
Abstract
Background:
Anaplastic thyroid carcinoma (ATC) is the most aggressive type of thyroid carcinoma. The purpose of this study was to evaluate the combined cytotoxic effects of paclitaxel and lovastatin in ATC cell lines.
Methods:
ATC cells were treated with paclitaxel and lovastatin, separately or together, and the cytotoxicity of the compounds was determined by quantifying cell viability and apoptosis. We conducted an isobologram analysis to investigate the combined effect of the two drugs.
Results:
In 8505C cells, cellular viability was inhibited by lovastatin and paclitaxel in a concentration-dependent manner (p = 0.002 and p = 0.020, respectively). The IC50 of lovastatin was 3.53 μM and that of paclitaxel was 5.98 nM. In BHT-101 cells, cellular viability was also inhibited in a concentration-dependent manner by lovastatin and paclitaxel (p = 0.020 and p = 0.032, respectively). The IC50 of lovastatin was 17.13 μM and that of paclitaxel was 35.26 nM. In 8505C cells, paclitaxel and lovastatin alone induced apoptosis in a concentration-dependent manner. However, both an isobologram analysis on inhibition of viability and an analysis of apoptosis demonstrated antagonism between paclitaxel and lovastatin. In BHT-101 cells, however, neither drug had an apoptotic effect when used individually. There was a variable effect when used in combination, depending on the drug concentrations.
Conclusions:
Paclitaxel and lovastatin were cytotoxic in two ATC cell lines and increased apoptosis in 8505C cells. However, in these cells, the combination of drugs resulted in antagonism that affected both the cytotoxicity of the compounds and the apoptosis of 8505C cells. The combination of paclitaxel and lovastatin did not enhance the treatment effect in ATC cell lines.
Introduction
Various doxorubicin-based chemotherapeutic agents are used for the treatment of ATC. Paclitaxel is a compound that enhances tubulin polymerization via high-affinity binding to microtubules, thereby inhibiting the normal dynamics of the microtubule network. This results in the inhibition of mitosis and cell division. Paclitaxel is widely used to treat various types of solid tumors including breast, ovarian, and lung cancer (5). Several studies have examined the cytotoxic effects of paclitaxel on ATC cell lines (6,7). In a phase-II clinical trial, paclitaxel has shown some therapeutic effects in the treatment of ATC (8), and Higashiyama et al. have reported that induction chemotherapy with weekly paclitaxel is a promising therapeutic strategy for treating stage patients with IVB ATC (9).
Lovastatin, a 3-hydroxy-3-methylgutaryl CoA (HMG-CoA) reductase inhibitor (commonly referred to as a statin), has demonstrated both preventative and therapeutic effects against cardiovascular diseases. Recently, there have been emerging interests in its value as an anticancer agent (10 –13). Lovastatin has shown cytotoxic effects on ATC cell lines in several studies (14,15), and Zhong et al. reported that the cytotoxicity of lovastatin was mediated via Rho geranylgeranylation and RhoA/ROCK signaling (16). Although Holstein and Hohl reported that paclitaxel and lovastatin have synergistic cytotoxicity in leukemia cell lines, there have been no studies examining the effectiveness of this combination in ATC cell lines (17).
The purpose of this study was to evaluate the combined cytotoxicity of paclitaxel and lovastatin in an ATC cell line.
Materials and Methods
Cell culture and chemicals
The human ATC cell lines 8505C and BHT-101 were purchased from Deutsche Sammlung von Mikroorganismen und Zellculturen GmbH. It has been confirmed that the 8505C cell line and BHT-101 cell line are the original ATC cell line using DNA profiling analysis (18). The cells were cultured at 37°C in 5% CO2 and 95% humidity in RPMI 1640 medium (Sigma) supplemented with 10% heat-inactivated fetal calf serum, 100 U/mL of penicillin, and 100 μg/mL of streptomycin. Paclitaxel and lovastatin were kindly provided by Hanmi Pharmaceutical and dissolved in dimethyl sulfoxide (Carl ROTH). The stock solutions of these drugs were then diluted in complete culture medium to the appropriate concentration. Cells were plated (5 × 105 cells in 500 μL/well) in a 96-well plate and incubated with various concentrations of paclitaxel and lovastatin at 37°C in 5% CO2 and 95% humidity for 72 hours.
MTT assay
The MTT (Promega) assay was performed as recommended by the manufacturer. After 72 hours of culture with paclitaxel and/or lovastatin, the dye solution was added to each well. After an additional incubation at 37°C for 4 hours, the solubilizing solution/stop mix was added to each well. After an overnight incubation at 37°C, the plates were analyzed on a microplate spectrophotometer (Multiskan spectrum; Thermo Labsystem) at 570 nm. The absorbance of control cells (incubated without drugs) was defined as 100% (19).
