
Abstract
Background:
In patients without metastases, capsular and vascular invasion must be noted to make the diagnosis of follicular thyroid carcinoma (FTC). Some patients are initially diagnosed as follicular adenoma (FA) but develop metastases, indicating the original lesion was FTC. A diagnostic marker for FTCs that appear to be FAs by conventional histopathology is urgently needed. CD147 is a transmembrane glycoprotein that induces matrix metalloproteinases (MMPs) and participates in carcinoma invasion. The objective of this study was to determine whether CD147 is upregulated in FTC and if measures directed against it could reduce the invasive activity of FTC cells.
Methods:
The expression levels of CD147, MMP-1, MMP-2, MMP-3, MMP-7, and MMP-9 in surgical specimens of normal thyroid (n=8), FA (n=20), and FTC (n=9) was determined using immunoblot and immunohistochemical techniques. CD147 protein expression levels of epithelial growth factor stimulated FTC-133 cell lines was measured by immunoblotting with and without cell signaling inhibitors such as wortmannin, PD98059, SP600125, and SB203580. This was also done after exposure to short-hairpin interference RNA directed against CD147.
Results:
Immunoblot analysis of thyroid tissues revealed significant increases in signals for CD147, MMP-3, MMP-7, and MMP-9 in FTC compared with FA or normal tissue, or both. Immunohistochemical analysis revealed colocalization of determinants of CD147 with those of all of MMPs studied, mainly in follicular cells in normal and neoplastic cells in FA and FTC; their immunoreactivities were to some extent more intense in the FTC than FA or normals. In FTC-133 cells, immunoreactive signals for CD147 were upregulated by epidermal growth factor (EGF), and the EGF-driven increases in CD147 were prevented by inhibitors against phosphoinositol-3 kinase (PI3K), extracellular signal-regulated protein kinase (ERK), or c-Jun N-terminal kinase (JNK) but not p38. RNA interference targeted against CD147 reduced the invasive activity of FTC-133 cells and was associated with downregulation of MMP-2, MMP-3, MMP-7, and MMP-9.
Conclusions:
These results provide in vivo evidence for CD147 upregulation in FTC and in vitro evidence for EGF-stimulated CD147 induction via the PI3K, ERK, and JNK pathways. They suggest the involvement of CD147 in the invasiveness of FTC cells via regulation of MMPs.
Introduction
Matrix metalloproteinase (MMP) is an endopeptidase that degrades the extracellular matrix, including the basement membrane. A growing body of evidence implicates MMPs in the invasion processes of cancer (2,3). MMPs are biosynthesized initially as proenzymes and subsequently activated by enzymatic cleavage. Over 20 isoforms of MMPs have been indentified in mammals (4). MMPs in the thyroid gland have been investigated in a number of studies of chronic thyroiditis and malignant neoplasms such as FTC (5,6). Expression of MMPs is mediated by several cell signaling enzymes including mitogen-activated protein kinase (MAPK), and transcription factors including nuclear factor-kappaB. They are also regulated by inflammatory or cell growth stimuli via proinflammatory cytokines, growth factors (7), and CD147 (8).
CD147, also known as extracellular MMP inducer, is a transmembrane glycoprotein that is categorized as a member of the immunoglobulin superfamily; it was first identified by Biswas (8). This protein is biosynthesized as an inactive form lacking glycosylation (27–29 kDa), becomes active following posttranslational glycosylation (35–65 kDa), translocates to the cell membrane, and becomes a membrane-bound form or a soluble form as a ligand of an unidentified receptor (9,10). To exert an effect, CD147 requires binding to its receptor, in an autocrine or paracrine manner. CD147 induces several MMPs, including collagenase (MMP-1), gelatinase A (MMP-2), stromelysin (MMP-3), gelatinase B (MMP-9), membrane type 1-MMP (MT1-MMP) (MMP-14), and MT2-MMP (MMP-15) (11 –16). In the case of neoplasms, CD147 is a tumor cell-derived MMP-inducer that stimulates both tumor cells and nearby fibroblasts and endothelial cells to produce MMPs, facilitating cancer cell invasion (10). There is emerging evidence for elevated CD147 levels in carcinomas of the urinary bladder (17), breast (18,19), lung (18,19), oral cavity (20), skin (21), esophagus (22), and thyroid (23 –25) in addition to malignant lymphoma (26,27) and malignant peripheral nerve sheath tumors (28). In immunohistochemical and immunocytochemical studies on papillary thyroid carcinoma, CD147 expression correlates with the degree of dedifferentiation (23,24). Another study on 156 differentiated thyroid carcinomas, including follicular and papillary carcinomas, showed that CD147 expression increased in thyroid carcinoma patients with distant or lymph node metastasis as compared with those without metastasis. In this study there was a correlation between CD147 expression and the pathological grading of tumor progression (25). The study also demonstrated that thyroid carcinoma patients with low CD147 expression had better survival rates (25). These observations suggest that CD147 plays a pivotal role in the invasive activity of thyroid carcinomas. However, little is known about whether CD147 has a role in the malignant behavior of FTC. To address this issue, we studied morphological and quantitative aspects of CD147 in surgically resected FTC tissues, including the mechanisms of CD147 induction and CD147-dependent invasiveness in cultured FTC cells. For this we employed signaling inhibition and RNA interference (RNAi) techniques.
Materials and Methods
Primary antibodies
The primary antibodies used in immunohistochemistry, immunocytochemistry, and immunoblotting were the following: mouse monoclonal IgG1 antibodies against CD147 (Cat. No. sc-21746 for immunohistochemistry and immunocytochemistry); (Cat. No. sc-53693 for immunoblotting), MMP-1 (Cat. No. F-67), MMP-2 (Cat. No. F-68), MMP-3 (Cat. No. F-66), MMP-7 (Cat. No. F-82), MMP-9 (Cat. No. F-69), epidermal growth factor receptor (EGFR) (Cat. No. K1492), extracellular signal-regulated kinase (ERK) phosphorylated at Tyr204 (p-ERK) (Cat. No. sc-7383), c-Jun N-terminal kinase (JNK) phosphorylated at Tyr183 and Tyr185 (p-JNK) (Cat. No. sc-6254), and β-actin (Cat. No. A-5441); goat polyclonal IgG antibodies against ERK (Cat. No. sc-94), JNK (Cat. No. sc-571), and Akt (Cat. No. sc-7126); and rabbit polyclonal IgG antibodies against Akt phosphorylated at Ser473 (p-Akt) (Cat. No. sc-7985-R), p38 MAPK(Cat. No. 506123), and p38 MAPK phosphorylated at Tyr180 and Tyr182 (p-p38 MAPK) (Cat. No. 506119). The antibodies to CD147, p-ERK, ERK, p-JNK, JNK, p-Akt, and Akt were purchased from Santa Cruz Biotechnology, the antibodies to the examined MMPs were from Daiichi Fine Chemical, the antibodies to p-p38 MAPK and p38 MAPK were from Calbiochem, the anti-EGFR antibody was from Dako, and the anti-β-actin antibody was from Sigma-Aldrich. According to the manufacturer's instructions, the anti-CD147 antibodies recognize both the nonglycosylated (NG-) and glycosylated (G-) forms of CD147, whereas the anti-MMP antibodies recognize both the precursor (P-) and cleaved (C-) forms of MMPs. In our study, while immunoreactive signals for total (T-) CD147 were defined as the sum of those for NG-CD147 and G-CD147, those for total (T-) MMP were defined as the sum of those for P-MMP and C-MMP.
