
Abstract
Background:
More than 80% of all thyroid cancers, the most common endocrine malignancy, are papillary thyroid cancer (PTC). It is well established that CITED1 (Cbp/p300 Interacting Transactivators with glutamic acid [E] and aspartic acid [D]-rich C-terminal domain) mRNA is characteristically overexpressed in PTC. Our previous study suggested a positive association of BRAF mutation with CITED1 overexpression. However, the mechanism of CITED1 expression in PTC remains to be elucidated. In the present study, we analyzed whether aberrant methylation of CITED1 gene promotes CITED1 overexpression in PTC.
Method:
CITED1 mRNA expression levels were analyzed by quantitative polymerase chain reaction in three PTC-derived cell lines, TPC1, K1, and KTC-1, and in surgically dissected PTC and surrounding normal tissues from 19 patients. The BRAF mutation status of the cells and clinical specimens was determined by direct sequencing. The methylation status of the deoxycitidine-phosphate-deoxyguanosine dinucleotides (CpGs) in the CITED1 promoter was analyzed by the bisulfite-sequencing method using genomic DNA. Finally, the expression of CITED1 mRNA in TPC1 cells, when subjected to pharmacological inhibition of methylation, was analyzed.
Results:
CITED1 mRNA was expressed at lower levels in TPC1 than in K1 and KTC-1 cells. A BRAF mutation was present in K1 and KTC-1 cells, but not in TPC1 cells. CITED1 promoter was hypomethylated in K-1 and KTC-1 cells, but not in TPC1 cells. In surgically dissected specimens, the mean expression level of CITED1 mRNA was 30-fold higher in PTC tissues than in normal tissues. CpGs in the CITED1 promoter were more heavily methylated in normal tissues than in PTC tissues. In PTC specimens without a BRAF mutation, two CpGs were more heavily methylated than in PTC specimens with the BRAF V600E mutation. Pharmacological inhibition of methylation in TPC1 cells by 5′-aza-2′-deoxycitidine resulted in increased expression of CITED1 mRNA.
Conclusion:
Hypomethylation of the CpGs in the promoter region of CITED1 is associated with higher expression of CITED1 mRNA in PTC tissues, consistent with the hypothesis that epigenetic regulation is involved in the overexpression of CITED1. This hypothesis is supported by pharmacologic inhibition studies in TPC1 cells.
Introduction
Genetic alterations are responsible for thyroid tumorigenesis and progression. BRAF is one of three Raf kinases (8,9), and among the RET–Ras–Raf–MEK–MAPK/ERK signaling pathway, it is the most potent activator of the mitogen-activated protein kinase (MAPK) pathway, which contributes to various cellular events such as cell proliferation, apoptosis, survival, differentiation, and tumorigenesis (9 –12). BRAF gene mutations have been identified in various cancers (13), and point mutations in the BRAF gene are the most prevalent genetic alteration in PTC (14 –18). The most common hot-spot mutation of the BRAF gene site is a thymine-to-adenine transversion at nucleotide 1799 (T1799A) in exon 15, resulting in valine-to-glutamate substitution at residue 600 (V600E) in thyroid carcinoma, with an average prevalence ranging from 28% to 83% (7,19 –22).
It is known that a number of genes are characteristically overexpressed in PTC, and these genes have served as possible markers for the molecular diagnosis of this cancer (20,23 –25). However, the mechanisms involved in the overexpression of these genes are not fully understood. Cbp/p300 Interacting Transactivators with glutamic acid [E] and aspartic acid [D]-rich C-terminal domain (CITED1) is one such gene (26,27), and our previous study suggested a possible link between BRAF mutation and CITED1 overexpression; however, silencing of BRAF expression by siRNA in a cell line expressing BRAF V600E did not result in suppression of CITED1 expression (23,28), leaving the mechanism of CITED1 overexpression to be elucidated.