Isobologram analysis
The data from the MTT assay were analyzed using CalcuSyn software (Biosoft). This software analyzes the combined effects of drugs, automatically determining synergism and antagonism. It performs multiple drug dose-effect calculations using the median effect methods described by Chou and Talalay (20). In the isobologram graph, there is considered to be an antagonistic effect when the point is above the oblique line, a synergistic effect when the point is beneath the oblique line, and an additive effect when the point is on the line. A combination index (CI) less than 1.0 is indicative of synergy, whereas an additive effect results in a CI equal to 1.0, and antagonism results in a CI greater than 1.0.
Detection of apoptotic cells
Annexin V-FITC was used to quantitatively determine the percentage of cells within a population that were actively undergoing apoptosis. Annexin V-FITC and propidium iodide (PI) staining was performed using the detection kit according to the manufacturer's protocol (BD Pharmingen). A total of 1 × 106 cells/mL were incubated with various concentrations of paclitaxel and/or lovastatin for 72 hours. The cells were washed with cold PBS and suspended in 100 μL buffer (10 mM HEPES/NaOH (pH 7.4), 140 mM NaCl, 2.5 mM CaCl2). After 5 μL of annexin V-FITC and 5 μL of PI were added, the cells were incubated for 15 minutes at room temperature (25°C) in the dark. After this incubation, 400 μL of binding buffer solution was added, and flow cytometry was performed within one hour using an FACS Canto II (Becton Dickinson). The quadrant containing Annexin V-FITC–positive/PI-negative cells represents early apoptotic cells, and the quadrant containing Annexin V-FITC–positive and PI–positive cells represents cells undergoing the end stage of apoptosis or necrosis or already dead cells.