Human subjects and tissue preparation
The design of our study using human tissues was approved by the Ethics Committee of Tokyo Women's Medical University, and performed after obtaining written informed consent of each subject. Thyroid gland tissue specimens were surgically resected from subjects at the Department of Endocrine Surgery of Tokyo Women's Medical University Hospital from 2001 to 2006. Of the cases examined, 9 were FTC cases (age: 25–66 years; sex: 3 males and 6 females), 20 were FA cases (age: 17–69 years; sex: 4 males and 16 females), and 8 were normal control cases (age: 27–65 years; sex: 1 male and 7 females); none of them were associated with any inflammatory disorder. There was no significant difference in ages between the normal, FA, and FTC groups (p>1.2) by nonrepeated measures analysis of variance (ANOVA). The pathological diagnosis of FTC was based on proliferation of tumor cells that lacked the characteristic nuclear features of papillary thyroid carcinoma and showed follicular arrangement in addition to capsular invasion, vascular invasion, or both. Of the nine FTC cases, six showed the combined capsular and vascular invasion pattern, one showed the capsular invasion without vascular invasion pattern, one showed the vascular invasion without capsular invasion pattern, and one had been initially diagnosed as having FA because of the lack of capsular and vascular invasion but was proven to be FTC after lymph node metastasis appeared 2 years after initial diagnosis. Of the FTC cases, six had distant metastasis, two had lymph node metastasis, and one did not have metastasis. The 20 FA cases did not have capsular or vascular invasion, local recurrence, or distant metastasis during follow-up periods of at least 4 years. Archival, formalin (20%)-fixed, paraffin-embedded materials stored at room temperature (RT) and freshly frozen materials stored at −80°C from each case were used for morphological and quantitative investigations, respectively.
Immunohistochemistry on human thyroid tissue sections
For morphological analysis, multiple 3-μm-thick sections were cut from each paraffin-embedded material, and used for hematoxylin-eosin (H&E) staining and immunohistochemical staining. For immunohistochemistry, sections were deparaffinized, rehydrated, quenched for 5 minutes at room temperature (RT) with 3% H2O2 for inhibiting endogenous peroxidase activity, rinsed in phosphate-buffered saline (PBS), pH 7.6, and pretreated for 30 minutes at RT with 5% skim milk in PBS for blocking nonspecific antibody binding. For unmasking antigens of CD147, MMP-2, MMP-3, and MMP-9, sections were processed with microwaving (95°C; 400 W; 20 minutes) in 1 mM ethylenediamine N,N,N′,N′-tetraacetic acid, pH 8.0 (EDTA) before skim milk pretreatment. They were subsequently incubated overnight at 4°C with the antibodies to CD147 at a dilution of 1:200, MMP-1 at a dilution of 1:200, MMP-2 at a dilution of 1:200, MMP-3 at a dilution of 1:50, MMP-7 at a dilution of 1:100, and MMP-9 at a dilution of 1:200. Antibody binding was visualized by the polymer-immunocomplex method using the Envision System (Dako). 3,3′-Diaminobenzidine tetrahydrochloride (DAB) was used as the chromogen, and hematoxylin, the counterstain. Sections processed with omission of the primary antibody or incubated with nonimmune serum derived from the same animal species producing the antibody served as negative controls.
Cell culture and immunocytochemistry
FTC-133 cells were obtained from the European Collection of Cell Culture (Health Protection Agency), and maintained in growth medium composed of a 1:1 mixture of Dulbecco's modified Eagle's medium (DMEM) and Ham's F12 (Gibco BRL) supplemented with 2 mM glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 10% fetal bovine serum (FBS) (Gibco) in a humidified, 5% CO2 incubator at 37°C. Cells were used for morphological and quantitative analyses. For morphological analysis, cells were fixed for 10 minutes at RT with 99% ethanol, and used for H&E staining and immunocytochemical staining. The cells were subsequently incubated overnight at 4°C with the anti-CD147 antibody at a dilution of 1:200, and antibody binding was visualized by the polymer-immunocomplex method, using the Envision System (Dako). EGFR immunoreactivity was detected with the EGFR pharmDx™ kit (Dako) according to the manufacturer's instructions. DAB and hematoxylin were used as the chromogen and the counterstain, respectively.
CD147 silencing and cell motility assay
For transient knockdown of the CD147 gene, a stealth select RNAi oligo targeting CD147 (si-CD147, GenBank Acc. No. AF320819; HSS141396) was designed and synthesized by Invitrogen as the following sequences: sense, 5′-AACUCACGAAGAACCUGCUCUCGGA-3′, and anti-sense 5′-UCCGAGAGCAGGUUCUUCGUGAGUU-3′. A stealth RNAi™ siRNA negative control Hi GC (Invitrogen) was used as a negative reaction control.
Cells were cultured in growth medium without antibiotics on 6 cm dishes until reaching 80% confluency. Transfection was then performed with 20 nM stealth si-CD147 that had been gently mixed and incubated for 20 minutes at RT with 8 μL of lipofectamine RNAiMAX (Invitrogen) and 800 μL of Opti-MEM (Invitrogen). The CD147 siRNA solution was added to the medium on each dish. The cells were incubated for 5 days. After harvesting, cells were stored at −80°C until use. Effects of negative control and CD147 siRNA on expression levels of CD147 and MMPs in FTC-133 cells were compared on immunoblots using cell lysates obtained 5 days after knockdown treatment.
Influence of RNAi on cell motility was evaluated by the transwell assay. Cells were incubated with siRNA in 6 cm diameter dishes at a density of 6×105 cells per dish within 5 days, and then were harvested and incubated in 24 well plates at a density of 2×105 cells per well with Matrigel-coated inserts for invasion (Beckton Dickenson). The upper and lower culture compartments were separated by polycarbonate filters with a pore size of 8 μm. After trypsinization, 2.5×104 cells were plated on each insert with 500 μL of serum-free medium. Growth medium containing 10% FBS was used as a chemoattractant in the bottom well. After incubating for 22 hours, cells on the upper surface were removed by scrubbing with a cotton swab, whereas those on the lower membrane were stained with H&E and the number was counted by light microscopy. Both the number of migrating and invading cells and the percentage of the mean numbers of invading cells through the Matrigel inserts in those of migrating cells through the uncoated inserts (% invasion in migration) in the si-CD147 treated and negative control groups were used for statistical analysis. RNAi treatment did not have any influence on cell morphology or cell growth during the observation period (22 hours) for the transwell assay (data not shown).
