
Abstract
Background:
The human pathogenic parvovirus B19 (B19) has recently been detected in papillary thyroid carcinoma (PTC) and Hashimoto's thyroiditis (HT) tissues at a high frequency in two studies of a Chinese cohort. We wanted to extend these data to include another cohort and expand the thyroid tumor tissue types assessed. In particular, we were interested to find whether B19 also infects anaplastic thyroid carcinoma (ATC), one of the most aggressive human cancers.
Methods:
Commercially available thyroid tumor tissue arrays were used to detect B19 capsid protein by immunohistochemistry in various types of thyroid tumors and disorders. The arrays were representative of the four main types of thyroid tumors, as well as other thyroid autoimmune disorders such as HT and Graves' disease, and adenomas, goiters, lymphomas, and normal thyroid tissue. In total, at least 12 different types of thyroid conditions as well as normal tissue were represented, many with multiple subjects.
Results:
Twenty-one of the 24 (88%) PTC tumors, 3 of the 3 ATC/undifferentiated tumors, and 3 of the 3 HT tissue samples were positive for B19 capsid protein by immunohistochemistry. The localization of the protein differed based on pathological disease type, with a nuclear to cytoplasmic shift seen from unaffected to tumor tissue.
Conclusions:
We extend the data available on B19 detection in the thyroid to show a high correlation of virus in another cohort of PTC and HT at the protein level. We also show, for the first time, B19 infection of much more highly aggressive ATC/undifferentiated tumors. Nuclear to cytoplasmic shift in B19 protein in cancer tissue suggests a possible link between B19 and thyroid cancer pathogenesis/progression.
Introduction
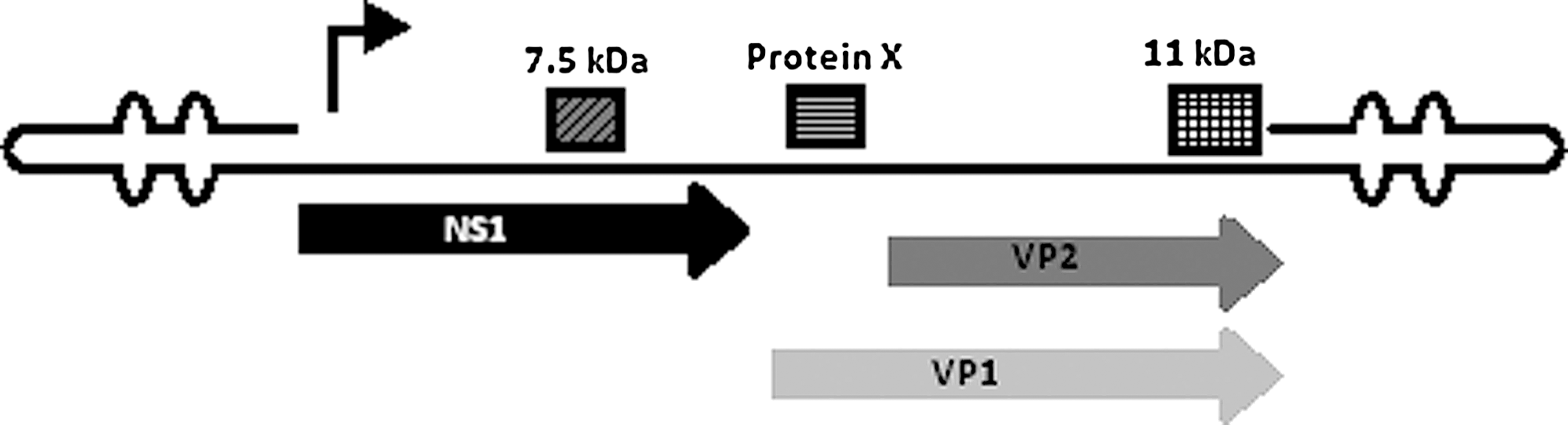
Schematic of the parvovirus B19 genome. Depiction of major proteins below (NS1 and VP1/VP2) and smaller proteins above (7.5 and 11 kDa) encoded by B19.