Recently, the epigenetic modification of the genome has emerged as a mechanism of cancer-associated alteration in gene expression (29). Methylation of deoxycitidine-phosphate-deoxyguanosine dinucleotide (CpG) islands is involved in various biological and pathological processes and is physiologically involved in genomic imprinting, X-chromosome inactivation, and embryonic development (16). Aberrant methylation of CpG islands in the promoter regions of tumor suppressor genes is known to cause silencing of the gene and can work as one of the mechanisms to inactivate it, along with gene mutation and loss of heterozygosity (16,29 –31). In addition to inactivation of a gene by methylation being reported for many tumor suppressor genes, evidence for activation of specific genes, including oncogenes, by demethylation in cancer has also been reported (32 –34). In the present study, we analyzed the methylation status of CITED1 promoter in PTC cell lines and clinical specimens. We explored the possibility that a lower methylation ratio of the CpGs in the promoter region of CITED1 is associated with higher expression of CITED1 mRNA.
Materials and Methods
Clinical samples
The thyroid tissues used in this study were obtained between 2007 and 2008 from 19 patients who had undergone total thyroidectomy at the Department of Breast and Endocrine Surgery, Nagoya University Hospital. The patients ranged in age from 25 to 77 years. Thirteen were women and six were men. Approximately 30 mg of PTC and surrounding normal thyroid tissues were dissected immediately from the material removed by surgery. The samples were snap-frozen in liquid nitrogen and stored at −70°C. Informed consent for the use of the tissues for clinical research was obtained before surgery and the study protocol was approved by the Institution Review Board of Nagoya University Hospital and Medical School in accordance with the guidelines in the Declaration of Helsinki.
Cultured cells
Three cell lines derived from PTC were analyzed in the present study. TPC1 (35) was provided from Dr. Norisato Mitsutake (Nagasaki University) and K1 (36) was purchased from the European Collection of Cell Cultures; KTC-1 has been described previously (37). TPC1 and KTC-1 cells were cultured in RPMI 1640 medium containing 10% fetal bovine serum at 37°C under a humidified atmosphere of 5% CO2 and 95% air. K1 cells were cultured in Dulbecco's modified Eagle's medium/Ham's F-12 with 10% fetal bovine serum under the same conditions.
Genomic DNA extraction
Genomic DNA from frozen PTC samples and cell lines was isolated with SDS-proteinase K digestion, phenol–chloroform extraction, and ethanol precipitation as previously described (23).
BRAF mutation analysis by direct sequencing
A 224-bp sequence including the T1796A locus in exon 15 of the BRAF gene (NT_007914) was amplified using specific primers (forward primer: 5′-TCATAATGCTTGCTCTGATAGGA-3′; reverse primer: 5′-GGCCAAAAATTTAATCAGTGGA-3′) and Platinum®PCR SuperMix High Fidelity (Invitrogen, Life Technologies Japan). The sequence of the amplified fragments was analyzed by direct sequencing, using a BigDye terminator v3.1 cycle sequencing kit and ABI 3100-Avant genetic analyzer (Applied Biosystems, Life Technologies Japan).
Sodium bisulfite modification and sequencing
To convert unmethylated cytosine residues into uracil, genomic DNA was modified and purified using an EZ DNA Methylation-Gold Kit (Zymo Research) according to the supplier's protocol. Bisulfite-modified genomic DNA was amplified by nested polymerase chain reaction (PCR) using two sets of primers and a Platinum PCR SuperMix High Fidelity (Invitrogen). The sequence of the primers was designed using Methyl Primer Express Software v.1.0 (Applied Biosystems). The primers used for the first-round amplification were the forward primer 5′-AGTGGAATTTATTGGGTAAGTTT-3′ and the reverse primer 5′-ATACCCCTTCTACAACTTTACC-3′. For the second-round amplification, the forward primer 5′-GTGGAATTTATTGGGTAAGTTTA-3′ and the reverse primer 5′-CCRAACACATTTTTATAACAAC-3′ were used.