Identification of CD147 induction pathways
FTC-133 cells were plated and incubated in 35 mm diameter dishes for 24 hours at a density of 2×105 cells in growth medium. Subsequently, the medium was replaced by serum-free 5H medium, 1:1 mixture of DMEM and Ham's F12 supplemented with 10 μg/mL insulin, 5 μg/mL transferrin, 10 ng/mL somatostatin, 10 ng/mL Gly-His-Lys, and 400 pg/mL hydrocortisone, and cells were incubated for 24 hours. To identify the CD147 induction pathways, the cells were then pretreated with the following cell signaling inhibitors: wortmannin (2 μM), PD98059 (50 μM), SP600125 (2 μM), and SB203580 (5 μM), all of which were purchased from Calbiochem. Wortmannin, PD98059, SP600125, and SB203580 were used as inhibitors of the phosphoinositol-3 kinase (PI3K)-catalyzed phosphorylation of Akt, ERK MAPK kinase-catalyzed phosphorylation of ERK, JNK MAPK kinase-catalyzed phosphorylation of JNK, and the p38 MAPK kinase-catalyzed phosphorylation of p38, respectively. Preliminary examinations revealed no effect of the cell signaling inhibitors on cell morphology or cell growth (data not shown). Cells were harvested 30 minutes after EGF stimulation for the detection of the signaling enzymes or 18 hours after for the detection of the CD147 isoforms, and then stored at −80°C until use. Effects of EGF stimulation and cell signaling inhibition on expression levels of CD147 and phosphorylated forms of the cell signaling enzymes in FTC-133 cells were verified by comparison of immunoblots using cell lysates. Vehicle cells were defined as FTC-133 cells incubated in 5H media alone.
Immunoblotting and densitometry
The primary antibodies employed in immunoblotting were the mouse antibodies against CD147 at a dilution of 1:1000, MMP-1 at a dilution of 1:1000, MMP-2 at a dilution of 1:500, MMP-3 at a dilution of 1:500, MMP-7 at a dilution of 1:200, MMP-9 at a dilution of 1:2000, p-ERK at a dilution of 1:1000, p-JNK at a dilution of 1:1000, and β-actin at a dilution of 1:10,000; the goat antibodies to ERK at a dilution of 1:1000, JNK at a dilution of 1:2000, p-Akt at a dilution of 1:3000, and Akt at a dilution of 1:2000; and the rabbit polyclonal IgG antibodies to p-p38 MAPK at a dilution of 1:2000 and p38 MAPK at a dilution of 1:1000.
Both the human thyroid materials and the cultured cell materials were used for immunoblotting. The freshly frozen thyroid specimens were homogenized in radioimmunoprecipitation assay buffer composed of Tris-buffered saline (TBS), pH 7.4, containing 5 mM EDTA, 10% glycerol, 0.1% sodium-dodecyl sulfate (SDS), 1% Triton X-100, and 0.5% sodium deoxycholate, supplemented with the protease inhibitor cocktail Complete Mini (Roche Diagnostics) and the phosphatase inhibitor cocktails 1 and 2 (Sigma-Aldrich), according to the manufacturer's instructions. The frozen FTC-133 cells were incubated for 10 minutes on ice with lysis buffer composed of PBS, containing 1% NP-40, 0.5% sodium deoxycholate, and 0.1% SDS.
Protein concentrations of these tissue and cell samples were determined by the Bradford method (29). Samples were boiled for 5 minutes at 100°C with four volumes of modified Laemmli's sample buffer composed of 150 mM TBS containing 25% glycerol, 5% SDS, 100 mM 1,4-dithiothreitol, and 0.05% bromophenol blue, and they were separated by 12% SDS-polyacrylamide gel electrophoresis (PAGE) for MMP-7 or by 10% SDS-PAGE for other proteins. Aliquots of these samples (20–30 μg protein per lane) were loaded on a gel, electrophoresed, and electroblotted onto a polyvinylidene difluoride (PVDF) blotting membrane (Millipore). To block nonspecific antibody binding, the PVDF membranes were pretreated for one hour at RT with 5% nonfat skim milk in TBS containing 0.1% Tween-20 (TBS-T) for CD147, MMP-3, p-ERK, ERK, p-JNK, p38, p-Akt, and Akt, or with 5% bovine serum albumin in TBS-T for MMP-1, MMP-2, MMP-7, MMP-9, and p-p38, or with 5% blocking agent (GE Healthcare) in TBS-T for JNK. To amplify immunoreaction sensitivity, the antibodies to MMP-1, MMP-3, MMP-9, p-JNK, JNK, p-p38, and p38 were diluted in Can Get Signal® (ToYoBo). Other antibodies were diluted in 5% skim milk in TBS-T. Blots were then incubated overnight at 4°C with the primary antibodies, followed by rabbit anti-mouse IgG labeled with horseradish peroxidase (HRP), HRP-labeled rabbit anti-goat IgG, or HRP-labeled donkey anti-rabbit IgG. Antibody binding was visualized by the chemiluminescence method using the ECL-plus kit (GE Healthcare). Blots from which the primary antibodies were omitted gave negative reaction controls. Immunoreactive signals on each lane were scanned with the Light-Capture Cooled Camera System® (ATTO) and imported into a personal computer. After detection of these immunoreactive signals, blots were treated with stripping buffer, composed of 62.5 mM Tris-HCl, pH 6.8, 2% SDS, 1% β-mercaptoethanol, and then reprobed with the anti-β-actin antibody or the antibodies to nonphosphorylated cell signaling enzymes followed by the secondary antibodies. Optical density was quantitatively measured using the CS analyzer® (ATTO).
Statistics
All values in the groups were expressed as mean±standard deviation, and statistically compared, using the Excel® Statistical Program File ystat2004.xls (Igaku Tosho). Densitometric data of NG-, G-, and T-CD147 and P-, C-, and T-MMPs normalized with β-actin on immunoblots using thyroid tissue samples and cell culture samples were compared by nonrepeated measures ANOVA between the normal thyroid tissue, FA, and FTC groups or between the EGF-stimulated groups in the absence and presence of pretreatment with cell signaling inhibitors, and followed by multiple comparisons with the post hoc Student–Newman–Keuls test. In the case of CD147 silencing experiments, densitometric data of NG-, G-, and T-CD147 and P-, C-, and T-MMPs normalized with β-actin on immunoblots and cell counting data in cell motility assay were compared by unpaired Student's t-test between the si-CD147-treated and normal control groups. All experiments using cell materials were in triplicate. Statistical significance was defined as p-values less than 0.05.
Results
Immunohistochemical localization of CD147 and MMPs in thyroid tissue sections
To verify tissue localizations of CD147 and MMPs, immunohistochemical analysis was performed, using their specific antibodies. No immunoreaction product was visible on negative reaction control sections. Immunoreactivities for CD147 (Fig. 1a–c), MMP-1 (Fig. 1d–f), MMP-2 (Fig. 1g–i), MMP-3 (Fig. 1j–l), MMP-7 (Fig. 1m–o), and MMP-9 (Fig. 1p–r) were mainly localized in follicular epithelial cells of the normal thyroid tissue sections and neoplastic cells of the FA and FTC tissue sections, with staining predominant in the cytoplasm (insets in Fig. 1). The immunoreactivities for CD147, MMP-3, MMP-7, and MMP-9 were only very weak or not at all in the normal group, while those seemed to be more intense in the FA and FTC groups to some extent. On the other hand, there was no significant difference for the immunoreactivities for MMP-1 and MMP-2 between the three groups.
