study was an analysis of the influence of thyroid hormones on blood DC subtypes ex vivo, including the surface expression of molecules involved in antigen presentation, costimulation, and maturation, as well as on functional properties of DCs in vitro.
Methods:
Blood samples for the quantitative and phenotypic analysis of peripheral blood plasmacytoid and myeloid DC subtypes were collected from thyroidectomized patients at two time points: (i) at the time of the so-called stimulation with endogenous thyrotropin—a group of hypothyroid patients after
Results:
The percentage of peripheral blood plasmacytoid and myeloid DCs was higher after L-T4 administration when compared with the pretreatment group. Moreover, the expression of CD86 on both DC subtypes was higher in the L-T4 treated than in the hypothyroid patients. In the in vitro experiments, T3 stimulation increased CD86 expression on cultured DCs. The phenotypic difference was paralleled by enhanced ability of T3-stimulated DCs to activate interleukin-12 secretion and proliferation of autologous peripheral blood mononuclear cells (PBMLs) in coculture experiments.
Conclusions:
In the present study, we provide for the first time an evidence that the thyrometabolic status has an influence on the phenotype and function of human peripheral blood DCs. This observation may be of potential importance for the understanding of the pathogenesis of immune and endocrine disorders.
Introduction
Dendritic cells (DCs) are considered as the most potent antigen-presenting cells (1,2). In the immature state, DCs employing a wide range of pattern recognition receptors are the important part of innate immunity against various pathogens (1,3). Later on, because of the extraordinary high ability to capture, process, and present antigens, DCs initiate and direct adoptive immunity, forming the most effective link between these two major types of host defense. Apart from their role in immune system activation, DCs are the main regulators of tolerance induction and maintenance. Thus, DCs may play a role in protective immunity, but also in the induction or perpetuation of autoimmune diseases such as Hashimoto's or Graves' diseases.
Under normal conditions, DCs compose a very rare immune cell population localizing mainly to lymph organs, skin, and peripheral blood. Human peripheral blood DCs consist of two main subsets: myeloid DCs (mDCs) and plasmacytoid DCs (pDCs) (4,5). mDCs and pDCs differ functionally in many aspects, including antigen uptake, cytokine secretion profile, and priming of the immune response and tolerance (5). Moreover, it has been shown that the state of DC maturation (6,7) and, in particular, the expression of costimulatory molecules strongly influence the function of all DC subtypes (8,9). In adoptive immunity, DCs are critically involved in the T-cell differentiation and activation and determine the outcome of the primed immune response. Successful interaction between those cells requires not only effective antigen presentation but also proper costimulation signals delivered by DCs to T cells. The best-characterized costimulatory molecules belong to B7 and TNF/TNFR families. CD80 (B7.1), CD86 (B7.2), and CD40, expressed on antigen-presenting cells, are considered to play a crucial role in the costimulation process (10,11). However, the effect of DC–T-cell interaction depends also on the microenvironmental factors at the immune-reaction site, including numerous cytokines and hormones. Newly published results of animal experiments indicate a profound, direct influence of thyroid hormones (THs) on the maturation and function of bone marrow-derived DCs (12,13). However, the evidence for the role of THs in the biology of human DCs is still lacking.
The aim of the present study was to determine the influence of
Materials and Methods
Patients and controls
Venous blood samples, collected from thyroidectomized patients with differentiated thyroid carcinoma, were submitted to analysis. The patients were recruited from the Department of Endocrinology and Metabolic Diseases, Medical University of Lodz. All of them signed their informed consent, and the Ethics Committee of the Medical University of Lodz had approved the study protocol. Thyroidectomies were performed 1–3 years before the study. Thyroidectomized patients were withdrawn from their L-T4 therapy for at least 5 weeks. Blood samples were collected for plasma analysis in tubes containing heparin (final concentration of 10 IU/mL), between 8 and 9 AM, after an overnight fast. Venous blood was obtained by clean venipuncture (19-gauge needle), avoiding slow flowing draws and/or traumatic venipunctures. The samples were collected at two time points: (i) at the time of the so-called stimulation with endogenous thyrotropin (TSH), that is, when the patients were in the hypothyroid state (n = 21) and were subjected to whole-body scintigraphy, and (ii) during 2 months of L-T4 administration (3–4.5 μg/kg/day) to suppress serum TSH concentrations. The patients with either signs of metastases on whole-body scintigraphy or with immunological or metabolic disorders (i.e., diabetes mellitus) were excluded from the study. The control subjects (n = 20) were free from metabolic disorders and were age-, sex-, and body mass index-matched with the patients. They were recruited from the university staff and their relatives. Subjects with thyroid disorders, goiter, or positive antithyroglobulin antibody (TgAb) or antithyroperoxidase antibody (TPOAb) concentrations were excluded.
Free triiodothyronine (FT3), free thyroxine (FT4), and TSH concentrations were measured by the immunoradiometric method with appropriate kits (BRAHMS; normal values: TSH 0.3–4.0 mIU/L; FT3, 3.4–7.65 pmol/L; FT4, 10–25 pmol/L). The concentrations of TgAb and TPOAb were measured by the electrochemiluminescence method with appropriate kits (Roche Diagnostic; normal values: TgAb <115 IU/mL; TPOAb <35 IU/mL) and equipment (Modular Analytics E170-Roche Diagnostic).