B19 has been shown to be a cause of several, wide-ranging human illnesses (2), including aplastic crisis, erythema infectiosum (fifth disease), arthritis, thrombocytopenia, hydrops fetalis, and myocarditis. B19 has also been highly associated with neurological disorders (3,4) and autoimmune disorders with symptoms similar to rheumatoid arthritis and lupus erythematosus (5). The role of B19 in autoimmune thyroid disorders, including Grave's disease (6) and Hashimoto's thyroiditis (HT) (7 –9), has also gained more recent attention. Little data are available on B19-thyroid interactions, but the related virus, minute virus of mice (MVMp), has been more extensively studied. MVMp NS1 protein has been shown to increase thyroid hormone receptor expression (10,11), a known oncogene and possible factor in thyroid tumor progression (12 –14). The Ras oncogene is not uncommonly mutated in papillary thyroid carcinoma (PTC) (15), and there is a significant literature linking non-B19 parvovirus replication with Ras mutations. Other PTC-related oncogenes (e.g., RET/PTC and BRAF) and their interactions with parvoviruses have thus far not been studied, and will prove to be good targets in understanding the B19-thyroid/thyroid cancer interaction.
B19 infection was recently shown to be highly associated with PTC (16). This striking correlation was determined by analyzing a relatively large number of thyroid tissues (n = 112) from various types of samples ranging from normal tissue to benign nodules to various carcinomas, including 38 PTCs of which 97% showed evidence of B19 DNA sequences and 63% showed evidence of capsid protein expression by immunohistochemistry (IHC) to assess for VP1. Significantly, B19 capsid proteins were not found (or showed nonsignificant staining) in other thyroid cancers (follicular thyroid carcinoma [FTC] or medullary thyroid carcinoma [MTC]) or surrounding normal tissues; anaplastic thyroid carcinoma (ATC) or other undifferentiated tumors were not assessed. A second strength of this prior work is that B19 infection was assessed in multiple ways, including nested PCR, but also by in situ hybridization (to verify the presence of B19 nucleic acids) and IHC for capsid proteins. To date, this is the only study showing a strong correlation with PTC and B19 infection. It is yet to be determined whether B19 is actively replicating in thyroid tissue, plays a role in thyroid cancer development or progression, or is merely present because of the cellular environment of these tumors.
PTCs account for about 80% of all thyroid cancers and tend to grow slowly, so the prognosis is usually good, but PTCs can recur (nearly 10% at 10 years) if not completely treated (17). This sharply contrasts with the less frequent ATCs, which have a frequency of only about 1% of new thyroid cancers, but only an approximate 20% survival rate after 1 year (18). Recurrent PTC typically occurs in older individuals and is also more aggressive leading to increased mortality. A recent study has shown that about 10% of recurrent PTC tumors have double mutations in common PTC-related oncogenes (RET/PTC and BRAF; see below), whereas primary PTCs usually only have one of these mutations (19). While there is a strong debate over whether ATC tumors arise from PTC, those that are believed to share similar characteristics have been found to usually also contain a double mutation, usually in the BRAF and p53 oncogenes. Some PTC variants also have particularly unfavorable prognoses: oxyphilic (Hürthle cell), tall cell, and columnar cell PTCs. Understanding whether factors such as B19 may contribute to the formation of these variants or serve as markers of progression will further enhance our knowledge of PTC and inform detection and treatment development.