To eliminate primers and Platinum PCR SuperMix, PCR products were purified using QIA quick PCR Purification Kit (Qiagen). The purified PCR products were cloned into pGEM-T easy plasmids (Promega Japan) using DNA ligation kit (Takara Bio) according to the manufacturer's manual. At least four independent clones were sequenced for each clinical sample or cell line, and the methylation ratio was calculated. As all the cytosine residues that do not comprise CpG dinucleotide were converted into thymidine residues, the efficiency of bisulfite modification was considered to be 100%.
The methylation status of the promoter was also evaluated by M-quant analysis described by Leakey et al., which allows quantitative evaluation of methylation status with comparable accuracy to the combined bisulfite restriction assay (38). Briefly, the peak of thymidines were measured on electrophoregram of direct sequencing of bisulfite-modified DNA. Methylation of CpG sites results in decrease of the thymidine peaks because of the presence of methylated cytidines that are resistant to bisulfite-induced conversion. The percentage of decrease of the thymidine peak at a specific CpG site relative to the average thymidine peak surrounding the CpG sites represents the methylation ratio (38).
Pharmacological inhibition of DNA methylation in cultured cells
To inhibit DNA methylation in TPC1 cells, the cells were cultured in the presence of 5 or 10 μM of 5′-aza-2′-deoxycytidine. The culture medium containing 5′-aza-2′-deoxycytidine was changed every 24 hours. RNA was extracted for analyses after 96 hours of exposure to 5′-aza-2′-deoxycytidine.
RNA extraction and quantitative reverse transcription–PCR analysis
Total RNA was extracted from the tissue samples and cultured cells using the RNeasy Mini kit (Qiagen) according to the manufacturer's instructions. Complementary DNAs were synthesized using ReverTra Ace qPCR RT Kit (Toyobo Co.), and cDNAs equivalent to 5 ng of total RNA were subjected to real-time monitored quantitative PCR (cDNA-qPCR) as previously described in detail (23). Primers used to amplify CITED1 cDNAs were the forward primer 5′-ATGTCAAGGGTGGCACCTCA-3′ and the reverse primer 5′-GCCTATTGGAGATCCCGAGGA-3′, and those for beta-actin (ACTB) were the forward primer 5′-TTCCATCATGAAGTGTGACGTTGAC-3′ and the reverse primer 5′-GCTGATCCACATCTGCTGGAAG-3′. The expression levels of CITED1 controlled by those of ACTB were presented.
Statistical analysis
Results are expressed as means ± standard error. Statistical analyses were performed using Student's t-test and analysis of variance. p-Values of <0.05 were considered to be significant.
Results
We first analyzed the three PTC-derived cell lines for the mutation status of BRAF, expression levels of CITED1 mRNA, and methylation status of CITED1 promoter. K1 and KTC-1 had a heterozygous BRAF V600E mutation, whereas no mutation was detected in the analyzed region in TPC1 (data not shown). These results are in agreement with the report by Schweppe et al. (28). As shown in Figure 1a, the expression levels of CITED1 mRNA were significantly higher in K1 and KTC-1 cells compared with that of TPC1 cells. These results were in agreement with our previous studies suggesting an association of high CITED1 mRNA expression with BRAF mutation (23).