Results
Cytotoxicity of paclitaxel and lovastatin on ATC cells
8505C cells were treated with lovastatin at various concentrations (0, 5, 10, and 20 μM), and viability was measured using an MTT assay. The viability of 8505C cells was inhibited by lovastatin after 72 hours of treatment in a concentration-dependent manner (p = 0.002, Fig. 1, left). The IC50 of lovastatin was 3.53 μM. At a concentration of 5 μM, lovastatin decreased viability to 53.2% compared with the controls after 72 hours, and at 20 μM, viability was reduced to 87.0% compared with the controls. 8505C cells were also treated with paclitaxel at various concentrations (0, 5, 10, and 20 nM). After treatment with paclitaxel, 8505C viability was also inhibited in a concentration-dependent manner (p = 0.020, Fig. 1, right). The IC50 of paclitaxel was 5.98 nM. At a concentration of 5 nM, 10 nM, and 20 nM, paclitaxel decreased viability by 51.9%, 80.5%, and 85.8%, respectively, compared with the controls.
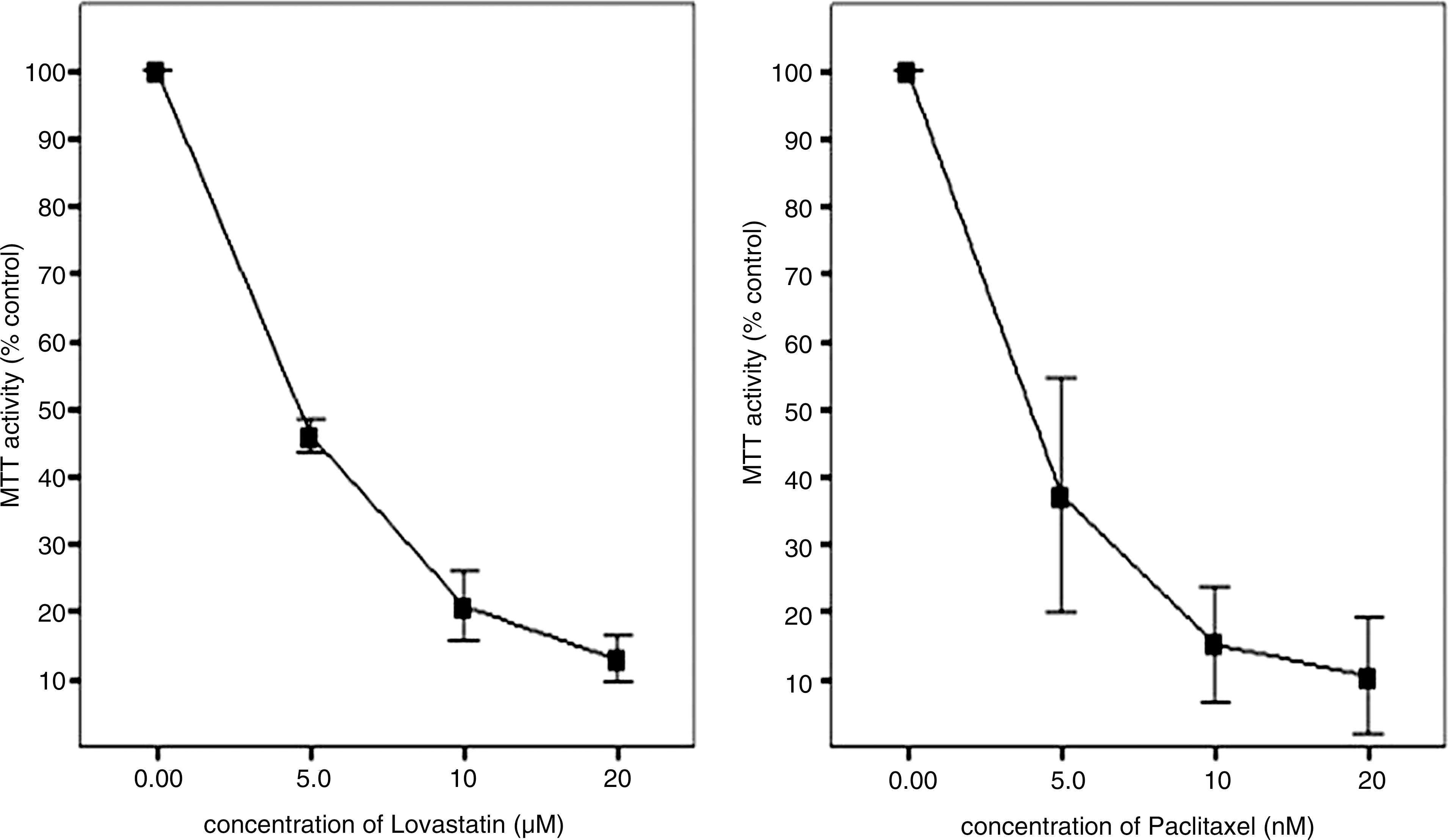
Cytotoxic effects of lovastatin (left) and paclitaxel (right) on 8505C anaplastic thyroid cancer cell line. Cells are grown in monolayer culture and exposed to lovastatin (at 0, 5, 10, and 20 μM) and paclitaxel (at 0, 5, 10, and 20 nM).