Few diagnostic tools or treatments are currently available for ATCs. Further, patients with ATC have a median survival rate of only 3 months, with >75% of patients experiencing distant metastases (20). Although not all ATC tumors are derived from other follicular tumors, there is a correlation between BRAF mutations in some PTC (but not FTC) tumors and subsequent progression to the less differentiated anaplastic tumors (21). What drives this possible progression to a more aggressive phenotype is currently not known, but mutations of p53 have been found in ATC tumors with BRAF mutations and small areas of tissues with pathological features similar to PTC tumors (22). Some known B19-infected cells have also been associated with similar mutated oncogenes. B19-positive bone marrow giant proerythroblasts, a hallmark of B19 infection, contain a large nucleus and prominent nucleoli, and have been shown to be associated with p53 expression (23). Knowledge of whether the presence of B19 in PTC and ATC eventually leads to similar features and oncogene expression may greatly improve the diagnosis and treatment of more aggressive PTC and ATC tumors.
We aimed to determine if the B19 detection pattern in PTC and HT seen by the Wang group (16) was mirrored in a geographically different (non-Chinese) cohort and determine if other thyroid cancers such as ATC also had detectable amounts of B19 protein.
Materials and Methods
Thyroid tumor tissue samples
Thyroid tumor tissue arrays were obtained from BioChain and consisted of 6-μm-thick formalin-fixed, paraffin-embedded tissue sections mounted on positively charged slides. The first arrays contained 12 tumor samples and corresponding unaffected tissue from the same patient representing 9 PTC tumors of various grades, 1 ATC tumor, 1 FTC tumor, and 1 MTC tumor according to BioChain's diagnoses (Cat. No. Z7020097). The second contained 48 different thyroid tumors or related disorders, including 3 normal thyroid tissue sections, 2 lymphomas, 3 HT tissues, 2 Graves' disease tissues, 4 multinodular goiters, 23 PTC tumors, 2 undifferentiated carcinomas, 3 FTC tumors, and 1 MTC tumor (Cat. No. Z7020098). All sections on the 48 tumor array had duplicate samples from each patient. Tissue arrays were stored at 4°C until analyzed. Tumor diagnosis was assigned by the company using TMN classification of malignant tumors and the histological type reviewed by our pathologist (L.J.F.). An acutely infected fetal liver tissue sample was used as a positive control.
Immunohistochemistry
Tissue array slides (see above) were baked for 30 minutes at 60°C before use. Tissues were dehydrated using xylene and a descending alcohol series. Briefly, tissues were kept in xylene for two times 20 minutes, followed by 100% alcohol for two times 5 minutes, and then 95%, 90%, 80%, and 70% alcohol for 5 minutes each. Heat-induced antigen retrieval was performed in 10 mM sodium citrate (pH 6.0) for 10 minutes at 99°C. Slides were allowed to cool to room temperature before proceeding. Endogenous peroxidase activity was blocked using 0.3% hydrogen peroxide in Tris-buffered saline for 30 minutes at room temperature. Blocking and detecting solutions, secondary antibody, and diaminobenzidine developer were provided in an ImmunoChem Immunohistrochemistry Kit from BioChain (Cat. No. K3181100). Tissues were blocked in 1% normal serum provided by the BioChain kit in a phosphate-buffered saline (PBS) for 30 minutes at room temperature. Tissues were incubated in blocking solution containing a primary antibody detecting a common epitope of the B19 capsid proteins VP1/VP2 (MAB8293, 1:200, Chemicon, Billerica, MA) overnight at 4°C in a humid chamber. The sections were then rinsed three times for 5 minutes in PBS and incubated 30 minutes at room temperature in biotinylated anti-mouse immunoglobulin G (IgG) provided in the BioChain IHC kit followed by three additional 5-minute PBS washes. Tissues were incubated for 30 minutes in detection solution at room temperature, followed by three additional 5-minute washes in PBS.
IHC staining was revealed by diaminobenzidine liquid detect for 20 minutes. To identify the nucleus, cells were stained with hematoxylin (Electron Microscopy Sciences, Hatfield, PA). Images were taken on a Lecia DM 2500 upright microscope. To confirm correct antigen staining, an IgG control was performed alongside B19 capsid staining.