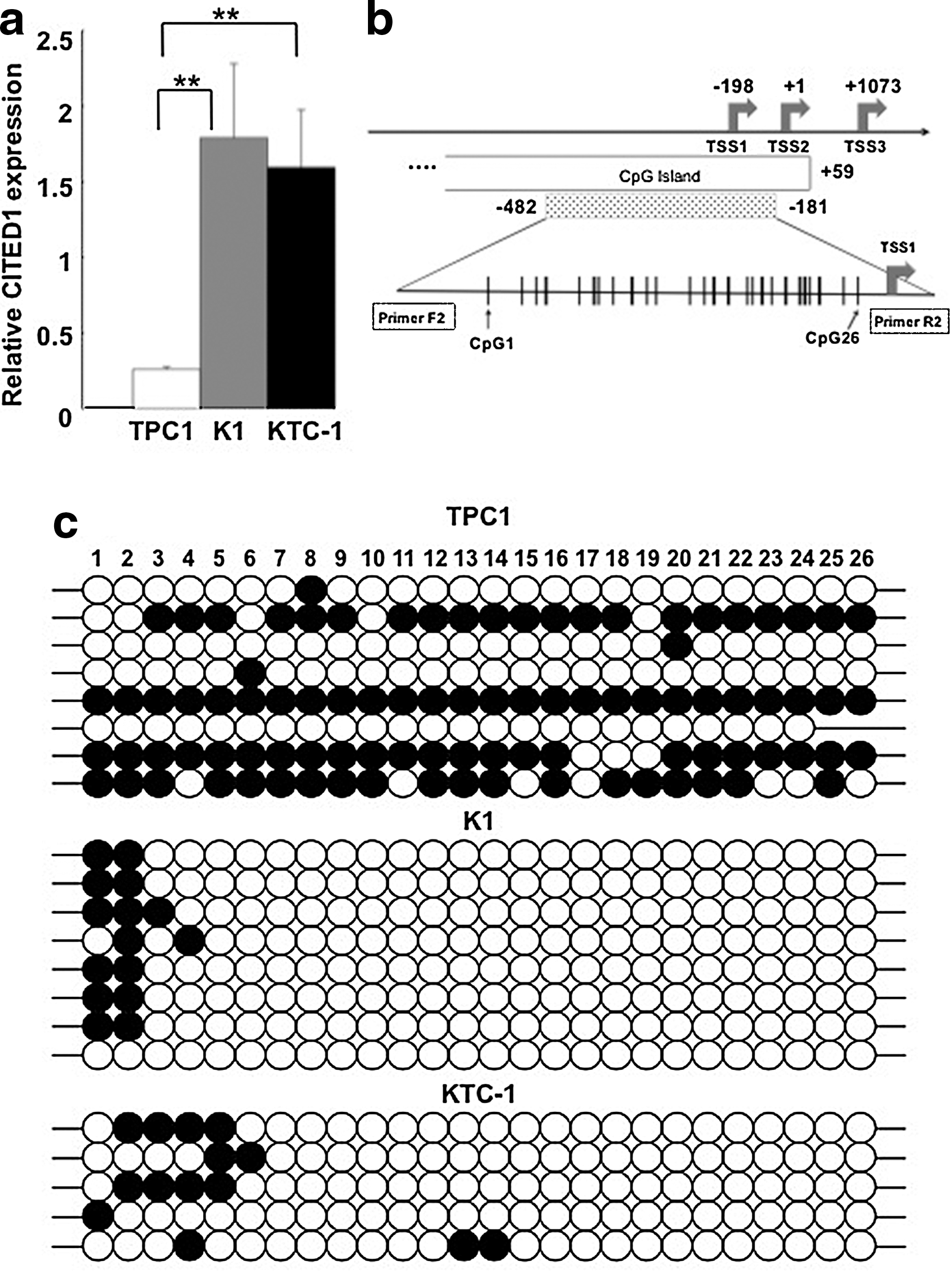
Expression of CITED1 mRNA and methylation status of CITED1 promoter in papillary thyroid cancer (PTC)-derived cell lines.