Fluorescence-activated cell-sorting analysis
Peripheral blood mononuclear cells (PBMCs) were isolated from blood samples (obtained as described above) by centrifugation on a discontinuous density gradient (Histopaque 1077; Sigma-Aldrich). The mononuclear cell fraction was washed three times in phosphate-buffered saline, counted, and suspended in phosphate-buffered saline for flow cytometry analysis. PBMCs from patients and healthy controls were assessed by three- or two-color flow cytometry, using a FACSCalibur® cytometer and CELLQuest® software (BD Biosciences). Peripheral blood DCs were recognized by surface expression of the panel of blood DC antigens (BDCAs). The myeloid subset of DCs is defined as a population positive for anti-BDCA1 (AD5-8E7, mouse IgG2a) and negative for anti-CD19 (HIB19, mouse IgG1). Because of a very low number of BDCA3+ cells, mDCs were not further divided for the subtype analysis in the study. The expression of BDCA2 and BDCA4 is specific for peripheral blood pDCs. pDCs were recognized as BDCA2-positive cells by staining with fluorescein isothiocyanate-conjugated antibodies specific for BDCA2 (AC144, mouse IgG1). All DC populations were counterstained with phycoerythrin-conjugated monoclonal antibodies (mAb): anti-BDCA4 (AD5-17F6, mouse IgG1), anti-HLA-DR (G46-6(L243), mouse IgG2a), anti-CD11c (B-ly6, mouse IgG1), anti-CD40 (5C3, mouse IgG1), anti-CD80 (L307.4, mouse IgG1), anti-CD83 (HB15e, mouse IgG1), anti-CD86 (2331(FUN-1), mouse IgG1), anti-CD123 (9F5, mouse IgG1). mAb specific for BDCA antigens were purchased from Miltenyi Biotec. All the other mAb and appropriate isotype controls were purchased from BD Biosciences Pharmingen. A representative flow cytometry analysis of blood DC subpopulations is shown in Figure 1.
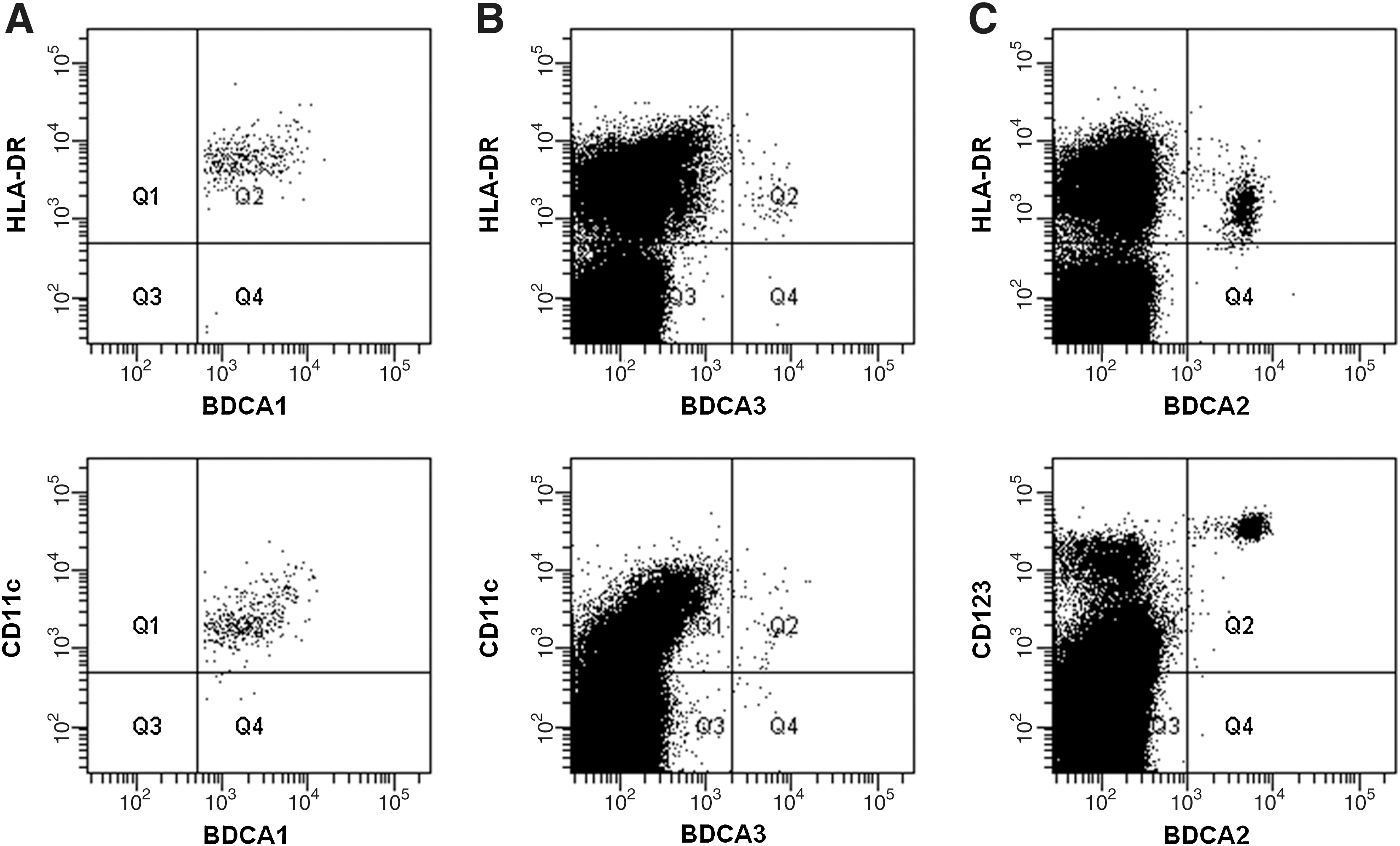
Representative flow cytometry analysis of blood DC subpopulations. Multicolor staining for myeloid DC (mDC) subtypes and plasmacytoid DCs (pDCs) is shown.