Results
Confirmation of B19 capsid protein detection in HT and PTC with novel detection in ATC
Two different thyroid cancer tissue arrays were analyzed for the presence of B19 capsid proteins using IHC. The results from these two tissue arrays are compiled in Table 1. This table represents all positive tissues detected in both arrays. While the second array of 48 thyroid disorders contained a large amount of samples, only the tumors and HT tissues listed in Table 1 were positive for viral proteins. Most importantly, this array contained normal tissue that was not taken from the same thyroid tissue as a tumor and this normal tissue was negative for B19 proteins (data not shown). B19 capsid protein was detected in HT samples as shown in Figure 2B (representative). This figure also shows the tissue control positively infected by B19 (Fig. 2A), as well as the extensive amount of B19 detected in the enlarged nuclei of the ATC tissues (Fig. 2C). Figure 2D is the IgG control for the ATC tissue shown in Figure 2C. These results corroborate that B19 infects HT and PTC, and we now show that ATC is also infected by B19.
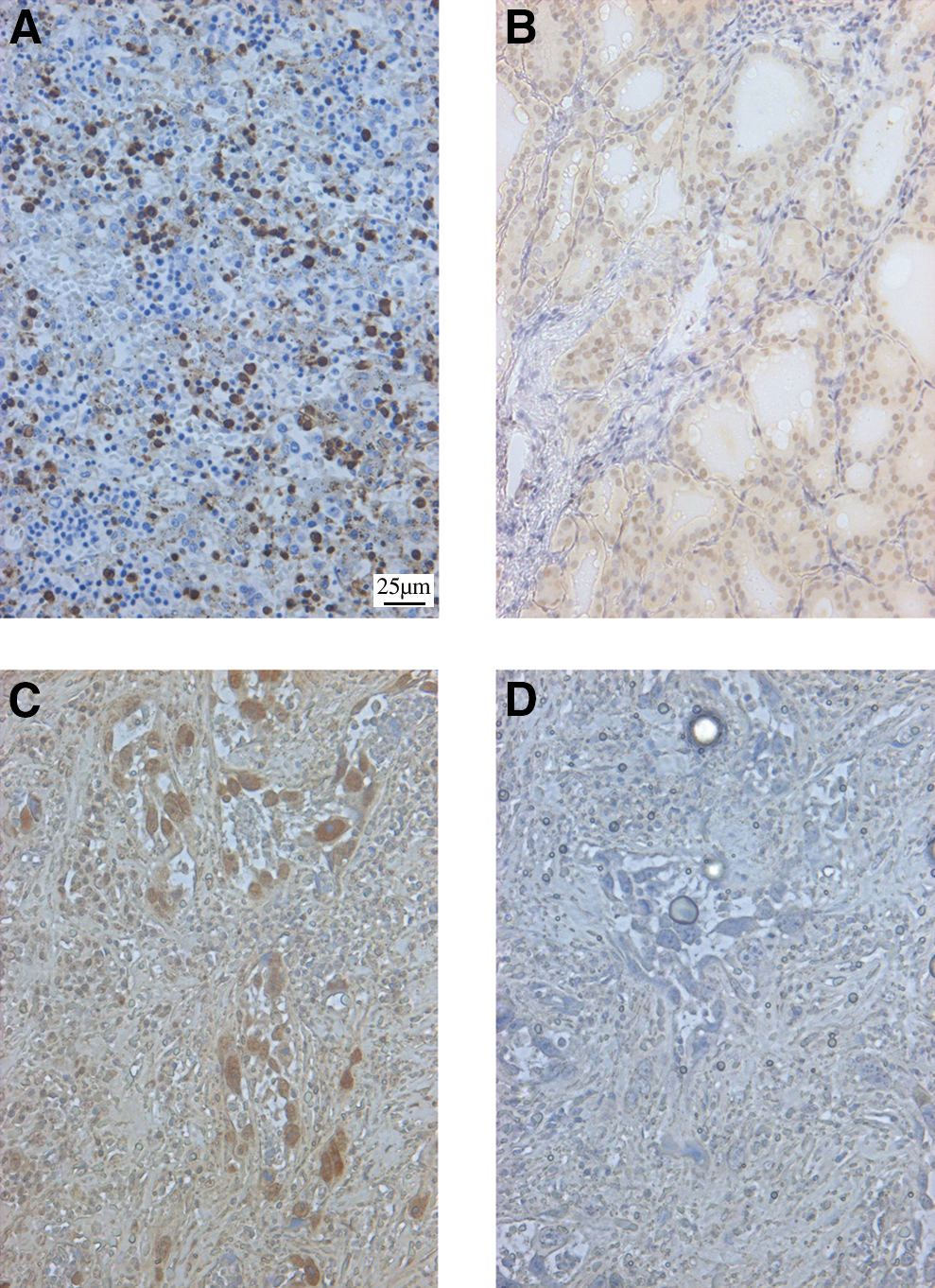
B19 detection in the thyroid.