We then analyzed the methylation status of CITED1 promoter in these three cell lines. Using Methyl Primer express v.1.0., a CpG island prediction program, we identified the CpG island in CITED1 promoter (Fig. 1b); the island included two of the three transcription start sites (TSS1, TSS2, and TSS3) that have been identified in CITED1 gene. We choose a primer that would amplify nucleotide position −482 to −181 bp, relative to the second transcription start sites (TSS2). Twenty-six CpGs were present in the amplified region, and the methylation status was analyzed by bisulfite sequencing. As shown in Figure 1c, almost all the CpGs were methylated in half of the TPC1 clones (4/8), whereas only four CpGs, CpG1 (−459 bp) to CpG4 (−431 bp), were methylated in K1 cells. In KTC-1 cells, six CpGs in the upstream portion, CpG1 (−459 bp) to CpG6 (−401 bp), were heavily methylated. These results supported the hypothesis that low methylation of the analyzed region promotes high expression of CITED1 mRNA.
We then analyzed PTC tissues and the surrounding normal thyroid tissues from the 19 subjects for expression levels of CITED1 mRNA and methylation status of its promoter. The expression levels of CITED1 mRNA in PTC tissues were ∼30-fold higher than in normal tissues (p = 0.000056; Fig. 2a). The methylation status of CITED1 promoter in the 19 PTC tissue samples and the adjacent normal tissues was determined by bisulfite sequencing. On average, four independent clones were sequenced for each sample to calculate the methylation ratio for each sample. As shown in Figure 2b, methylation ratios in normal tissues were slightly higher than those in PTC tissues for most of the CpGs analyzed. Among these, for two