
Abstract
Background:
Angiogenesis plays an essential role in embryonic and tumoral developments. Vascular endothelial growth factor (VEGF), one of the best known proangiogenic factors, is increased in thyroid cancers, especially in papillary carcinomas (PC). However, other regulating mechanisms refine VEGF-induced cellular changes, such as the Notch family of ligands and receptors. Their role has not yet been investigated in the thyroid. The purpose of our study was to analyze the expression of Notch1, Notch4, and Delta-like 4 (DLL4) in benign and malignant thyroid lesions.
Methods:
The expression of Notch1, Notch4, and DLL4 was analyzed by immunohistochemistry, quantitative reverse transcriptase–polymerase chain reaction (qRT-PCR), and Western-blot in normal thyroids (NTs), hyperplasic thyroids from patients with Graves' disease (GD), microcarcinomas, PC, and follicular carcinomas.
Results:
The immunohistochemical expression of Notch1, Notch4, and DLL4 was highly variable in thyrocytes from NTs and GD. In contrast, the staining in tumors was homogeneous and often intense. The increased expression of Notch1, Notch4, and DLL4 in carcinomas compared with the neighboring normal tissue was confirmed by qRT-PCR and Western-blot. However, only capillary endothelial cells from GD samples were positive for DLL4, the expression being restricted to large vessels in carcinomas and NTs.
Conclusions:
The detection of Notch1, Notch4, and DLL4 in thyrocytes and their regulation in various pathologies suggest that this pathway may play a role in thyroid carcinogenesis and angiogenesis.
Introduction
In humans, four Notch receptors (1 –4) and five ligands (delta-like 1, 3, 4 and Jagged-1 and -2) have already been identified. The Notch receptor is a transmembrane protein consisting of an extracellular, a transmembrane, and an intracellular domain. The activation of the Notch receptors occurs through contacts between the membrane-bound ligands and the receptor extracellular domain, each located on two neighboring cells (4). The translocation of the receptor intracellular domain into the nucleus triggers the transcription of genes regulating apoptosis, differentiation, cell-cycle progression, angiogenesis, and cellular metabolism (5,7,8). This cell-to-cell signaling pathway is highly conserved during evolution in vertebrates and invertebrates (9). First described in the nervous system, the Notch pathway also plays an important role during angiogenesis, particularly when activated by the Delta-like 4 (DLL4) ligand.
DLL4 is the specific ligand for Notch1 and Notch4 (5,10,11). In mice, the haplo-insufficiency of DLL4 results in embryonic death due to vascular defects, malformations, and arterio-venous shunts (12,13). Of note, similar features were previously described only in VEGF-A haplo-insufficient mice, which underlines the critical importance of DLL4 in angiogenesis. Interestingly, DLL4 is induced by VEGF and acts downstream to VEGF, refining the endothelial response to the growth factor. During angiogenesis, DLL4 is mainly expressed in tip cells, whereas Notch1 is expressed in stalk cells. VEGF induces DLL4, which then binds to the Notch1 receptor expressed in adjacent endothelial cells. Notch1 activation decreases VEGFR expression, inducing the transformation of endothelial cells from the tip to the stalk type, thereby permitting the differentiation of new vessels. DLL4 prevents excessive vessel proliferation by blocking branching processes and by triggering the differentiation of endothelial cells (14). Conversely, the inhibition of the DLL4/Notch pathway increases the vascular density, but this excessive sprouting is nonproductive, thereby leading to poorly functional vascular networks (15,16). In tumors, although the over-expression of DLL4 does not correlate with tumor growth (16,17), DLL4 is able to block the growth of anti-VEGF resistant tumors (15).
The thyroid gland is a highly vascularized organ, and angiogenesis plays a critical role, not only in several pathologies including carcinomas, but also during goiter development that may rather be considered as a physiological process of adaptation to everlasting changing environmental conditions, such as the iodide supply (18 –21).
In the thyroid gland, only Notch 1, 2, and 3 have been reported in a study that suggested a role for the Notch pathway in thyroid cell differentiation (22). However, DLL4 and Notch4 have never been studied in the thyroid, even though they have been clearly identified as crucially involved in angiogenesis regulation. The purpose of the current study was, therefore, to analyze the expression of important components of the Notch pathway, including DLL4, Notch1, and Notch4, in correlation with angiogenic processes occurring in normal thyroids (NTs), hyperplasic thyroids from patient with colloid nodules and with Graves' disease (GD), microcarcinomas (MCA), classical papillary carcinomas (PC), follicular variant of PC, and follicular carcinomas (FC). We analyzed not only the level of expression but also the distribution of the proteins to differentiate between epithelial and endothelial expression.