Total data from the two tissue arrays were combined following IHC analyses.
ATC, anaplastic thyroid carcinoma; FTC, follicular thyroid carcinoma; IHC, immunohistochemistry; MTC, medullary thyroid carcinoma; PTC, papillary thyroid carcinoma.
Evaluation of various types of thyroid cancers for B19 capsid protein
Representative images from these thyroid tumors and unaffected surrounding tissues are shown in Figure 3. This figure illustrates the detection of B19 in 3 PTC tumors and adjacent unaffected tissue from the same patient (Fig. 3A–C and D–F, respectively). The adjacent tissue is described as normal tissue from the same patient, but many features of these tissue sections lack typical thyroid pathology. These tumors are classified, according to TNM classification, as grade I–II, T2N0M0 (Fig. 3A); II–III, T2N1M0 (Fig. 3B); and III, T2N0N0 (Fig. 3C). ATC, FTC, and MTC tumors and unaffected tissue are shown in Figure 3G–I and J–L, respectively. These tumors are classified as G: grade III, T2N1M0; H: T2N0M0; and I: III, T2N0N0. The cellular morphology in the FTC tumor was felt to have nuclear features suggestive of follicular variant of PTC, as assessed by our pathologist (L.J.F.). The tissue sample described as MTC also shows more nuclear variation than typical medullary carcinoma and was unable to be confirmed based upon the limited size and staining by our pathologist (L.J.F.). Gender and age of patients are as follows: male, 50 (Fig. 3A, D); female, 24 (Fig. 3B, E); male, 52 (Fig. 3C, F); female, 55 (Fig. 3G, J); female, 19 (Fig. 3H, K); and male, 16 (Fig. 3I, L). Unaffected tissues all showed nuclear staining for B19 capsid. Some PTC tumors and the ATC tumor also showed cytoplasmic virus capsid staining (although weaker in intensity) along with stronger/denser nuclear staining than normal background cells. Figure 3A shows the cytoplasmic and nuclear staining in the PTC tissues, as compared to Figure 3E, where the unaffected thyroid tissue shows only slightly weaker nuclear capsid detection. A few cells in the unaffected tissue from a PTC patient (Fig. 3F) appeared to also have cytoplasmic staining, but were felt to also show oncocytic metaplasia features possibly explaining the cytoplasmic staining. This staining may be positive B19 staining, or it may be a false-positive from IHC techniques. It has been shown that oncocytic cells (Hurthle cells) have a tendency for nonspecific antibody binding, which may be what is seen in Figure 3E (24). These results show that B19 is detected in a wide range, but not all, of thyroid tumors, and that there is a shift in localization of detected B19 capsid protein in unaffected and tumor tissue from the same patient (nuclear to cytoplasmic). This shift is seen in cells containing abnormal morphology.