Isolation of pDCs
pDCs were isolated from the buffy coats obtained from healthy individuals in Blood Donation Center, Lodz, Poland. All the donors underwent medical examination and were routinely tested for the major infection parameters. PBMCs were isolated by Histopaque 1077 gradient centrifugation (30 minutes, 300 g, 20°C) (Sigma-Aldrich). A total of 2 × 109 cells were then sorted with a BDCA4 isolation kit (Miltenyi Biotec) in the magnetic field of a MidiMACS® sorter (Miltenyi Biotec). Positive fractions routinely contained >90% of BDCA2+CD123+ pDCs. The content of DCs in DC-depleted fraction was routinely <0.1%.
Cell culture
pDCs, isolated as described above, were cultured at a concentration of 3 × 105 cells/mL on a 48-well culture plate for 24 hours in a culture medium (RPMI 1640, streptomycin 100 μg/mL, penicillin 100 U/mL, 2 mM
In coculture experiments, DC-depleted PBMCs, obtained from healthy subjects, were stimulated on round-bottomed 96-well plates (2 × 105 cells/well) with concanavalin A (ConA; Sigma-Aldrich), in the presence or absence of T3 (100 nM) and/or soluble CD40 ligand (sCD40L; 0.5 μg/mL; R&D Systems). In parallel experiments, DC-depleted PBMCs were resupplemented with isolated autologous DCs (ratio of DCs to DC-depleted PBMCs: 1:100, 1:20, and 1:10). All the experiments were done in triplicates. After 48-hour culture, the supernatants were collected for cytokine measurements and 1 μCi of [3H]-thymidine (Amersham) was added to each well for 16 hours. At the end of the culture, the cellular incorporation of [3H]-thymidine was determined. The outcome of coculture proliferation assay was counted as a stimulation index.
Cytokine secretion
Cell culture supernatants were collected, aliquoted, and stored at −20°C. Immediately before the measurement, the aliquots were brought to room temperature and analyzed for the cytokine content (interferon [IFN]-alpha, interleukin [IL]-12). The concentration of cytokines was analyzed by Sandwich ELISA kits (human IFN-alpha ELISA kit [PBL Interferon Source]; human IL-12 Quantikine High Sensitivity Colorimetric Sandwich ELISA kit [R&D Systems]) according to manufacturers' protocols, using the Microplate Reader Model 450 (Bio-Rad).
Statistical analysis
Statistical analyses were performed, using the STATGRAPHICS PLUS v. 5.0 software. Student's t-test for paired samples was used to determine the significance of differences in all the measured parameters with normal distribution, observed between patients before and during L-T4 therapy. Student's t-test for unpaired samples was used to determine the significance of the differences in all the measured parameters with normal distribution, between hypothyroid patients and control subjects, as well as between the L-T4–treated patients and the controls. For the data that were not normally distributed, nonparametric Wilcoxon's rank test (for paired samples) and Mann–Whitney's test (for unpaired samples) were used to determine the statistical significance of differences.
Results
Patients' characteristics
Laboratory data for the patients before and after the treatment are shown in Table 1. Before the treatment, the enrolled patients had higher initial serum TSH levels than age-, sex-, and body mass index-matched healthy controls. All the patients had evident TH deficiency, with a few of them showing very high TSH levels. At the same time, serum FT3 and FT4 levels were significantly and markedly lower in hypothyroid patients than those in the control subjects. During 2 months of L-T4 therapy, as expected, FT3 and FT4 concentrations significantly increased, whereas TSH levels decreased. Serum TgAb and TPOAb concentrations did not differ significantly between the groups, either before or during L-T4 treatment, and were higher in the controls than in the patients groups (Table 1).
Data are presented as means ± standard deviation.
p > 0.05 versus before L-T4 treatment.
p > 0.05 versus control.