CpGs, CpG22 (−251 bp) and CpG25 (−210 bp), the differences in methylation ratio between PTC and normal adjacent tissues was significant (Fig. 2b). The methylation status was also evaluated by M-quant analyses of direct sequencing results and similar results were obtained (Fig. 2c). Collectively, in both cell lines and clinical specimens, a lower methylation ratio of CpGs in CITED1 promoter was associated with higher expression of CITED1 mRNA. These results suggested that the differential methylation status of the CITED1 promoter is involved in the regulation of CITED1 expression in PTC cell lines and tissues.
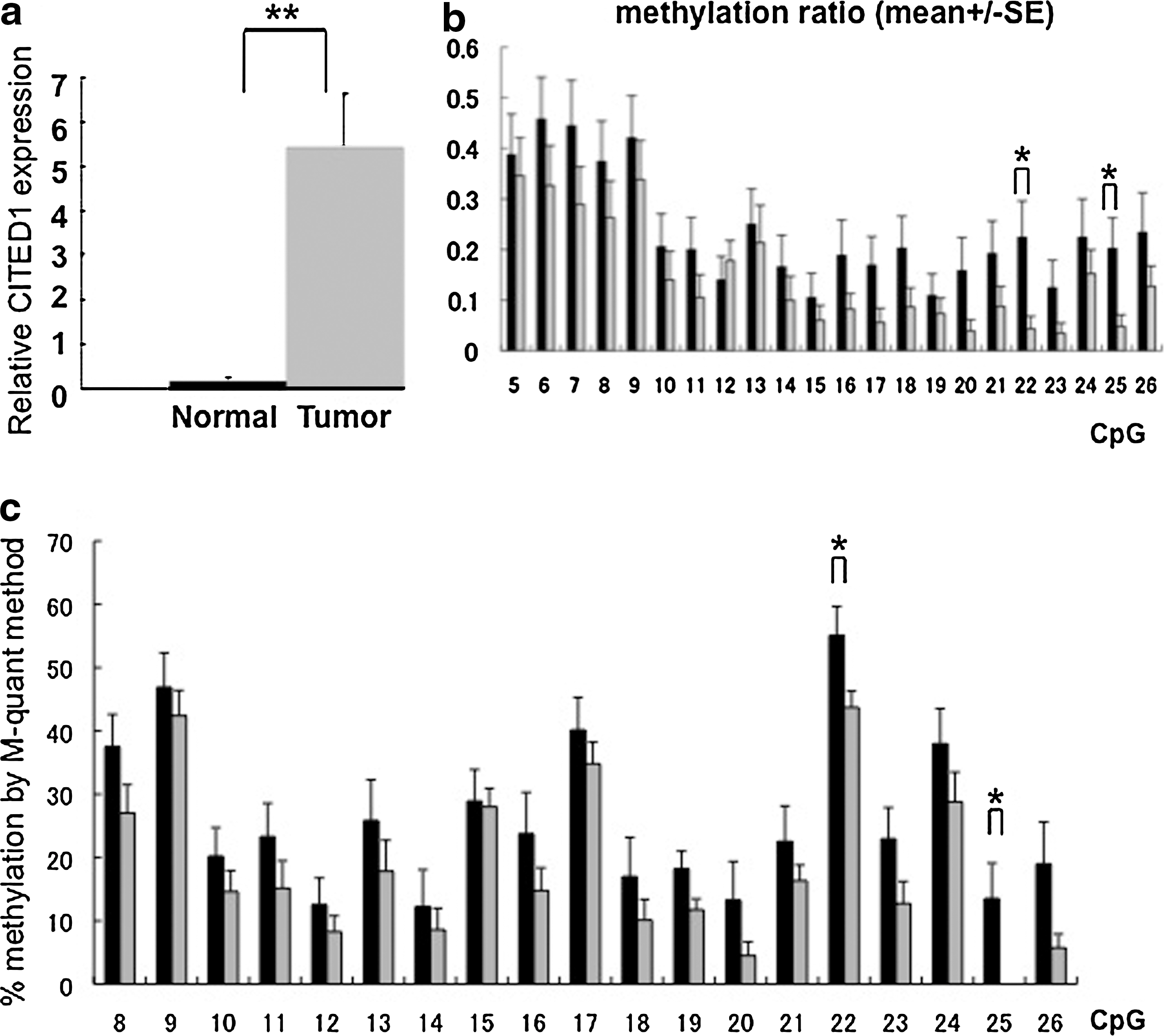
Expression of CITED1 mRNA and methylation status of CITED1 promoter in PTC and surrounding normal thyroid tissue samples from 19 subjects.
As 15 of the 19 PTC specimens carried BRAF V600E mutations (data not shown), we analyzed whether the presence or absence of BRAF mutation affects CITED1 mRNA expression and promoter methylation. Although the difference did not reach statistical significance, the expression of CITED1 mRNA in PTC specimens with BRAF mutation was higher than that in PTC specimens with the wild-type BRAF (Fig. 3a). Methylation ratios were lower in the CpGs close to TSS (CpG12–26) in PTC specimens with BRAF mutation. Especially, CpG18 (−287 bp) and CpG24 (−230 bp) showed lower methylation ratio in PTC specimens with BRAF mutation (Fig. 3b). Although the difference did not reach statistical significance, similar results were obtained by M-quant analyses (Fig. 3c). Therefore, BRAF V600E may affect the methylation status of the CITED1 promoter, and this mechanism may be, at least in part, involved in the overexpression of CITED1 mRNA in PTC.