BHT-101 cells were treated with lovastatin at various concentrations (0, 5, 10, 20, 30, and 40 μM), and viability was measured using an MTT assay. The viability of BHT-101 cells was inhibited by lovastatin after 72 hours of treatment in a concentration-dependent manner (p = 0.020, Fig. 2, left). The IC50 of lovastatin was 17.13 μM. BHT-101 cells were also treated with paclitaxel at various concentrations (0, 2.5, 5, 10, 15, and 20 nM). After treatment with paclitaxel, BHT-101 viability was also inhibited in a concentration-dependent manner (p = 0.032, Fig. 2, right). The IC50 of paclitaxel was 35.26 nM.
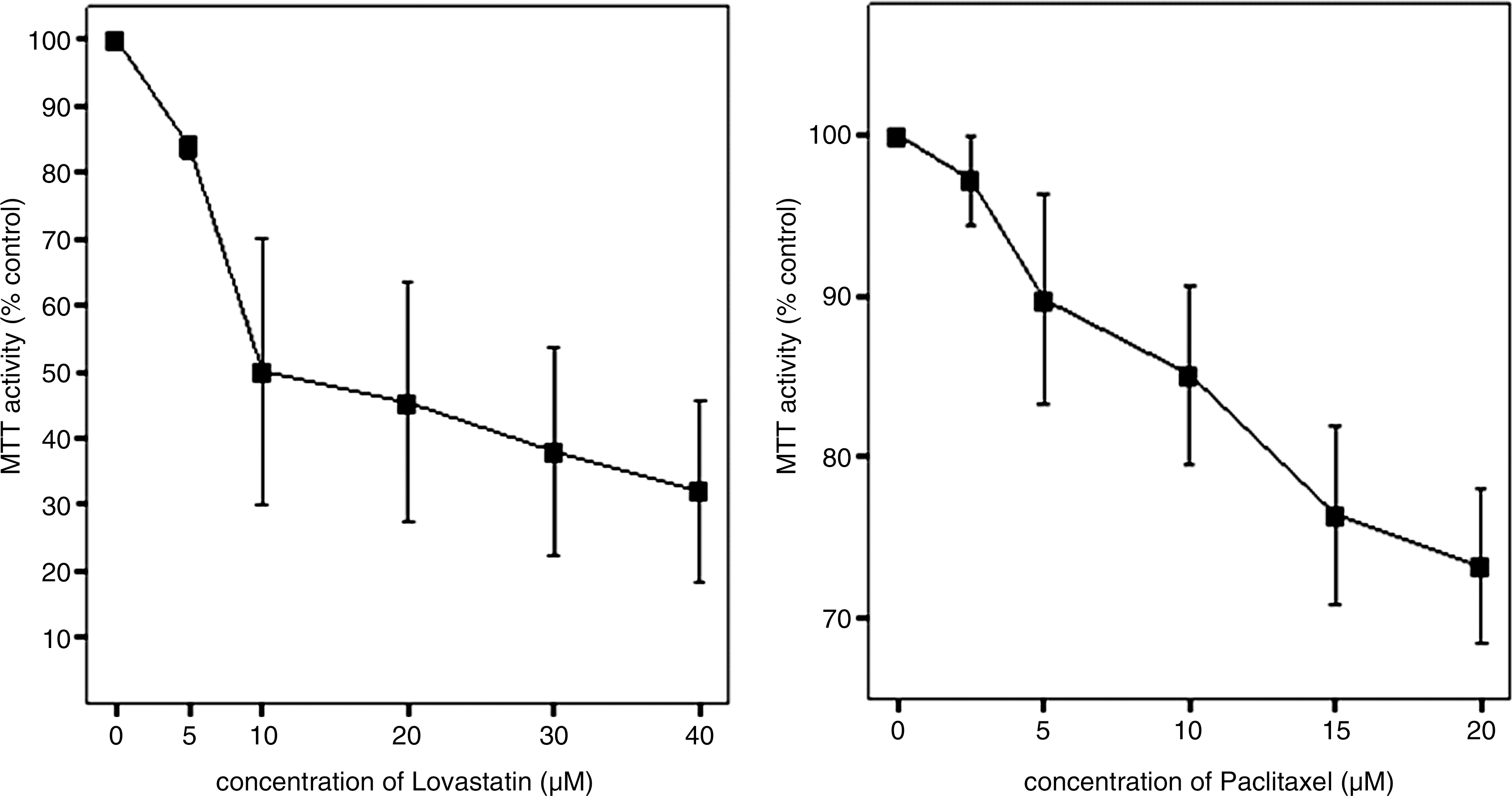
Cytotoxic effects of lovastatin (left) and paclitaxel (right) on BHT-101 anaplastic thyroid cancer cell line. Cells are grown in monolayer culture and exposed to lovastatin (0, 5, 10, 20, 30, and 40 μM) and paclitaxel (0, 2.5, 5, 10, 15, and 20 nM).
Combinatorial effects of paclitaxel and lovastatin on the 8505C cell line
An isobologram analysis was performed to determine whether there were interactions between paclitaxel and lovastatin. Since the point in the graph was above the oblique line, there was an antagonistic effect in 8505C cell line. The CI was 3.19 at ED50, 2.10 at ED75, and 1.48 at ED90, demonstrating that there was an antagonistic interaction between paclitaxel and lovastatin at ED50, ED75, and ED90 (Fig. 3a). In BHT-101 cells, however, there was a little different result. They showed an antagonistic effect (CI was 1.54) at ED50, an additive effect at ED75 (CI was 0.94), and a synergistic effect at ED90 (CI was 0.62) (Fig. 3b).

Results of analysis of apoptosis
Lovastatin-induced apoptosis of 8505C cells increased in a concentration-dependent manner (Fig. 4, left). The increased level of early apoptosis and end stage of apoptosis/necrosis was statistically significant after lovastatin treatment (p = 0.011, Table 1).