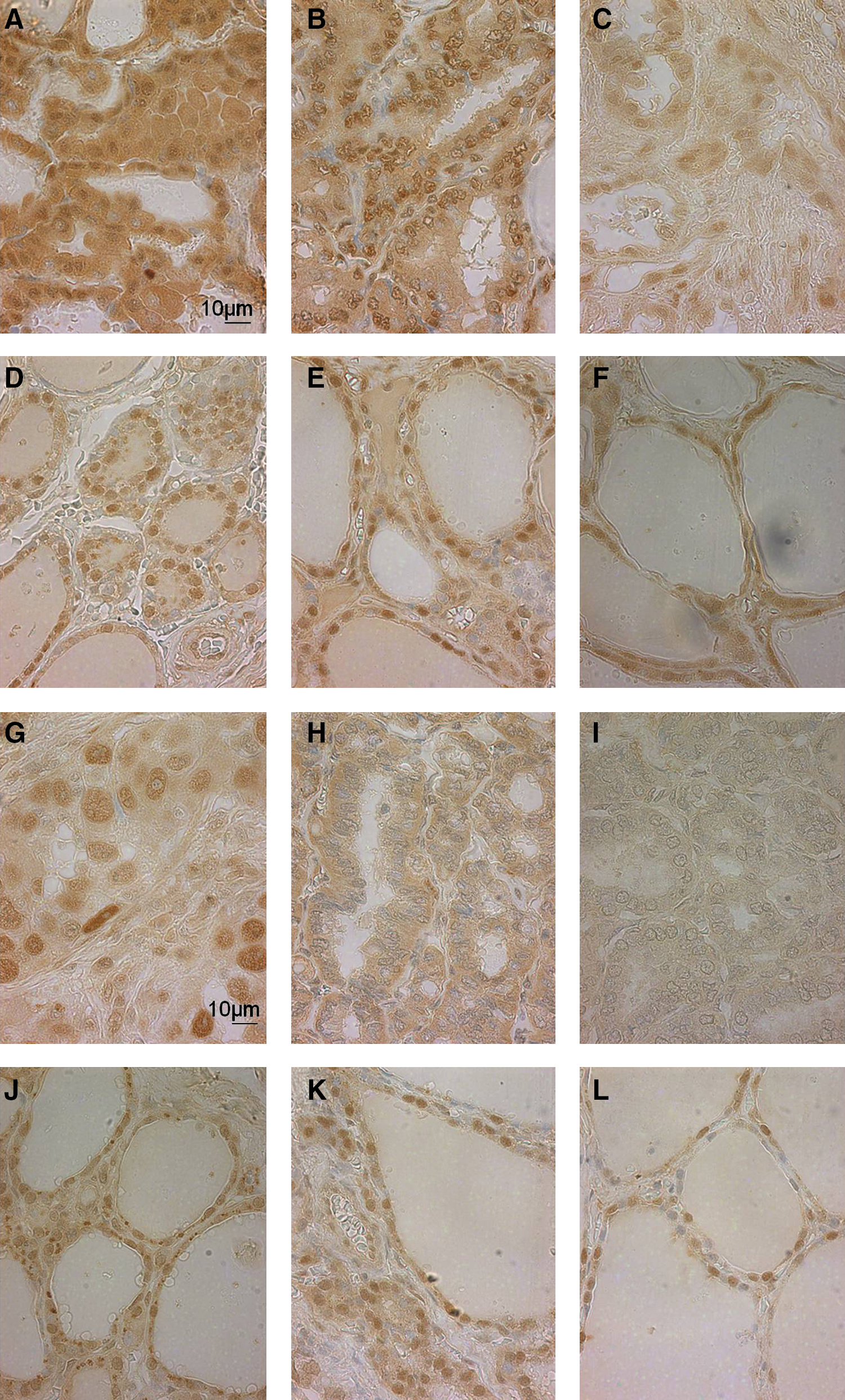
Immunohistochemistry detection of B19 capsid protein in thyroid tumors.