DTC, differentiated thyroid carcinoma; BMI, body mass index; FT3, free triiodothyronine; FT4, free thyroxine; TSH, thyrotropin; TgAb, thyroglobulin antibody; TPOAb, thyroperoxidase antibody; L-T4,
The percentage and phenotype of blood DC subtypes are influenced by L-T4 administration
The administration of L-T4 significantly increased the percentage of pDCs (0.198% ± 0.133% vs. 0.298% ± 0.19%; p < 0.05) and mDCs (0.155% ± 0.136% vs. 0.307% ± 0.196%; p < 0.05) in peripheral blood of the patients (Fig. 2). To assess the maturation state of DC subsets in the blood of investigated patients and healthy controls, phenotypic analyses were performed, evaluating the expression of several molecules involved in antigen presentation and costimulation. CD40, CD80, and CD83 were all expressed at negligible levels and L-T4 administration had no influence on their surface expression on DCs. In contrast, L-T4 treatment increased the level of HLA-DR expression on pDCs (57 ± 27.9 mean fluorescence intensity [MFI] vs. 114 ± 38 MFI; p < 0.05) and mDCs (89 ± 9.5 MFI vs. 60.5 ± 4.12 MFI; p < 0.05). As shown in Figure 2, also the percentage of CD86-expressing pDCs and mDCs increased during L-T4 administration (0.079% ± 0.04% vs. 0.126% ± 0.07% [p < 0.05], respectively, for CD86+ pDCs, and 0.14% ± 0.12% vs. 0.28% ± 0.16% [p < 0.05], respectively, for CD86+ mDCs).
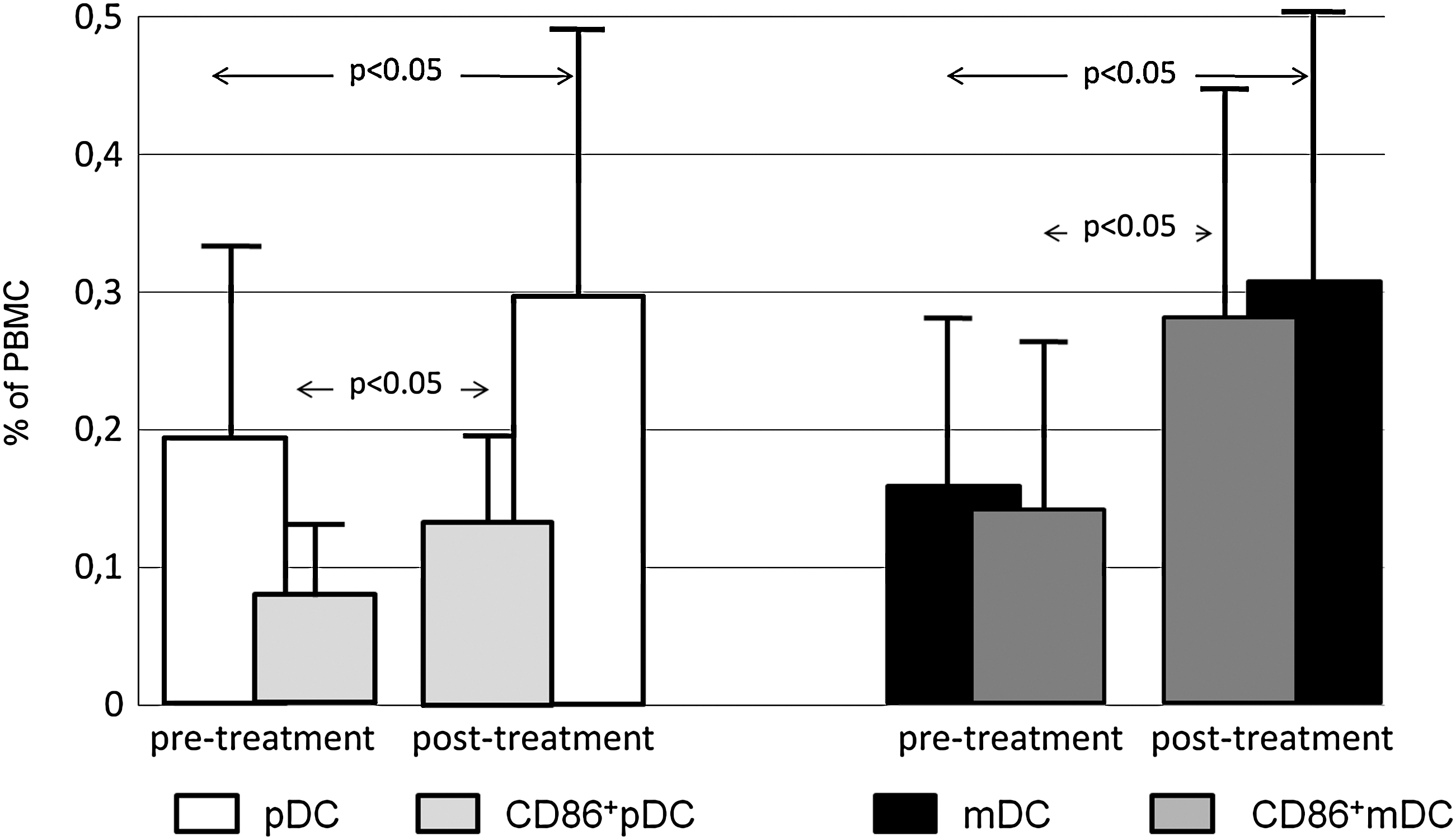
Distribution of human DC subtypes and CD86 expression on DC subtypes (pDC CD86+ and mDC CD86+) in the peripheral blood of patients, monitored after thyroidectomy because of differentiated thyroid carcinoma, before (n = 21, pretreatment group) and during
T3 influences pDCs maturation in vitro
Investigation of direct effects of THs on DCs in humans is extremely difficult, because of very low numbers of naturally occurring peripheral blood DCs—the only fraction of human DCs possible to obtain ex vivo with relatively low invasive measures (venipuncture). Taking into consideration the available methods of DC purification and expected cell yield, human peripheral blood pDC population was isolated from buffy coats from healthy donors for further in vitro experiments.