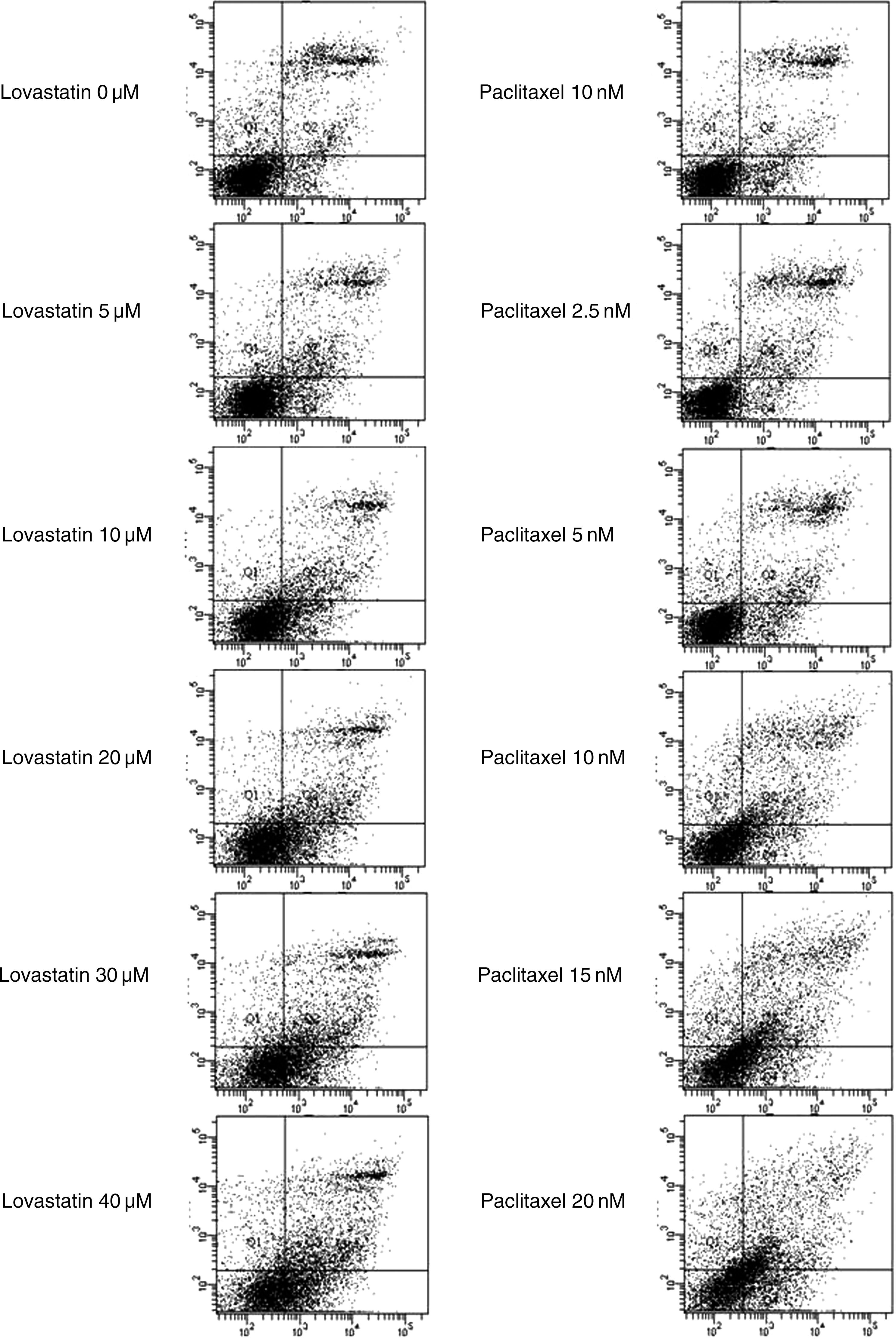
Apoptotic effect of lovastatin (left) and paclitaxel (right) on 8505C anaplastic thyroid cancer cell line. Horizontal line means annexin V-FITC staining and vertical line means PI staining. Viable cells (the quadrant of annexin V-FITC negative and PI negative) decreased concentration dependently. Early apoptotic cells (the quadrant of annexin V-FITC positive and PI negative) and late apoptotic/necrotic cells (the quadrant of annexin V-FITC positive and PI positive) increased concentration dependently. PI, propidium iodide.
8505C cells were incubated with lovastatin. Control cells were incubated without drug under the same environment. p < 0.05 was considered significant (n = 3).
LV, lovastatin; Lt, left; Rt, right.
The apoptosis of 8505C cells in the presence of paclitaxel increased in a concentration-dependent manner as well (Fig. 4, right). The increase in the proportion of cells undergoing early apoptosis was statistically significant after paclitaxel treatment (p = 0.014, Table 2). However, the increase in the proportion of cells undergoing late apoptosis/necrosis was not significant (p = 0.139, Table 2).
8505C cells were incubated with paclitaxel. Control cells were incubated without drug under the same environment. p < 0.05 was considered significant (n = 3).
PC, paclitaxel.
When 5 μM of lovastatin was added to various concentrations of paclitaxel, viable cells (Annexin V-FITC negative and PI negative) were more abundant than when only paclitaxel was added in 8505C cells (Fig. 5). Similarly, there were fewer early and late apoptotic cells when both paclitaxel and lovastatin were added to 8505C cells than when only paclitaxel was added (Fig. 5). The quantification of the annexin V-FITC and PI staining is presented in Table 3.