Materials and Methods
Human thyroid tissues
Western blots (WB) and quantitative reverse transcriptase–polymerase chain reaction (qRT-PCR) analyses were carried out by using snap-frozen materials that were prospectively and anonymously collected without a link to the living subjects, from 10 PC (1 classical type, 8 follicular variants, and 1 mixed type), from normal surrounding tissues, near (NN) and far (NF) from the tumor and from colloid nodules in multinodular goiters.
The immunohistochemical analysis was performed from 5 NT tissues, 10 hyperplastic thyroids from patients with GD, 8 micro-carcinomas, 20 PC (10 classical types and 10 follicular variants), and 5 FC that had been retrospectively selected from the files of the UZ Brussels' pathology department. These specimens were formalin fixed and paraffin embedded. Tumor specimens were selected only when normal peritumoral tissues were also present.
Immunohistochemistry
Four-micrometers sections were deparaffinized, rinsed with phosphate-buffered saline (PBS), and incubated for 30 minutes in 3% hydrogen peroxide in methanol. The slides were rinsed with PBS, and incubated for 30 minutes in PBS-bovine serum albumin containing normal goat serum (Life technologies) at a dilution of 1/50. The first antibody was then applied (see Table 1 for concentrations, incubation times, and pretreatments). After two washes in PBS, the antibody binding was detected with a second antibody conjugated to a peroxidase-labeled polymer (Envision Detection; DAKO) for 30 minutes at room temperature. After washing, peroxidase activity was revealed with the AEC substrate (3-amino-9-ethylcarbazole; DAKO). Sections were counterstained with Mayer's hematoxylin, rinsed, and mounted in Faramount (DAKO). Several negative controls were performed, for instance by omitting the first antibody or by using a commercially available isotype control antibody (ab18457, mouse IgG2b kappa monoclonal antibody directed against trinitrophenol and KLH). Positive controls for DLL4 and Notch4 were performed in tubuli of kidney tissue and in colon carcinomas for Notch1. One thousand cells from each sample were evaluated for the staining of DLL4, Notch1, and Notch4 and classified as negative, weakly, and highly positive.
Santa Cruz, Heidelberg, Germany.
Abcam, Cambridge, MA.
Western blottings
Thyroid homogenates from frozen materials were suspended in Laemmli buffer (50 mmol/L Tris-HCl, pH 6.8, 2% sodium dodecyl sulfate, and 10% glycerol) containing a protease inhibitor cocktail (Sigma). Protein concentration was determined by using a BCA protein assay kit (Pierce). Proteins (30 μg/lane) were heated at 95°C for 5 minutes in the loading buffer (Laemmli buffer containing 100 mmol/L dithiothreitol and 0.1% bromophenol blue), proteins were separated by 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto a nitrocellulose membrane (Hybond ECL; Amersham Biosciences). Membranes were blocked for 1 hour at room temperature in PBS (pH 7.4), 5% nonfat dry milk, 0.1% tween, and incubated overnight at 4°C with the previously described polyclonal antibody raised against DLL4 (1/500). Membranes were washed with PBS/Tween 0.1%, incubated for 1 hour at room temperature with EnVision (1/200; DakoCytomation) peroxidase-labeled secondary antibody, washed, and visualized with enhanced chemiluminescence (SuperSignal West Femto; Pierce) on CL-Xposure TM films (Pierce). The same membranes were also incubated with an anti-β-actin antibody (1/2000; Sigma). The WB were scanned and quantified by pixel densitometry with the NIH Scion Image Analysis Software (National Institutes of Health). Values were normalized by reporting the signal intensity to β-actin expression.
RNA purification and reverse transcription
Frozen materials were homogenized and suspended in TriPure isolation reagent (Roche Diagnostics GmbH). Total RNA was purified according to the manufacturer's protocol and resuspended in 6 μL of H2O. Reverse transcription was performed by incubating RNA with 200U Moloney murine leukemia virus-reverse transcriptase (M-MLV-RT; Invitrogen) in the recommended buffer containing 4 U of RNasin (Promega), 0.5 mmol/L dNTP (Promega), 2 μmol/L oligodT (Promega), and 10 mmol/L dithiothreitol (20 μL final volume) overnight at 42°C. H2O (80 μL) was then added, and products were used for PCR amplification analyses.
Quantitative PCR
cDNAs (2 μL) were mixed with 500 nmol/L of each of the selected primers (Table 2) and SYBR Green reaction mix (Quanta Biosciences) in a final volume of 25 μL. Reactions were performed by using an iCycler apparatus (BioRad) as follows: 95°C for 1 minute, followed by 40 cycles of 95°C for 15 seconds, annealing temperature for 45 seconds (Table 2), and 81°C for 15 seconds. Amplification levels were normalized to those of β-actin.