The sorted pDCs (purity repetitively >90%) were cultured with T3 or with the well-known, very potent pDC stimulator—CpG-ODN—as a positive maturation control. The addition of T3 into the pDC culture significantly increased the percentage of CD86-expressing cells, when compared with the pDCs cultured without T3, thus confirming the effect of THs on peripheral blood DCs ex vivo (Fig. 3). As expected, the addition of CpG-ODN resulted in an even more pronounced increase of the percentage of CD86-positive pDCs (Fig. 3).
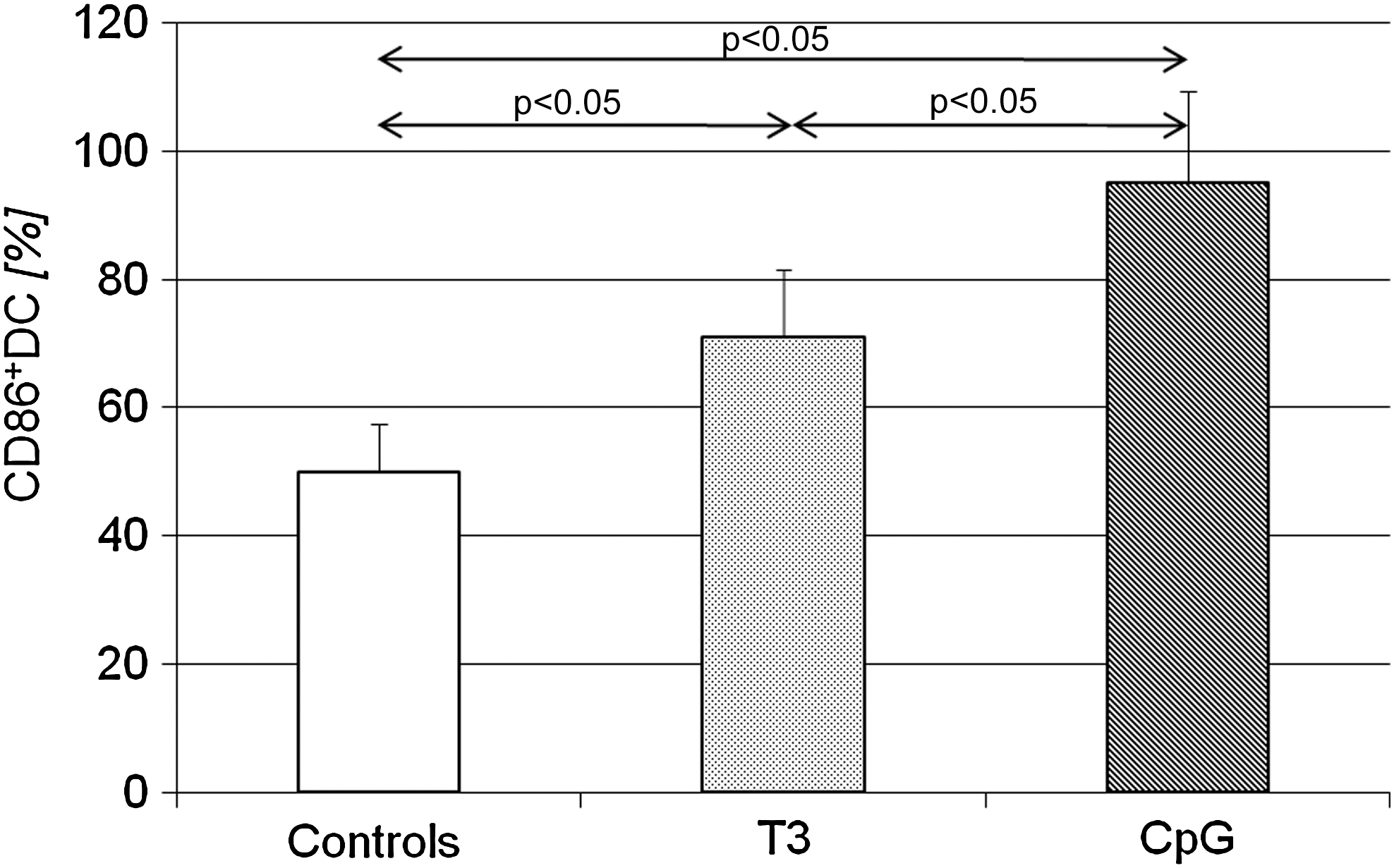
Percentage of pDCs expressing CD86 after 24 hours of culture with T3 or CpG. Data are presented as means ± SD. T3, triiodothyronine.