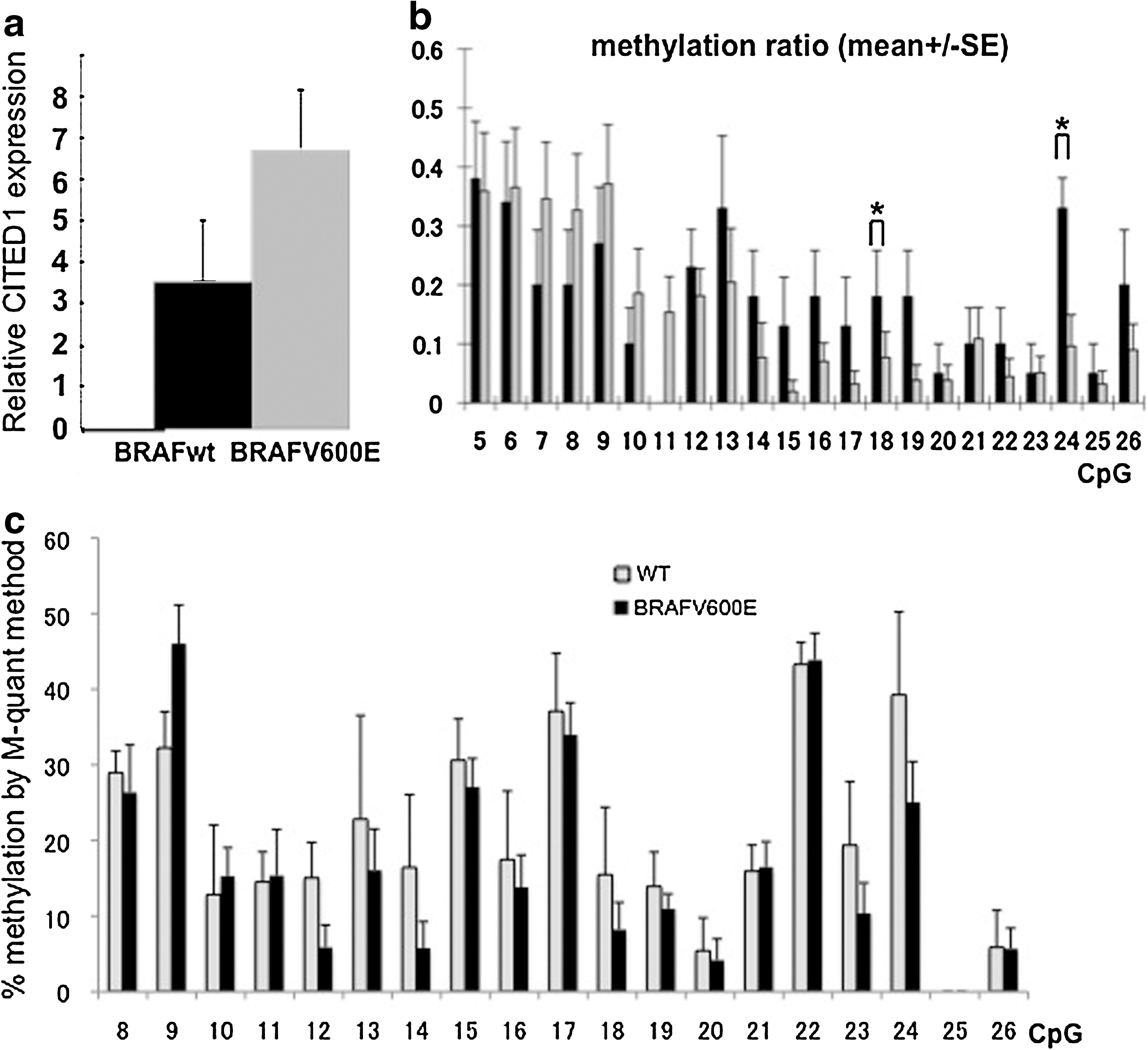
Expression of CITED1 mRNA and methylation status of CITED1 promoter in PTC with or without BRAF V600E mutation.
Finally, we tested whether the pharmacological inhibition of methylation in TPC1 cells alters the expression of CITED1 mRNA. As shown in Figure 4, treatment of TPC1 cells with 5 μM of 5′-aza-2′-deoxycitidine resulted in significant increase in CITED1 mRNA expression. These results further indicated that lower promoter methylation is associated with high expression of CITED1 mRNA. Treatment with 10 μM of 5′-aza-2′-deoxycitidine, however, did not significantly induce CITED1 mRNA expression. It may be that 5′-aza-2′-deoxycitidine at this concentration has a direct repressive effect on CITED1 expression.