Statistical analysis
All data were expressed as mean±standard error of the mean. Statistical analyses were performed by using ANOVA followed by Tukey-Kramer Multiple comparison test, or by unpaired-t test (GraphPad Instat). p<0.05 was considered statistically significant.
Results
Elevated expression of DLL4 in thyroid carcinomas and GD
To investigate the expression of DLL4, a qRT-PCR analysis was carried out to detect DLL4 mRNA expression (Fig. 1A), and WB for DLL4 protein expression (Fig. 1B, C). Since several factors may influence DLL4 expression, NT tissue located at distance from the tumor served as a control, and a paired-sample analysis was performed. Samples from malignant, normal near the tumor (NN), and normal far away from the tumor (NF) were available in eight patients, as well as samples from colloid nodules, close to the nodule (NN nod), and far from the nodule (NF nod) were available in five patients. There was no significant difference in the expression of DLL4 mRNA between normal tissues (NN, NF, NN nod, and NF nod), and between normal tissues and colloid nodules (nodule). By contrast, DLL4 mRNA expression in thyroid PC was higher than in NT tissues and colloid nodules (p<0.05).

DLL4 mRNA and protein expression in multinodular goiters and in papillary carcinomas. Relative expression of DLL4 mRNA adjusted to the β-actin signal, expressed as mean±standard error of the mean (SEM) (n=8) on specimens prospectively harvested
DLL4 protein was detected as a band of 74 kDa in NTs (Fig. 1B, lower panel). DLL4 protein expression was significantly higher in PC compared with normal tissues (NN, NF, NN nod, and NF nod). It was lower in the NT tissue close (NN) to the tumor compared with that distant (NF) from the tumor. There was no significant change in the expression of DLL4 protein between normal tissues (NN nod, NF nod) and colloid nodules (Fig. 1B).
To localize the protein expression of DLL4, an immunohistochemical analysis was carried out in normal, hyperplastic (GD), and cancerous thyroid tissues (MCA, PC, and FC) (Fig. 2A). DLL4 expression was detected in benign and malignant thyroid tissues. Immunohistochemistry showed a heterogenic cytoplasmic positivity in normal and hyperplastic thyrocytes. Areas with strong staining were observed around negative zones. In all tumors, the cytoplasmic staining for DLL4 was strong and diffuse.

Immunohistochemical detection of DLL4, Notch1, and Notch4 in normal, Graves' disease, and thyroid carcinomas.
Due to the heterogeneity of the staining, we counted the number of negative, weakly positive, and strongly positive cells. The cell counting revealed a significant higher number of DLL4-negative cells in samples from normal tissue and GD, compared with PC (data not shown). Papillary cancers contained the higher number of strongly DLL4-positive cells (Fig. 2B). This number was fourfold increased, as compared with the normal peritumoral tissue, and nine fold increased compared with NTs. The number of DLL4-positive cells was also higher in FC and in MCA than in normal, although less than in PC. The number of highly positive cells was also increased in hyperplastic tissues, but at a level that was half of that observed in PC.
The morphological analysis of DLL4 staining showed that in addition to thyrocytes, DLL-4 was also detected in vessels. Endothelial staining for DLL4 was restricted to large vessels in NTs and in cancers (data not shown), whereas small vessels were negative in these samples (Fig. 2A). By contrast, small and large vessels were positive in GD (Fig. 2A).
Thus, the present data indicate that DLL4 expression is increased in thyrocytes from carcinomas, as well as in GD, although to a smaller extent. In addition, DLL4 is also detected in microvessels, but only in GD, thereby suggesting (as expected) a differential regulation of angiogenesis in hyperplasia and thyroid carcinomas.
Elevated expression of Notch 1 and 4 receptors in thyroid carcinomas
The expression Notch1 and Notch4 mRNAs was analyzed by qRT-PCR (Fig. 3). Thyroid PC had the higher Notch1 expression compared with NT tissues (p<0.05) and colloid nodules (p<0.05) (Fig. 3A). There was no significant change in the expression of Notch1 between normal perinodular tissues (NF nod, NN nod) and colloid nodules. However, NN expressed lower levels of Notch1 mRNA expression than NF (p<0.05). Similar observations were made for Notch4 mRNA expression (p<0.05), although here, NN showed the same Notch4 mRNA expression as NF (Fig. 3B).