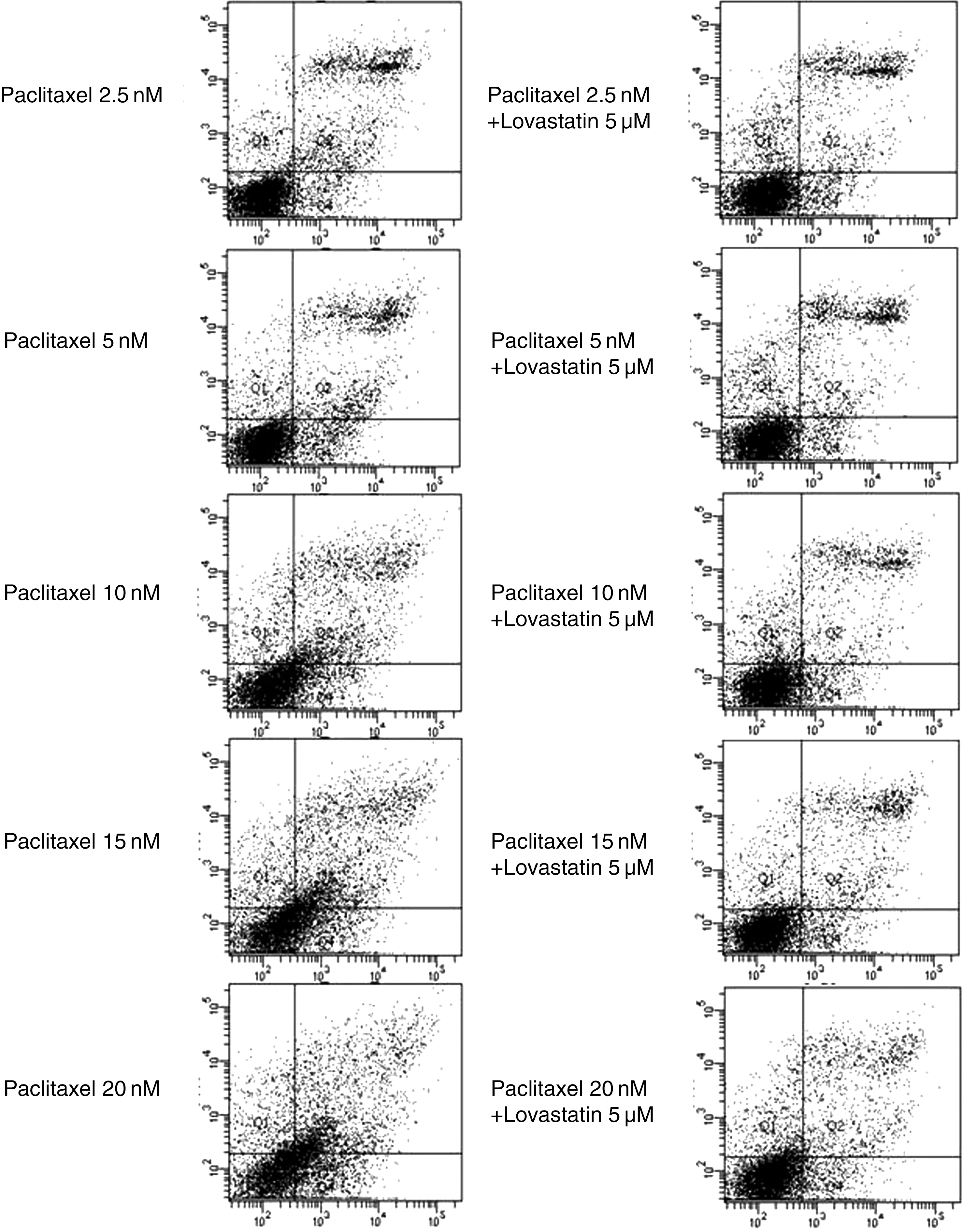
Apoptotic effect of paclitaxel (left) and paclitaxel plus lovastatin (right) on 8505C anaplastic thyroid cancer cell line. Horizontal line means annexin V-FITC staining and vertical line means PI staining.
8505C cells were incubated with paclitaxel only or paclitaxel plus lovastatin. Control cells were incubated without drug under the same environment.
In BHT-101 cells, both the paclitaxel and lovastatin did not show a concentration-dependent increase of apoptosis of the ATC cells (Fig. 6, left: lovastatin and right: paclitaxel). The increased level of early apoptosis and end stage of apoptosis/necrosis was not statistically significant after lovastatin treatment (p = 0.808 and p = 0.221, respectively). Similarly, the increased level of neither of the early apoptosis and end stage of apoptosis/necrosis was statistically significant after paclitaxel treatment (p = 0.911 and p = 0.522, respectively). The combination of the paclitaxel and lovastatin did not produce a synergistic or antagonistic effect (data are not shown).

Apoptotic effect of lovastatin (left) and paclitaxel (right) on BHT-101 anaplastic thyroid cancer cell line. Horizontal line means annexin V-FITC staining and vertical line means PI staining. Viable cells (the quadrant of annexin V-FITC negative and PI negative) did not decrease concentration dependently. Early apoptotic cells (the quadrant of annexin V-FITC positive and PI negative) and late apoptotic/necrotic cells (the quadrant of annexin V-FITC positive and PI positive) did not increase concentration dependently.