T3 modulates the DC regulatory function
To assess the influence of T3 on DC regulatory function, we investigated the ability of DCs to modulate the proliferation and cytokine secretion of ConA-stimulated autologous PBMCs in the presence of T3 and/or sCD40L as a DC maturation factor. For that purpose, we resupplemented, in parallel experiments, DC-depleted PBMCs with increasing numbers of freshly isolated autologous DCs. The presence of DCs did not influence either the proliferative or the secretive activity of ConA-stimulated PBMCs. However, in experiments with DCs in the concentration close to that in natural conditions (1% of cultured cells), the administration of T3 and CD40L together showed a statistically significant increase in both the proliferation activity (Fig. 4) and IL-12 secretion (Fig. 5) by ConA-stimulated PBMCs. T3 and CD40L, when administered separately, showed lower stimulatory activity, thus suggesting an additive nature of the observed effect.
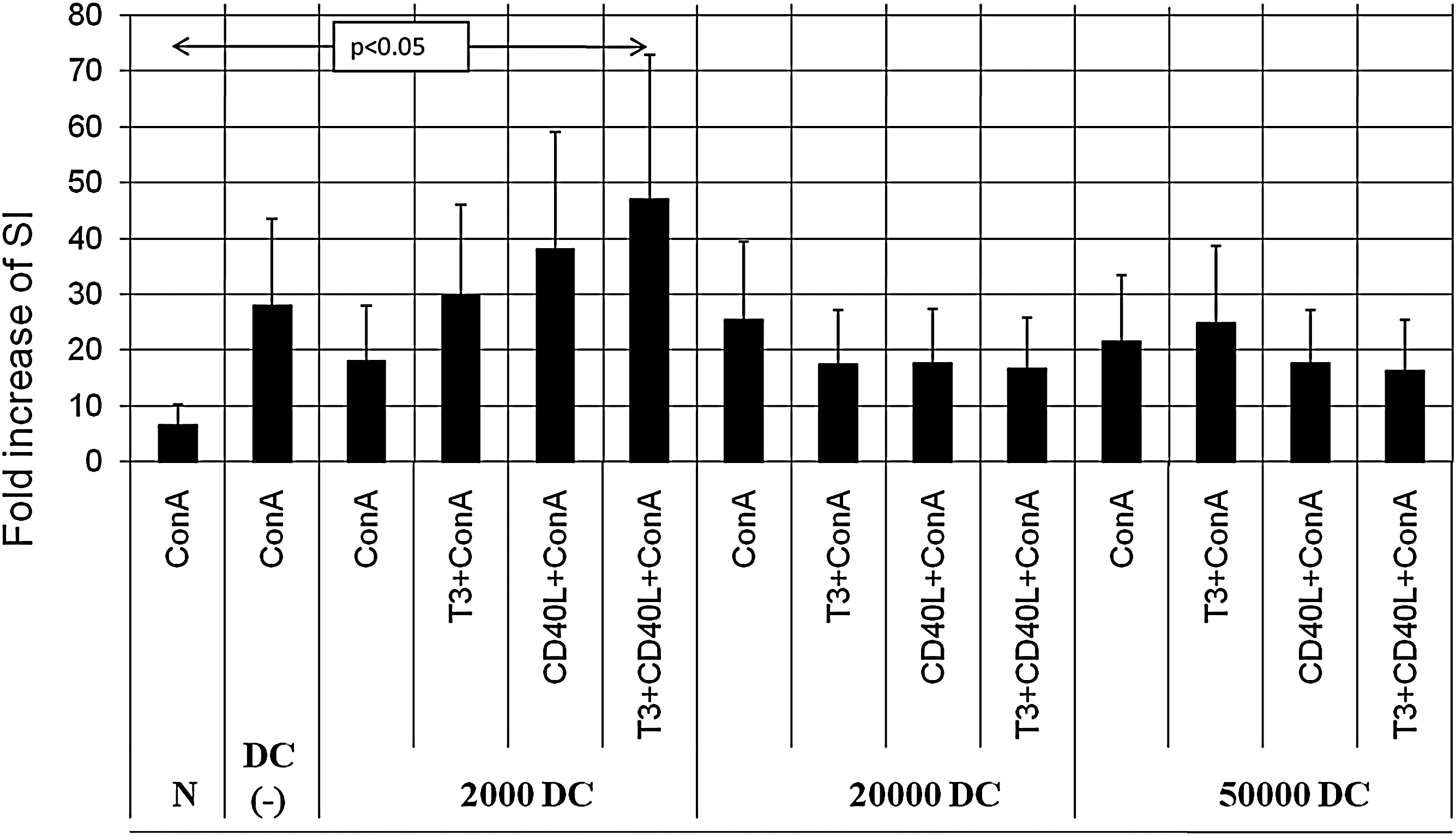
Proliferation of PBMCs stimulated with ConA. PBMCs depleted of DCs were supplemented with increasing numbers of autologous DCs (2000, 20,000, or 50,000 DCs added to DC-depleted PBMCs) and cultured with sCD40L and/or T3. Data are presented as fold increase of stimulation index (SI) ± SD. ConA, concanavalin A.