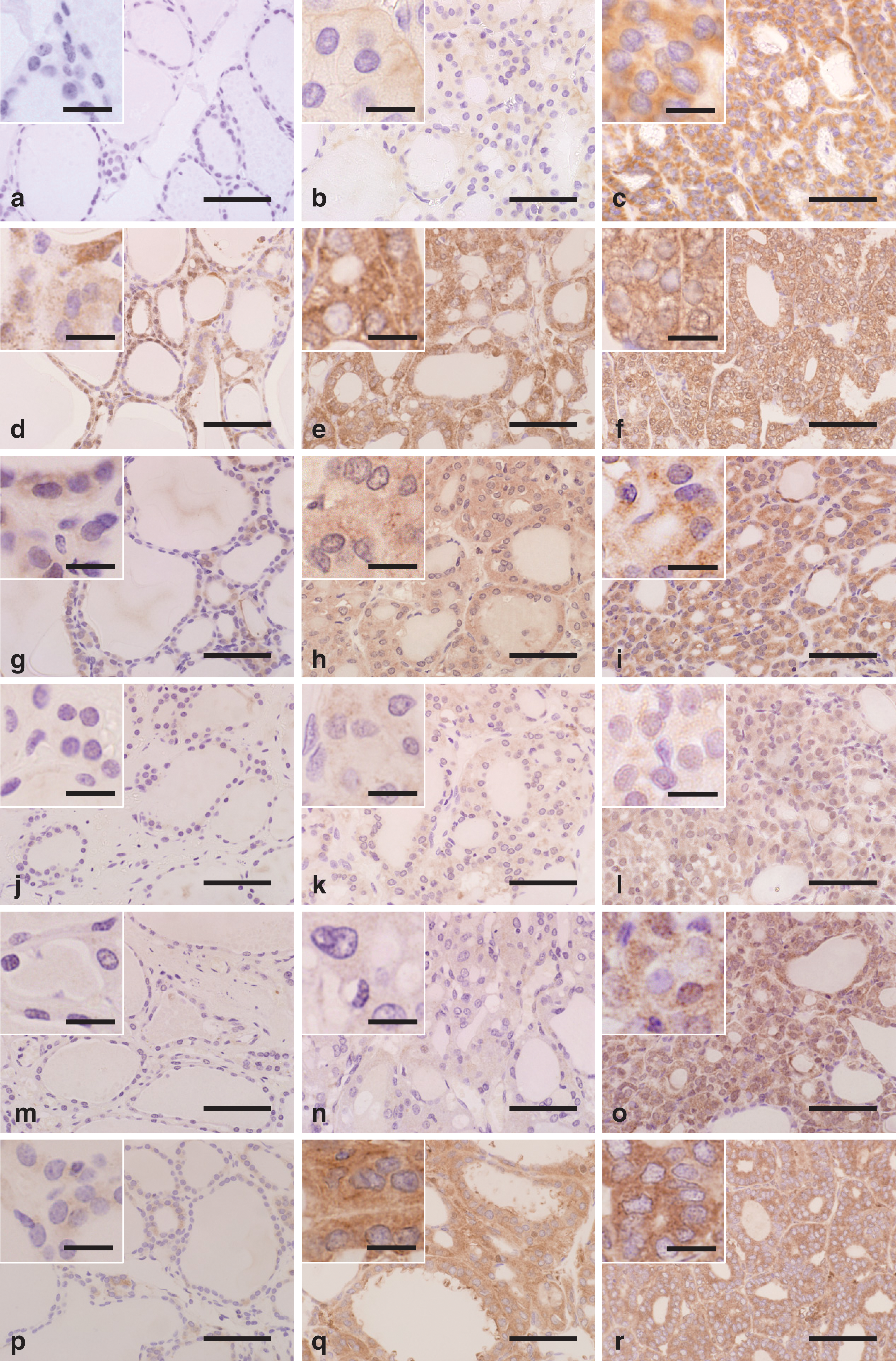
Representative microphotographs (the insets indicate magnified views) of formalin-fixed, paraffin-embedded thyroid gland tissue sections immunostained for CD147
Quantitative measurement of CD147 and MMPs in thyroid tissue samples
Intracellular levels of CD147 and MMPs in the normal, FA, and FTC groups were quantitatively measured by immunoblot and densitometric techniques. No immunoreaction product was visible on negative reaction control blots (data not shown). Both the G-CD147/β-actin optical density ratio (termed the normalized G-CD147 density) and the normalized T-CD147 staining density were significantly increased in the FTC group compared with the normal and FA groups (Fig. 2a). There was no significant difference in the normalized densities of C-MMP-1, T-MMP-1, C-MMP-2, or T-MMP-2 between the three groups (Fig. 2b, c). Both the normalized densities of P-MMP-3 and T-MMP-3 were significantly increased in the FTC group compared with the normal and FA groups (Fig. 2d). Both the normalized densities of P-MMP-7 and T-MMP-7 were significantly increased in the FTC group compared with the normal and FA groups (Fig. 2e). Both the normalized densities of P-MMP-9 and T-MMP-9 were significantly increased in the FA and FTC groups compared with the normal group (Fig. 2f).