Discussion
Similar to other studies, this study demonstrates that the cytotoxic effects of paclitaxel and lovastatin increased in a concentration-dependent manner (6,14,21,22). In the 8505C cell line, lovastatin-induced apoptosis also increased in a concentration-dependent manner. Although it was not statistically significant, paclitaxel treatment also increased the level of apoptosis in ATC cells. However, the combination of paclitaxel and lovastatin resulted in antagonistic effects with regard to viability and apoptosis. The concentration-dependent cytotoxic effect of paclitaxel and lovastatin was found out in BHT-101 ATC cell line as well. However, the degree of cytotoxicity was weaker than that of the 8505C cell line, and a concentration-dependent increase of apoptosis was not produced in paclitaxel and lovastatin treatment. The combinatorial effect was diverse in accordance to concentration of paclitaxel and lovastatin.
There were different results in 8505c cells and BHT-101 cells. It has been confirmed that both of the 8505C and BHT-101 cell lines are the original ATC cell line using DNA profiling analysis (18). The 8505C cell line was established from the primary tumor of a 78-year-old woman with undifferentiated thyroid carcinoma. Histologically, the tumor was a largely papillary adenocarcinoma with spindle, polygonal, and giant cells. BHT-101 cell line was established from the lymph node metastasis of a 63-year-old woman with anaplastic papillary thyroid carcinoma. BHT-101 cells do not produce hormones, but they are partly positive for thyroiglobulin and thyroxine (23). Schweppe et al. reported that 8505C and BHT-101 are originated from anaplastic thyroid cancer and unique cell lines in each other. Although BHT-101 did not show BRAF V600E mutation and express TTF-1 transcription factor gene, 8505C showed BRAF V600E mutation and express TTF-1 in high level. Both the cell lines did not show RET/PTC1 rearrangement or Pax-8 transcription factor (18). Such a different character of the cell lines could be a possible cause of diverse results.
In BHT-101 cells, the IC50 of lovastatin was 17.13 μM and that of paclitaxel was 35.26 nM. They were much higher than the IC50 of lovastatin and paclitaxel in 8505C cells (3.53 μM in lovastatin and 5.98 nM in paclitaxel). We used less than 40 μM of lovastatin and 20 nM of paclitaxel in MTT assay, isobologram analysis, and apoptosis assay. Considering that extrapolation method was used in isobologram analysis, the combinatorial effect of lovastatin and paclitaxel on BHT-101 cell line was an antagonism in our experimental concentrations. Further, although there was cytotoxicity in BHT-101 cells, there was no apoptotic effect. If a higher concentration of paclitaxel and lovastatin was used, the results of apoptosis assay could be the same as that of 8505C cell line.
Paclitaxel binds to the β-tubulin subunit of microtubules and excessively stabilizes the microtubule. Therefore, active cell division during M phase is hindered in cancer cells by the presence of paclitaxel (5). Lovastatin is a 3-HMG-CoA reductase inhibitor and inhibits the rate-limiting step in the transformation of HMG-CoA into mevalonate in the mevalonate pathway. This pathway is important in the biosynthesis of isoprenoid molecules such as cholesterol, dolichol, and ubiquinone (24).
An important molecular target mediating the anti-neoplastic effect of lovastatin is the RhoA GTPase (25). The overexpression of RhoA GTPase is involved in cell cycle progression and cell growth, and in apoptosis and tumor initiation. Additionally, RhoA also functions in the assembly and disassembly of E-cadherin-mediated cell-to-cell junctions and, therefore, contributes to the epithelial-to-mesenchymal transition and metastasis. Moreover, the regulatory functions of RhoA on cytoskeleton remodeling can influence cancer-related processes such as invasion, migration, and metastasis (26).
RhoA needs to be geranylgeranylated by geranylgeranyl diphosphate, a downstream product of the mevalonate pathway (25). Geranylgeranyl diphosphate acts as a lipid anchor and allows RhoA to localize and attach to the plasma membrane, where it can be activated by guanine nucleotide exchange factors (GEF) (26). GEF-H1, the RhoA-specific GEF, is uniquely regulated by microtubule binding and is crucial for coupling microtubule dynamics to Rho-GTPase activation in a variety of normal biological situations. Since paclitaxel stabilizes microtubules, GEF-H1 is suppressed when paclitaxel is added. In several studies examining GEF-H1, paclitaxel was used to stabilize microtubules (27 –29).