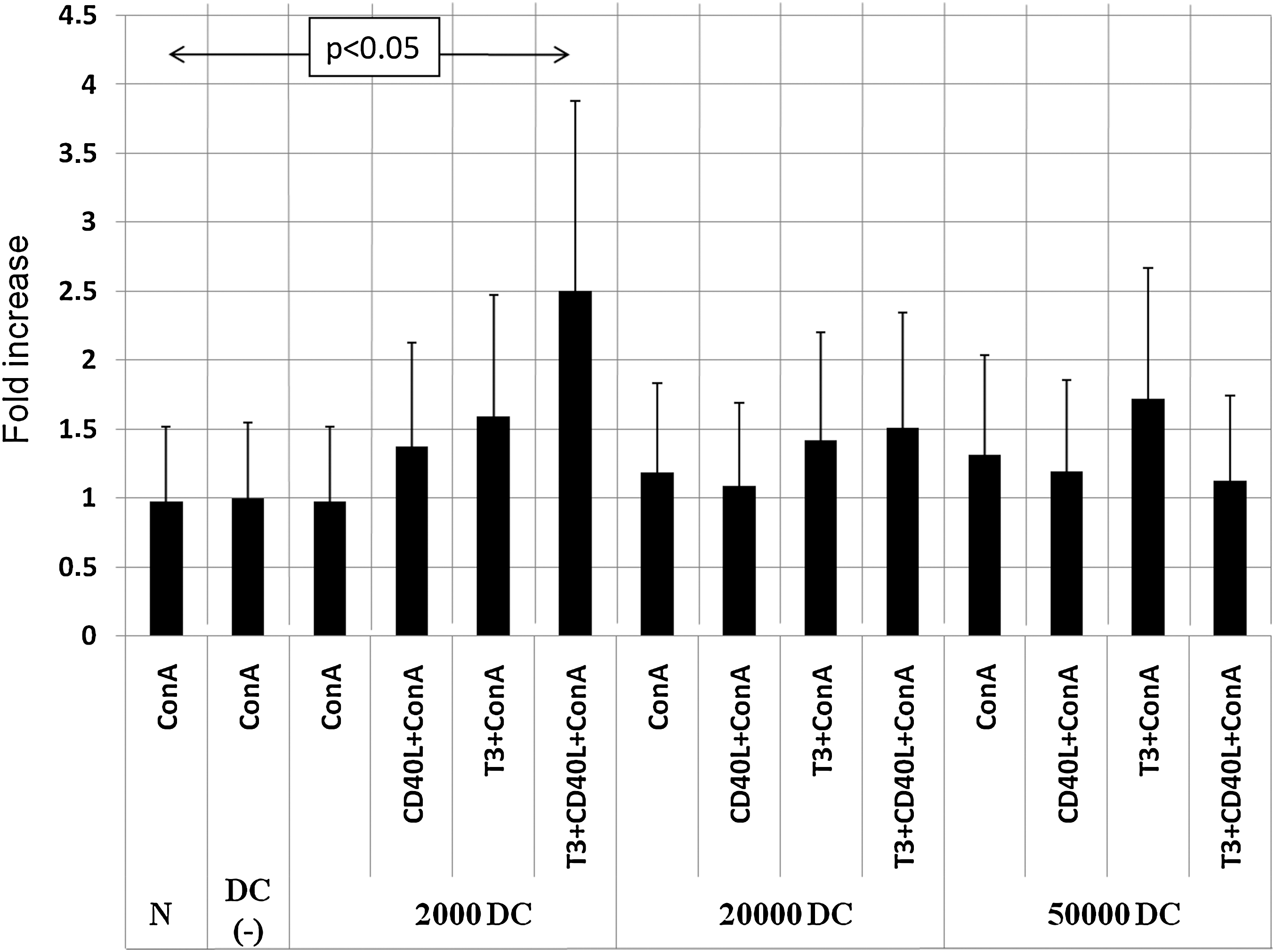
Interleukin-12 (IL-12) secretion by PBMCs stimulated with ConA. PBMCs depleted of DCs were supplemented with increasing numbers of autologous DCs (2000, 20,000, or 50,000 DCs added to DC-depleted PBMCs) and cultured with sCD40L and/or T3. Data are presented as fold increase of IL-12 concentration ± SD, with ConA group normalized as 1.
In those experimental settings, we observed a low IFN-alpha production, and there was no significant difference in secretion of this cytokine in the analyzed groups (data not shown).