Expression of CITED1 mRNA in TPC1 cells treated with 5′-aza-2′-deoxycitidine. TPC1 cells were treated with indicated concentration of 5′-aza-2′-deoxycitidine for 96 hours. Total RNA was extracted from the cells and subjected to quantitative polymerase chain reaction analyses. Expression levels of CITED1 mRNA controlled by those of ACTB mRNA are shown (n = 6, mean ± SE; *p < 0.05).
Discussion
In the present study, we analyzed the methylation status of the CITED1 promoter in PTC cell lines and specimens of PTC tissue. Our results show that a lower methylation ratio of the CpGs in the promoter region of CITED1 is associated with higher expression of CITED1 mRNA and suggest the involvement of epigenetic regulation in the overexpression of CITED1 in PTC. Although aberrant methylation of antioncogenes has been implicated in the pathogenesis of cancer development, the role of aberrant demethylation of promoters in cancer is obscure. Nevertheless, the activation of a specific gene by demethylation in cancer has been reported in several studies (32 –34,39).
CITED1 is characteristically overexpressed in PTC; however, it is not expressed in anaplastic thyroid cancer (27). These findings suggest that the expression of CITED1 is downregulated on anaplastic transformation of PTC. Alternatively, the expression of CITED1 may indicate a reduced risk for anaplastic transformation of PTC. Therefore, the significance of CITED1 as a marker to predict prognosis of the disease remains completely obscure. We have previously reported that CITED1 expression is higher in PTC expressing BRAF V600E than in PTC without BRAF mutation (23). In the present study, our analyses showed that the methylation ratio of the CpGs in CITED1 promoter, especially CpG18 and CpG24, was lower in PTC with BRAF V600E than in PTC without BRAF V600E. As several studies have reported that the BRAF mutation is associated with relatively poor prognosis, overexpression of CITED1 may be also associated with poor prognosis. It should be noted that, however, a recent large-scale Japanese study conducted in 631 patients with PTC did not find that the BRAF V600E mutation was associated with a poor prognosis in their Japanese population (7). Further studies are required to clarify the significance of CITED1 overexpression as a predictor of prognosis.
In nevi and primary melanomas, the aberrant expression of a microtubule-associated protein (MAP2) has been reported. Interestingly, MAP2 is not expressed in metastatic melanomas, and patients whose primary tumors expressed abundant MAP2 showed better disease-free survival rates than patients with weak or no MAP2 expression (34). Importantly, BRAF V600E induces the expression of MAP2 in melanoma cells, and a recent study has demonstrated that promoter demethylation plays an important role in the induction of MAP2 (34). However, promoter demethylation is not the sole mechanism that activates MAP2 expression, and alternative mechanisms, for example, the downregulation of transcription factor, are also involved (34). Such alternative mechanisms are different from one melanoma cell line to another.
It is likely that the altered methylation status of CITED1 promoter is not the only factor that induces overexpression of CITED1 mRNA. We have previously analyzed whether silencing of BRAF expression represses the expression of CITED1 in a cell line carrying BRAFV600E mutation. Unexpectedly, we found that silencing of BRAF was associated with an increase in CITED1 mRNA (23). To fully elucidate the mechanism of CITED1 overexpression, further analyses using multiple PTC-derived cell line are required. This is currently difficult as there is a relative paucity of PTC-derived cell lines compared with melanoma-derived cell lines (28).
In the present study, we have shown for the first time that the methylation status of the CITED1 promoter is altered in PTC tissues. As the difference in the promoter methylation of CITED1 gene in PTC and normal tissues was modest, mechanisms other than promoter demethylation might also be involved in the overexpression of CITED1 in PTC. Further studies should clarify the involvement of epigenetic regulation in the altered gene expression in PTC.
Footnotes
Acknowledgments
The authors are indebted to Dr. Norisato Mitsutake (Nagasaki University) for providing TPC1 cells. This work was supported in part by the Charitable Trust Soyu Medical Foundation (to M.S.).
Disclosure Statement
The authors declare that no competing financial interests exist.