Given this previous research, a synergistic effect of paclitaxel and lovastatin on anaplastic thyroid cancer cell lines was expected. However, contrary to our expectations, we observed an antagonistic effect in 8505C cells and various effects in BHT-101 cells in accordance with concentrations when the two drugs were combined. Several different mechanisms may underlie this observed antagonism. Wang et al. reported that lovastatin promotes the differentiation of ATC cells. At lower doses (25 μM), it promoted three-dimensional cytomorphological differentiation and also induced increased secretion of thyroglobulin by ATC cells (14). Several studies have demonstrated that tumors containing poorly differentiated cells had a better response to antitumor drugs than tumors with well-differentiated cells. These studies included head and neck cancers, soft tissue sarcomas, glioblastoma multiforme, malignant neuroendocrine tumors, and digestive tract malignancies (30 –34). Based on these studies, it is possible that the differentiation of cells exposed to lovastatin decreased the cytotoxicity of paclitaxel.
Second, lovastatin could affect the activity of paclitaxel through p21 Waf-1 in cell cycle. The anti-proliferative effects of lovastatin could be due to the enhancement of G1-S phase arrest via the increase in p21 Waf-1 and p27 Kip-1, two cyclin-dependent kinase inhibitors, as well as the inhibition of RhoA GTPase (35). In their research examining the differential role of p21 and p27 in the cell cycle, Schmidt et al. reported that the expression of p21 Waf-1 during the G2 phase impeded the entrance of cells into M phase during the period after D24851, a synthetic microtubule inhibitor, treatment. They said that it could contribute to D24851 resistance to cells (36). In another study, they reported that the expression of p21 Waf-1 markedly reduced paclitaxel-mediated mitotic arrest from 87.5% to 23% (37).
Bublik et al. investigated the mechanism to modulate p21 Waf-1 levels mediated by the hGTSE-1 (G2 and S phase-expressed-1 protein), a microtubule-localized protein whose expression in nontransformed cells is almost undetectable during the G1 phase, but increases during S phase and peaks in the G2 phase of the cell cycle (38). They previously demonstrated that hGTSE-1 down-regulates the p53 protein level, resulting in the inhibition of DNA damage-induced, p53-dependent apoptosis (39,40). They concluded that hGTSE-1-mediated p21 Waf-1 stabilization is also involved in the ability of cells to counteract paclitaxel-mediated cytotoxicity p53-independently (38). Therefore, we hypothesize that the lovastatin-enhanced p21 Waf-1 level could interfere with the anti-neoplastic effect of paclitaxel.
In contrast, Yang et al. reported that cells with both p21 Waf-1 alleles deleted were less sensitive to manumycin (a farnesyltransferase inhibitor) and paclitaxel than wild-type cells. The expression of p21 Waf-1 did not induce apoptosis but rather enhanced the cytotoxic effects of manumycin and paclitaxel in an ATC cell line. They proposed that p21 Waf-1 might be required to maintain cell sensitivity to the cytotoxic effects of manumycin and paclitaxel (41). However, further studies will be required to characterize the mechanism underlying the observed antagonism between paclitaxel and lovastatin.
Additionally, Lee et al. reported that the effectiveness of simvastatin combined with irinotecan, 5-fluorouracil, and leucovorin in the treatment of metastatic colorectal cancer. This was the first clinical trial to evaluate the antitumor activity of an HMG CoA inhibitor in combination with conventional cytotoxic chemotherapies for the treatment of a solid tumor. They found that low-dose simvastatin in combination with chemotherapeutic agents could prolong the time-to-progression of the patients without additional toxicity in comparison to the chemotherapeutic agents alone in other studies (42). Since this study was not a randomized study, however, the therapeutic effect could not be accepted completely, and the results of this study need to be further validated. Based on the above results and previous research, a clinical trial combining lovastatin and chemotherapeutic agents, particularly paclitaxel, in the treatment of ATC should be conducted carefully.
We investigated the combined effects of paclitaxel and lovastatin on ATC cell line. As the concentration of drug increased, each drug demonstrated increasing cytotoxicity when the cells were treated with individual drugs in 8505C and BHT-101 cell lines. Moreover, in 8505C cell line, each drug showed the concentration-dependent increase of apoptosis. However, the combination of paclitaxel and lovastatin resulted in antagonism that affected both cytotoxicity and apoptosis in 8505C cell line. In BHT-101 cells, the combinatorial effect was diverse along the concentration, and apoptotic effect was not produced. Our finding conclusively revealed that the combination of paclitaxel and lovastatin did not enhance the treatment effect in ATC cell lines. The mechanism of this antagonism between the two drugs requires further evaluation. The results of this study are important for guiding the development of a treatment for ATC.
Footnotes
Disclosure Statement
The authors declare that no competing financial interests exist.