Discussion
More and move evidence indicates an involvement of the endocrine system in the regulation of immunity, including DCs. As highly specialized antigen-presenting cells, DCs recognize, process, and present antigens to T cells and control the induction, maintenance, and direction of antigen-specific immune responses. Different hormones influence the maturation and activity of DCs. Progesterone inhibits TLR-9–induced IFN-alpha production by human and mouse pDCs (14) and is believed to have a suppressing influence on DCs. Estrogens seem to exert an activating effect on DCs in lupus erythematosus (15). On the other hand, tamoxifen and toremifen suppress the maturation and some functions of DCs (16). In culture, bone marrow progenitors give rise to the generation of DCs (17), which can be influenced by the action of androgens and estrogens (18,19). In addition, mRNAs for estrogen receptors have been demonstrated in CD14 monocytes, cultured immature CD1a cells, and mature CD83 cells (18). Moreover, glucocorticoids inhibit the in vitro differentiation of DCs from their progenitors and impair their capacity to undergo terminal differentiation or generate proinflammatory cytokines (20). The active form of vitamin D also exerts immunosuppressive effects, and numerous studies have shown that the regulation of DC function is an important part of this suppressive activity. 1α,25 Dihydroxy vitamin D3 suppresses profoundly the maturation and activation of DCs, consequently leading to decreased T cells (21,22).
Similar to THs, all the aforementioned hormones exert their action via nuclear receptors and belong to the nuclear hormone receptor superfamily. Although it is becoming increasingly evident that members of that group are involved in the regulation of DC biology (23), little is known about the influence of THs on DCs. Mascanfroni et al. investigated the influence of T3 on murine bone marrow-derived DCs in culture. The authors confirmed the presence of TH receptor in bone marrow-derived DCs and showed the stimulatory effect of T3 on DC maturation and function (12). In the newly published study, the same group described the intracellular mechanisms underlying the observed effect of T3 on murine DC. T3- and TH receptor-dependent activation of NF-κB and Akt transcriptional pathways in DCs implies a direct involvement of THs in the regulation of immune response (13).
In our study, we used a clinical model in which DCs were investigated in the same patients in two different thyrometabolic states—short-term hypothyroidism and short-term subclinical thyrotoxicosis. The patients participating in the study were free from other metabolic or immunologic diseases. So, they presented an almost perfect model to analyze the influence of L-T4 administration on DCs. The main finding of our study was the influence of thyrometabolic state on naturally occurring human peripheral blood DCs. In our longitudinal ex vivo measurements, L-T4 substitution in hypothyroid patients increased the percentage of both major peripheral blood DC subtypes: pDCs and mDCs. Moreover, under L-T4 treatment, an increased percentage of CD86-expressing pDCs and mDCs was observed and, additionally, an increased level of HLA-DR expression was found on both DC subsets. HLA-DR and CD86 play a crucial role in the two very important steps of adoptive immune response—antigen presentation and costimulation, respectively (24,25). The positive effect of thyrometabolic state normalization on the aforementioned parameters implies a modulatory role of THs in the immune balance control.
The relationships between L-T4 administration and changes in DC quantity and phenotype in our model are suggestive of a specific L-T4 effect. However, L-T4 treatment deprival and readministration caused direct and indirect changes in the thyrometabolic hormone profile. Therefore, the changes in DCs may have theoretically resulted from the simultaneous action of TSH and THs. TSH receptors are present in DCs and induce a stimulatory effect on phagocytosis and cytokine production in murine DCs (26).
To assess the direct influence of T3 on DCs, we performed a set of in vitro experiments with freshly isolated peripheral blood pDCs. PDCs, cultured in the presence of T3, showed an increased expression of CD86, thus confirming the observation made in ex vivo analysis in L-T4–treated patients. Moreover, in our coculture experiments, T3 in combination with the DC maturation factor sCD40L increased the ability of DCs to upregulate the proliferative response and secretion of IL-12 by PBMCs under nonspecific stimulation with ConA. This remains in agreement with the results of animal studies by Mascanfroni et al. (12,13). Similar to our findings in humans, exposure of murine DCs to T3 resulted in an increased expression of costimulatory molecules including CD86 as well as higher level of IL-12 secretion. The well-known proinflammatory characteristic of IL-12 and its putative involvement in Th1-mediated (27 –29) organ-specific autoimmune diseases (30) imply potential clinical relevance of our observation, especially with respect to previous studies reporting an increased synthesis of IL-12 by DCs obtained from hyperthyroid mice (31) as well as increased amounts of IL-12 in sera from patients with Graves' disease (32).
In the present study, for the first time, we present evidence that thyrometabolic status influences human DC phenotype and function. The pathological relevance of functional DC abnormalities has been suggested in various human disorders (33 –35), including the diseases without classical immunological background (36). It has been also shown that DCs can be considered as a specific therapeutical target (33,36). In the light of the aforementioned observations, our findings seem to have a potential meaning for the understanding of the mechanisms underlying TH-associated pathological processes as well as in designing of new therapeutic attempts.
Footnotes
Acknowledgments
This work was supported by grants from the Polish Ministry of Science and Higher Education (No. 2P05B14826) and the Medical University of Lodz (No. 50211422). The authors express great appreciation to the patients and healthy volunteers for their generous decision to participate in this study.
Disclosure Statement
The authors declare that no competing financial interests exist.