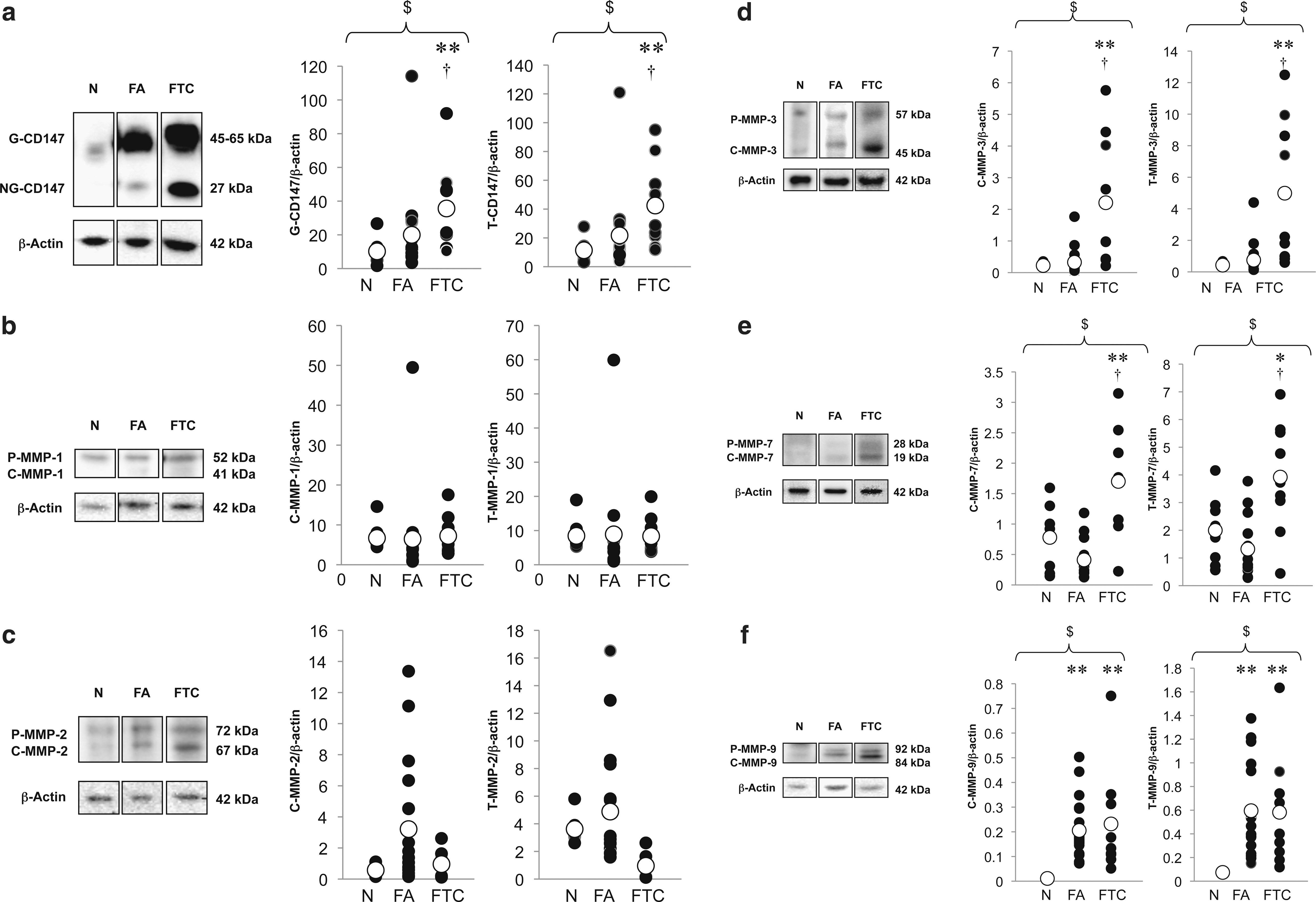
Representative immunoblots of CD147
Morphological observations of FTC-133 cells
On H&E-stained slides, FTC-133 cells showed polygonal or spindle cytoplasm and hyperchromatic nuclei with atypical mitoses (Fig. 3a). Immunoreactivities for EGFR (Fig. 3b) and CD147 (Fig. 3c) were detectable in the cytoplasm and prominent at the cell surface (insets of Fig. 3b, c). No immunoreaction product was visible on negative control slides (data not shown).
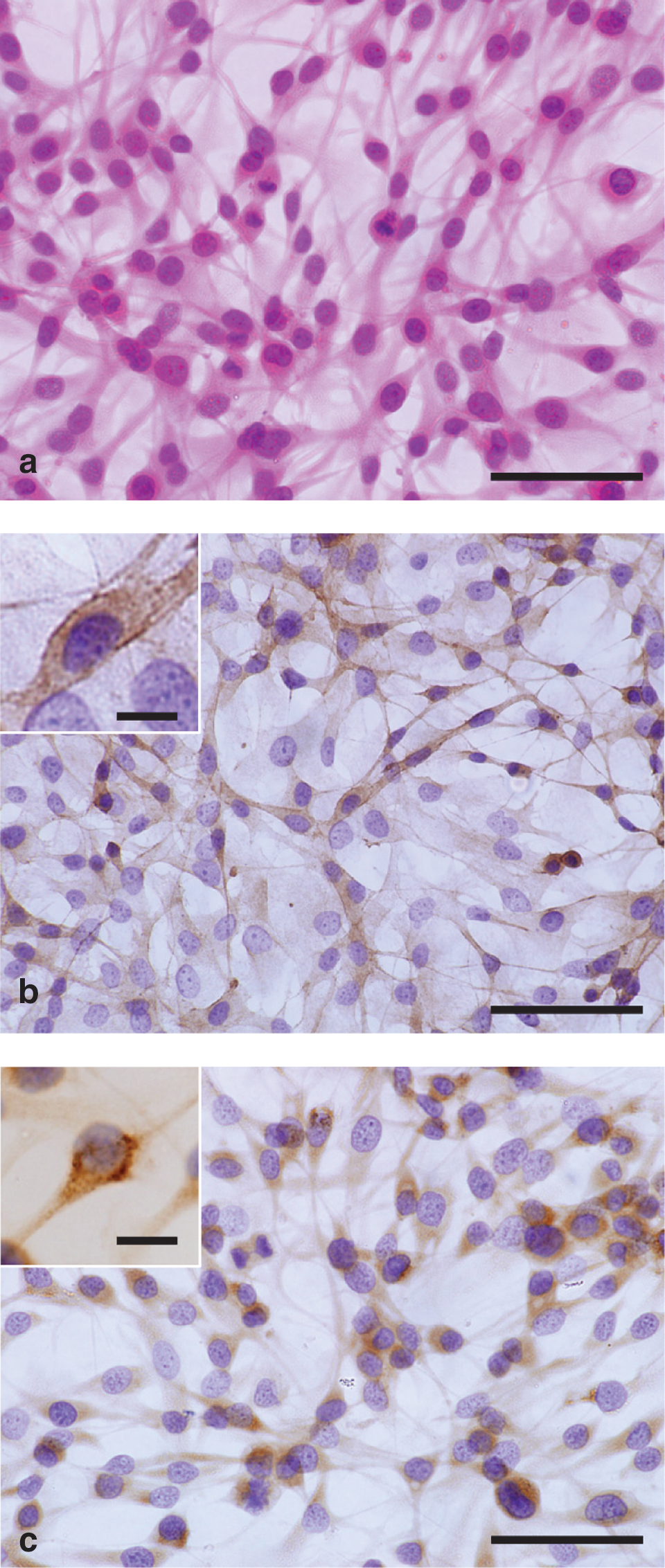
Microphotographs (the insets indicate magnified views) of FTC-133 cells stained with hematoxylin-eosin
EGF upregulates CD147 via PI3K and MAPK pathways
Increased levels of p-Akt, p-ERK, p-JNK, and p-p38 driven by EGF stimulation appeared optically reduced by pretreatment with their respective inhibitors, whereas the levels of Akt, ERK, JNK, and p38 were unaffected by the cell signaling inhibitors (Fig. 4a). These findings confirmed the efficacy of the inhibitors. Immunoreactive signals for NG-CD147, G-CD147 and β-actin appeared at 27, 45–65, and 42 kDa, respectively (Fig. 4b). The normalized staining densities of NG-CD147 (Fig. 4c), G-CD147 (Fig. 4d), and T-CD147 (Fig. 4e) were significantly increased in FTC-133 cells with EGF stimulation alone compared with vehicle cells. Staining was significantly reduced in EGF-stimulated FTC-133 cells after pretreatment with inhibitors against PI3K, MEK, and JNK but not p38 compared with cells having EGF stimulation alone.