Nuclear features of B19-positive tumors
Using the same tissue array, three thyroid tumors, a lower-grade PTC tumor (II–III), a high-grade PTC tumor (III), and an ATC tumor (III), were examined under high magnification to determine any unique nuclear features that may correlate with B19 infection. The arrows in Figure 4A and 4B point out the distorted and enlarged nuclei seen in PTC tumors. This same pattern is not seen in unaffected tissue from the same patient as 4A (Fig. 4D). In the higher grade PTC tumor, some enlarged nuclei appear in both the tumor and unaffected tissue, and these enlarged nuclei are highly correlated with B19 protein detection. This pattern is further identified in the grade III ATC tumor in Figure 4C, where the largest nuclei appear to have the most intense B19 staining. These cells contain prominent nucleoli and uncondensed chromatin. By examining the nuclear features of B19-positive tumors and unaffected tissue, these results show a change in the staining intensity of nuclei in the tumors compared to the unaffected tissue.

Nuclear features of B19-positive tumors.
Discussion
This is the first study to show that the human pathogenic B19 persists in ATC, one of the most deadly human cancers, as evidenced by the presence of B19 capsid proteins in the tissue. This study also expands the findings of Wang et al. (7,16), detecting B19 in PTC and HT tissue samples, with similar results in other types of thyroid tumors. In the B19-positive tissues, we detected a marked change in virus capsid localization between unaffected tissues with only nuclear staining and tumor tissue with strong nuclear as well as cytoplasmic staining. The different staining patterns seen in the unaffected, tumor-adjacent tissue compared to the tumor tissue, within different PTC samples, and between PTC and ATC samples suggest a subset of tumors and cell types that contain an ideal cellular environment for B19 persistence.
The increasing studies of B19 detection in thyroid are significant because of the detection of not only DNA in these tumors (16), but also B19 protein. B19 DNA is known to persist in many tissues and organs (25): it has been found in the skin (26), cerebellum (27), fetal liver (28), testis (29), and heart tissue (30), as well as bone marrow cells (31), which are the most common cells for active replication. How this DNA persists in these various tissues is still unclear. To our knowledge, our study and those of Wang et al. (7,16) discussing B19 detection in the thyroid are the first to detect capsid protein by IHC in thyroid tissues believed to be nonpermissive to B19 infection. The increased presence of B19 capsid could be a result of more efficient B19 infection in these tumors, or of B19 replication, an important area of future investigation. Figure 4B and C demonstrates nuclear features of these B19 capsid protein-positive cells from aggressive thyroid tumors that resemble the enlarged nuclei seen in giant proerythroblasts, cells known to support B19 replication (32). Detection of B19 in these enlarged bone marrow cells has also been correlated with p53 expression (23), a common mutation in ATC tumors. Whether B19 detection and p53 mutation are found to be correlated in ATC tissues may be an indication of how B19 persists in the thyroid.
There is still a debate about whether ATCs develop de novo or from PTC tumors. Whether B19 is causal or merely takes advantage of this environment for persistence remains to be investigated. The relationship between HT and PTC development also remains elusive, with some HT tissues sharing many features with early PTC tumors (33). We have shown that nontumor thyroid tissue has detectable amounts of capsid proteins in the nucleus, whereas several types of tumors have capsid detection in both the nucleus and cytoplasm. B19 has been detected now in HT, PTC, and ATC tissue samples, as well as in the nontumor tissue from the same patients. Common mutations are known for PTC tumors (RET/PTC rearrangement, BRAF V600E, Ras point mutations, etc.), as well as in ATC tumors that are believed to have risen from PTC tumors (above mutations, p53). Other parvoviruses, such as rat parvovirus H-1, have been shown to have replication activity directly proportional to Ras expression, making oncogene interaction with B19 a plausible model (34,35). Because virus capsid protein was detected only in the nucleus of unaffected tissue from patients with a thyroid tumor, the environment created in the tumor compared to the adjacent tissue as it accumulates a particular mutation may allow for the increased detection seen in these tissues, as well as the translocation of detected protein to the cytoplasm. Understanding this interaction may lead to better understanding of thyroid tumor oncogene expression and gene mutations, and the role these play in development of specific thyroid tumors. This knowledge may lead to improved detection and prevention of more aggressive tumors such as ATC.
In our study, we found one of the two MTC tumors to be positive for B19 capsid proteins (not shown), in comparison to zero out of nine found to be positive in the Wang et al. (2008) study. Both studies represent a small cohort of MTC tumors from varied populations. The possibility exists that these tumors are genetically distinct, and may have different abilities to support B19 persistence. In addition, our pathologist (L.J.F.) determined that the MTC and FTC tumors had features that made them inconsistent with the BioChain pathology diagnosis. In the arrays, what is described as unaffected tissue is a sample from a patient with a thyroid tumor, so the morphology and characteristics of the tissue, while being unaffected, is most likely not completely normal thyroid tissue. This varied diagnosis may explain the discrepancy between the Wang study (16) and ours in relation to B19 detection in certain tissue types. Further studies with a larger sample size and increased amount of tissue linked to a clear, known case history of the patients are necessary to clarify this discrepancy by examining protein localization not only by IHC, but also by Western blot in conjunction with B19 DNA and RNA analysis.
This study is limited without serum samples from patients with a thyroid tumor and definitive medical history to determine prior virus infections and environmental influences. In addition, because each tumor was represented by a small area of tissue on the array, analysis in a larger tissue sample with more detailed pathological diagnosis could not be achieved. Detection of B19 proteins due to acute infection cannot be ruled out as we lacked matched serum samples, but is unlikely because of the ages of many of the patients from which the samples were collected. Because of the lack of commercial antibodies to B19's nonstructural protein NS1, studies so far have only been able to confirm the presence of B19 capsid proteins. This does not allow us to distinguish between productive virus replication and empty capsids, or to confirm virus replication. Detection of both B19 DNA and RNA, as well as NS1 protein detection, would significantly contribute to the study of B19 replication in the thyroid. How B19 infects HT, PTC, and ATC cells/tissues is unknown. Detection of virus capsid proteins in multiple tumor types from patients at various stages of treatment further counters the probability that detected proteins are a result of acute B19 infection. It is possible that B19 is replicating in other tissues (e.g., bone marrow) and homing to the rapidly dividing thyroid tumor tissue. Reactivation of latent viral infection is another possibility. Further studies are warranted to test these hypotheses and to determine the milieu of these B19-supportive cells and tumor types.
Whether B19 plays a role in transformation/progression from normal thyroid tumor tissue to HT, PTC, or ATC is yet to be determined. Determining what cellular factors are present in the tumors and cells that have increased detection of B19 will be key in understanding the role this virus may play in thyroid cancer pathogenesis. Because of the vast amount of tissue types in these arrays compared to the smaller amount of positive samples, B19 detection appears to be targeted to a subset of thyroid tumors and disorders. By understanding this specificity and association, we may be able to develop better diagnostic techniques for thyroid cancer classification, and develop improved screening and treatment tools.
In conclusion, we have extended the findings of Wang et al. (16) by detecting B19 protein by IHC in several types of thyroid tissues, tumors, and disorders, and have expanded the repertoire of B19-positive tumors to include the aggressive ATC. There is a distinct difference in the localization of detected capsid protein when unaffected tissue and tumor tissue were compared from the same patient, suggesting a shift in microenvironment that is ideal for B19 infection. These findings contribute to the knowledge of tissues with persistent B19 protein detection, and our knowledge of this virus' relationship with thyroid cancer.
Footnotes
Acknowledgments
The authors thank Marda Jorgensen (University of Florida Histology Core) for histology consultations. This work was supported by an American Cancer Society Chris DiMarco Institutional Research Pilot Project Grant to J.A.H. and the University of Florida Medical Guild Research Incentive Award to L.A.A.
Disclosure Statement
The authors declare that no competing financial interests exist.