Representative immunoblots of p-Akt, Akt, p-ERK, ERK, p-JNK, JNK, p-p38, and p38 in total lysates of EGF-stimulated FTC-133 cells obtained one hour after pretreatment with or without cell signaling inhibitors
CD147 silencing impedes cell invasion in association with downregulation of MMPs
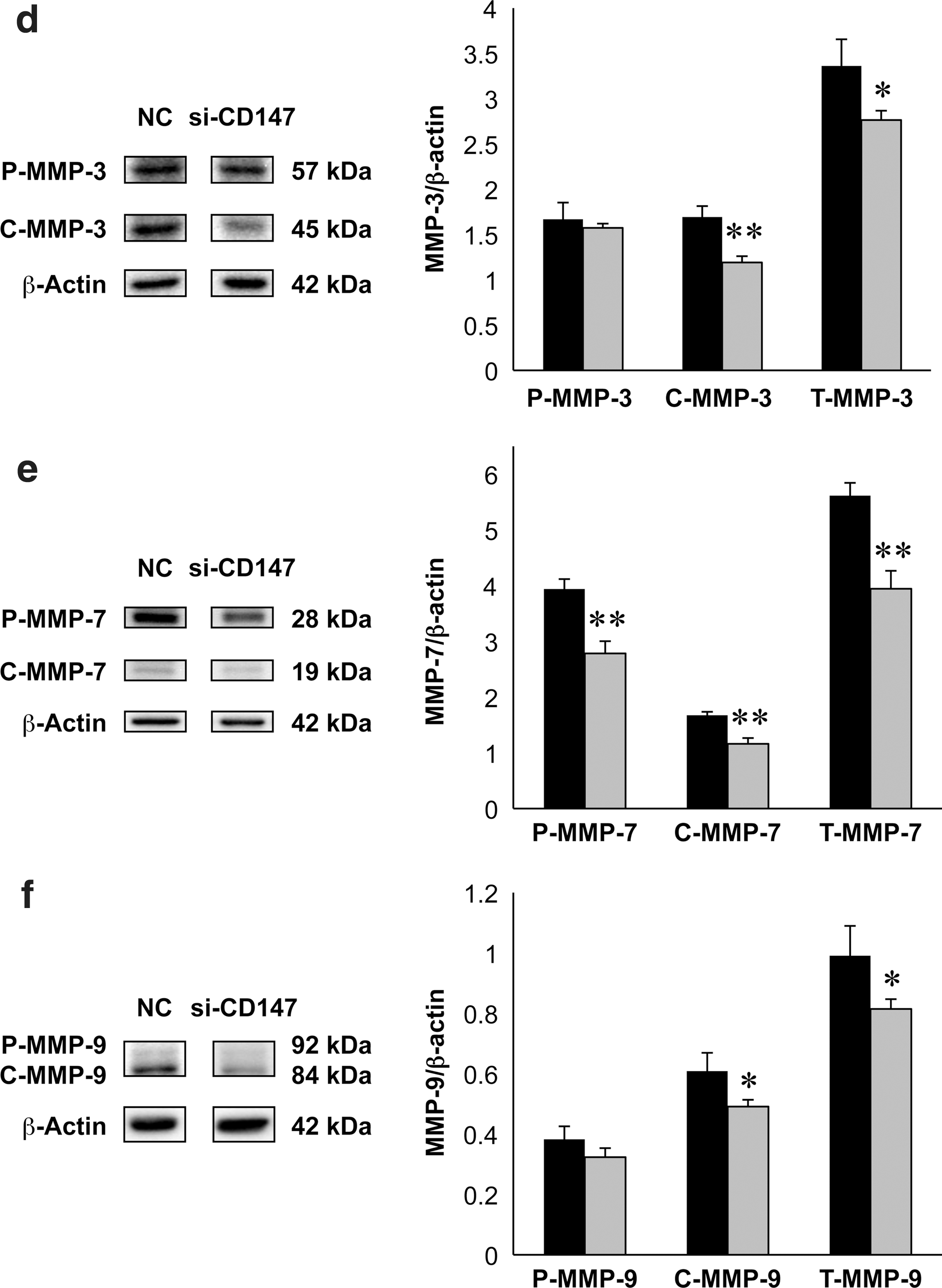
To elucidate the influence of CD147 silencing on the expression of MMPs, intracellular levels of CD147, and MMPs in total cell lysates, including the cytosolic and membranous components, were quantitatively measured on immunoblots using FTC-133 cells treated with or without CD147 RNAi. The normalized densities of NG-CD147, G-CD147, and T-CD147 was significantly reduced in the si-CD147-treated group compared with the normal control group (Fig. 5a). There were no significant differences in the normalized densities of P-MMP-1, C-MMP-1, or T-MMP-1 between the normal control and si-CD147-treated groups (Fig. 5b). The normalized densities of P-MMP-2, C-MMP-2, and T-MMP-2 were significantly reduced in the si-CD147-treated group compared with the normal control group (Fig. 5c). The normalized densities of C-MMP-3 and T-MMP-3 but not P-MMP-3 were significantly reduced in the si-CD147-treated group compared with the normal control group (Fig. 5d). The normalized densities of P-MMP-7, C-MMP-7, and T-MMP-7 were significantly reduced in the si-CD147-treated group compared with the normal control group (Fig. 5e). The normalized densities of C-MMP-9 and T-MMP-9 but not P-MMP-9 were significantly reduced in the si-CD147-treated group compared with the normal control group (Fig. 5f).
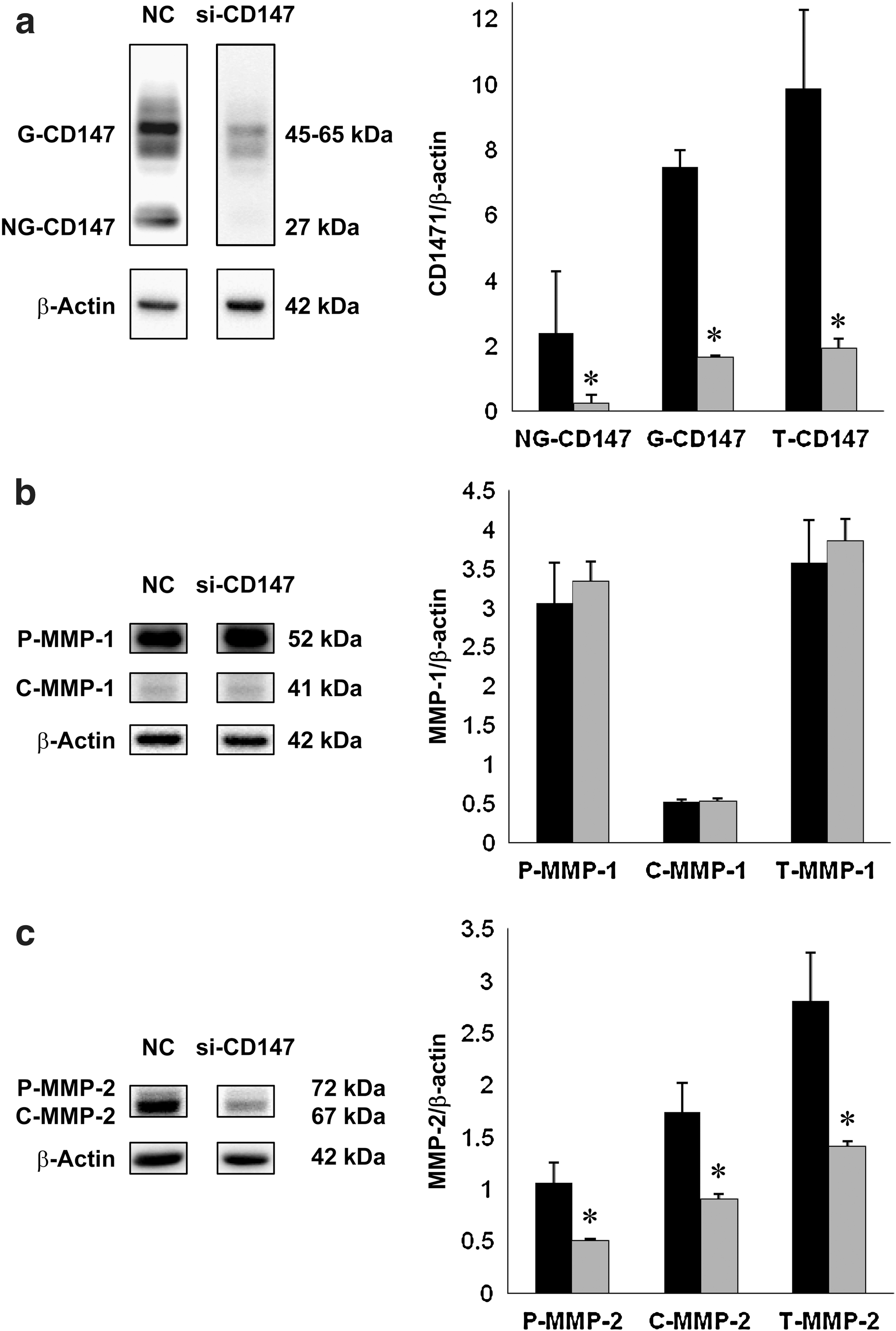
Representative immunoblots of CD147
To assess a role for CD147 in the invasive activity of FTC-133 cells, cell motility assays were performed. Morphologically, the number of invading cells on H&E-stained plates appeared smaller than that of migrating cells with or without RNAi treatment (Fig. 6a). Quantitatively, the % invasion in migration was significantly reduced in the si-CD147 treated group compared with the normal control group (Fig. 6b).
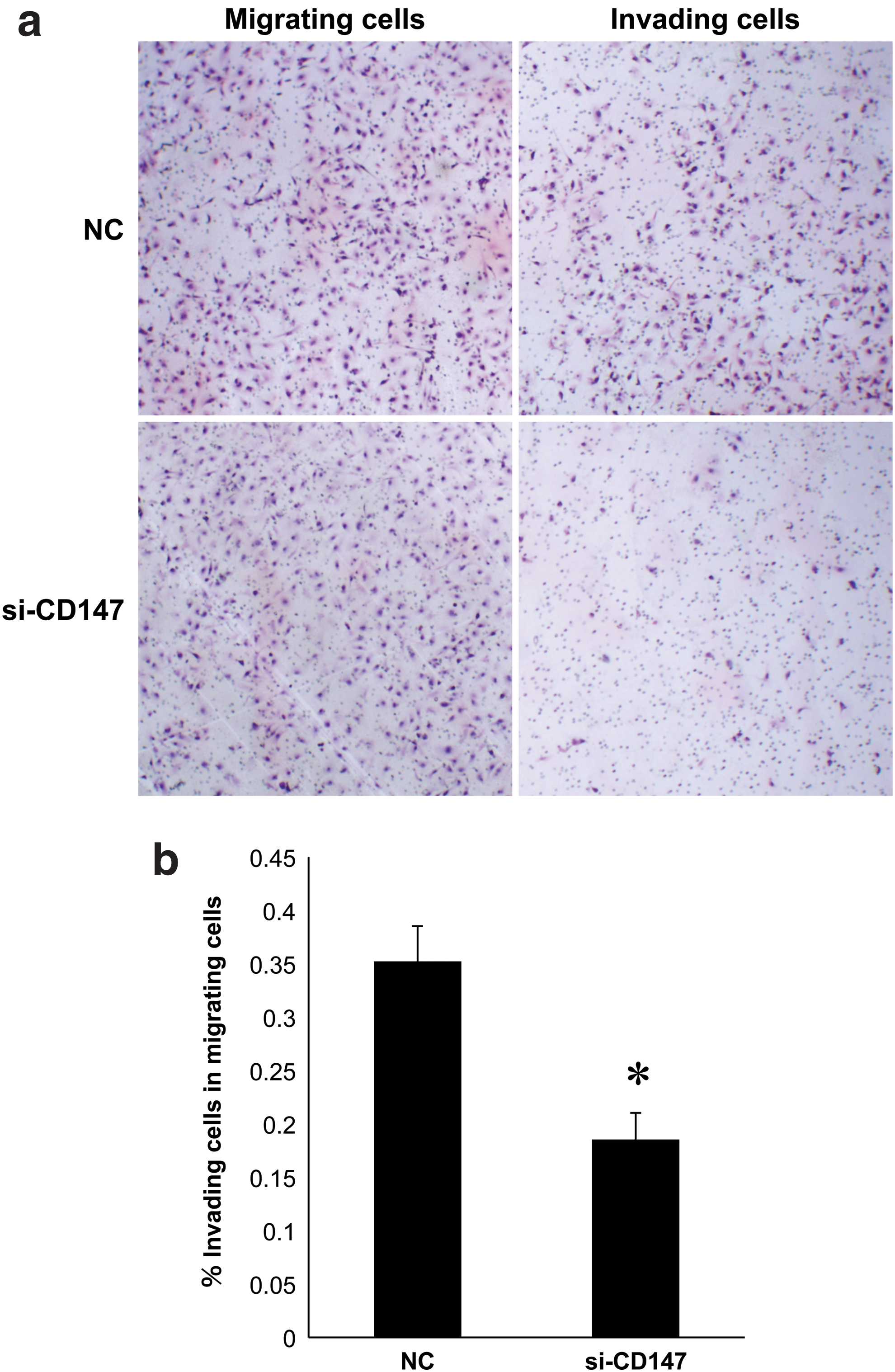
Microphotographs of migrating and invading FTC-133 cells
Discussion
CD147 is a cell surface glycoprotein that upregulates MMP, assisting cancer cell invasion during disease progression and hematogenous or lymphogenous metastasis (10). Several immunohistochemical studies have shown CD147 expression in human thyroid tissues. A report demonstrated that CD147 expression is increased in papillary thyroid carcinoma with poorly differentiated components or in undifferentitated carcinoma compared with well differentiated papillary thyroid carcinoma (24). Another study on 156 differentiated thyroid carcinomas indicated a link between CD147 expression levels and clinicopathological features including prognosis (25). In the present study, we demonstrated distinct CD147 immunoreactivity in FTC tissue sections that were in striking contrast to the weak immunoreactivity in normal thyroid tissue sections. However, since anti-CD147 antibodies generally recognize both NG-CD147 and G-CD147, it was not possible to identify the two forms in immunohistochemically stained sections. Similarly, since the anti-MMP antibodies recognize both P-MMP and C-MMP, it is difficult to distinguish them on immunostained sections. Moreover, immunohistochemistry is insufficient and inadequate for quantitative analysis. To address this issue, we employed immunoblot and densitometric techniques for the detection of CD147 and MMPs in normal thyroid, FA, and FTC tissues. It was noteworthy that both the NG-CD147 and G-CD147 levels were significantly greater in the FTC group than the normal thyroid and FA groups, suggesting the involvement of CD147 in the malignant potential of FTC. Since we examined only eight FTC cases for detection of CD147, the number of cases was too small for statistical evaluation. Thus, it remains to be determined whether CD147 expression levels are higher in widely invasive follicular carcinoma than in minimally invasive follicular carcinoma.
Given that MMPs mediate effects of CD147 on invasiveness and metastasis of FTC, we made quantitative measurements of MMPs in normal thyroid, FA, and FTC tissues. The expression levels of MMP-3, MMP-7, and MMP-9 were significantly greater in the FTC group than in the normal thyroid group. These observations may reflect the role of CD147 in the malignant behavior of FTC. In particular, it is of interest that the T-CD147, T-MMP-3, and T-MMP-7 levels were significantly increased in the FTC group compared with the FA group. This suggests the possibility that these proteins might be relevant to malignant transformation and be candidate markers to distinguish FTC from FA. Moreover, our finding of the significantly increased levels of MMP-9 in the FA and FTC groups compared with the normal thyroid group suggests a common mechanism between FA and FTC related to MMP-9 functions. A previous immunohistochemical study showed significant increases in the MMP-2, MMP-7, and MMP-9 levels in FTC compared with FA (5). Although these findings seem to be in keeping with our results to some extent, it is impossible to identify a preferential form of each MMP by a morphological approach. On the other hand, it should be noted that the expression of MMPs can be regulated by factors other than CD147 such as inflammatory cytokines, growth factors, mechanical stimuli, and phagocytotic activities (4,7). To minimize the influence of these factors in the current study, we used thyroid tissues without evidence of inflammatory disorder including chronic thyroiditis.
EGF drives CD147 expression in a human breast cancer cell line (30) and stimulates the invasive activity of FTC-133 cells (31,32). EGF also activates MMP-2 and induces MMP-9 and MT1-MMP in FTC-133 cells (32). Several studies on carcinoma cells other than FTC cells have revealed cell signaling pathways that mediate CD147 induction (30,33). Based on evidence for the expression of EGFR and CD147 in FTC-133 cells, we sought to identify the EGF-driven, CD147-upregulating signal pathways in FTC cells, by comparing expression levels of NG-CD147, G-CD147, and T-CD147 in FTC-133 cells with or without pretreatment with cell signaling enzyme inhibitors. Consequently, wortmannin, PD98059, and SP600125 but not SB203580 significantly prevented the EGF-driven CD147 upregulation. These observations could indicate that EGF upregulates CD147 expression via the PI3K, ERK, and JNK pathways.
To determine whether MMPs are upregulated in response to CD147 expression, we compared expression levels of MMPs on immunoblots using FTC-133 cells with or without CD147 silencing. Although CD147 has been implicated in MMP induction rather than MMP activation (10), our study showed that CD147 silencing downregulated the precursor, cleaved and total forms of MMP-2 and MMP-7, and that in contrast it downregulated the cleaved and total but not precursor forms of MMP-3 and MMP-9. These observations suggest that CD147 is responsible for MMP upregulation in FTC cells, and that the effects of CD147 on induction and activation of MMP-2, MMP-3, MMP-7, and MMP-9 vary among these MMP isoforms. Moreover, there is no precedent for CD147 silencing-induced MMP-7 downregulation. Our findings that MMP-2 was not upregulated in human FTC tissues and that it was downregulated in si-CD147-treated FTC-133 cells raise the possibility that there is another factor modulating expression levels of MMP-2, which may be derived from the stromal cells in thyroid tissues. Given the involvement of CD147 in invasiveness of several carcinoma cells, it is hypothesized that CD147 may also participate in FTC invasion. To test this hypothesis, we investigated invasive activities of FTC-133 cells with or without CD147 silencing. As previously described in human thyroid tissue sections (23 –25), it was evident that FTC-133 cells were immunoreactive for CD147. Our finding that CD147 silencing reduced the invasive activities of FTC-133 cells suggests the involvement of CD147 in the mechanism of FTC invasion.
Collectively, we demonstrate for the first time the role of CD147 in FTC as the following results: (i) CD147, MMP-3, MMP-7, and MMP-9 are upregulated in human FTC tissues; (ii) EGF-driven CD147 upregulation in cultured FTC-133 cells occurs via the PI3K, ERK, and JNK pathways; (iii) expression of MMP-2, MMP-3, MMP-7, and MMP-9 in cultured FTC-133 cells is CD147-dependent; and (iv) the invasiveness of cultured FTC-133 cells is CD147-dependent. The present results suggest a pivotal role for CD147 in the malignant behavior of FTC cells, in a paracrine or autocrine manner. Based on our results, it is likely that novel diagnostic approaches, including enzyme-linked immunosorbent assay of serum CD147 levels, immunoblot analysis of CD147 protein levels in frozen tissues, and quantitative reverse transcription-polymerase chain reaction analysis of CD147 mRNA levels on paraffin-embedded sections would be helpful in predicting the prognosis of FTC and distinguishing FTC from FA, although to date the differential diagnosis currently depends on the detection of morphological features such as capsular or vascular invasion and lymph node or distant metastasis. Since we did not perform a carcinoma-stroma coculture in this study, it remains to be determined whether FTC cells interact with stromal cells via the CD147-related mechanisms. Better understanding of the CD147-related nature of FTC and carcinoma-stroma interaction will contribute to both the improvement of diagnostic accuracy and the development of therapeutic strategies for this malignant neoplasm.
Footnotes
Acknowledgments
The authors wish to thank S. Iwasaki, F. Muramatsu, N. Sakayori, H. Takeiri, M. Karita, Y. Kato, and T. Yamamoto for technical assistance.
Disclosure Statement
The authors declare that no competing financial interests exist.