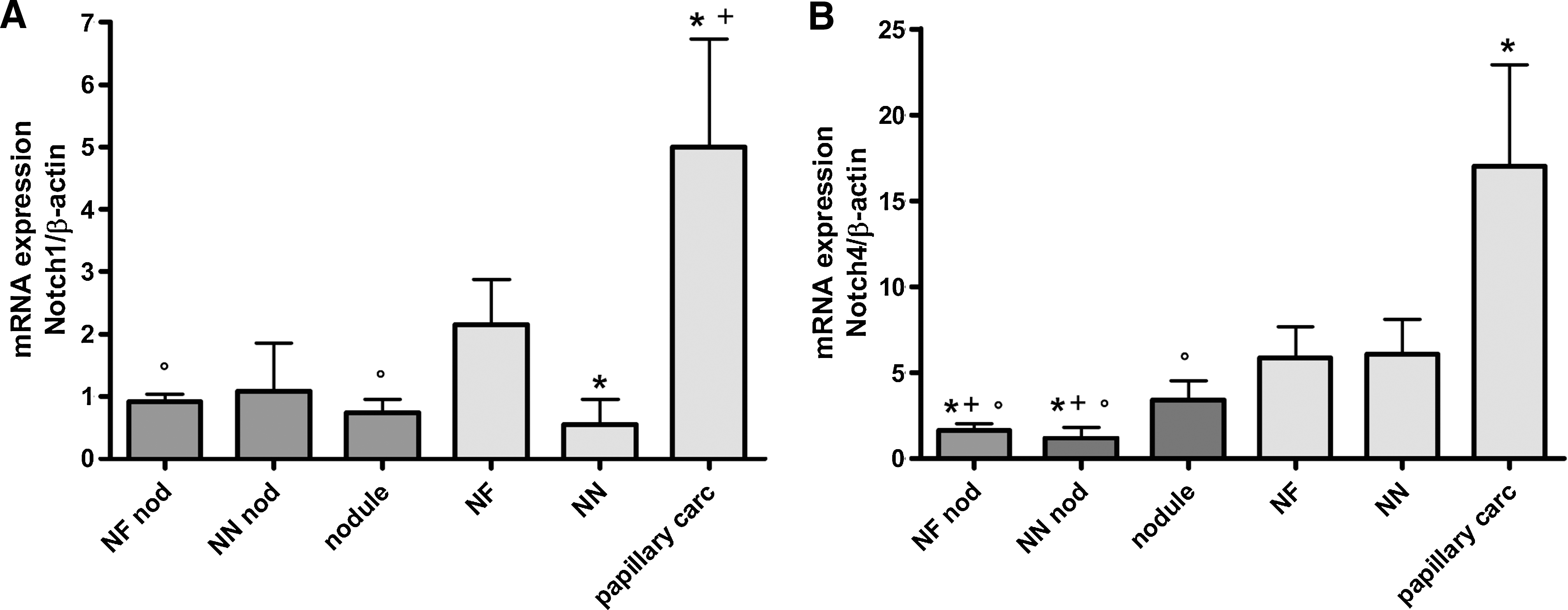
Notch1 and Notch4 mRNA expression in multinodular goiters and in papillary carcinomas. The relative expression of Notch1
Immunohistochemical stainings for Notch1 and Notch4 were performed to localize the protein expression in normal, hyperplastic, and cancerous thyroid tissues (Fig. 2A). Notch1 and Notch4 expression was detected in all samples of benign and malignant thyroid tissues. Immunohistochemistry for Notch1 showed a heterogenic cytoplasmic positivity in normal and hyperplastic thyrocytes. Areas of strong immunoreactivity were separated by negative zones. In malignant tissues, the cytoplasmic staining was diffuse and strong through all the tumors. In addition, some nuclear staining was observed in all the tissues. Cell counting showed that the number of Notch1-negative cells was significantly higher in normal and GD tissues compared with PC (data not shown). By contrast, all types of PC contained a higher number of strongly Notch1-positive cells compared with normal and hyperplastic tissues (p<0.05, Fig. 2B), whereas the expression of Notch1 in FC remained in the same range than in benign tissues. Interestingly, in GD thyroids, the number of highly positive cells was decreased compared with NTs (p<0.05). A weak endothelial staining was observed in all vessels, including microvessels, in all thyroids studied. Nevertheless, the endothelial staining remained weaker than that of thyrocytes.
The expression of Notch4 was the highest amongst the three analyzed proteins in normal tissues. In PC, the staining was diffuse and strong in the cytoplasm of thyrocytes. In addition, some stained nuclei were observed in malignant as well as in benign tissues. The expression of Notch4 was stronger in FC than in normal and hyperplastic tissues, but weaker than in PC. As for Notch1, the number of Notch4-negative cells was significantly higher in normal and GD tissues than in PC (data not shown). By contrast, PC exhibited a higher number of strongly Notch4-positive cells compared with normal tissues, although this increase was lower than that observed for Notch1 and DLL4 (p<0.05). The number of highly positive cells for Notch4 was similar in hyperplastic tissues than in NTs (Fig. 2B). Notch4 was expressed in all endothelial cells, but less than in thyrocytes, and without notable differences between pathologies.
Discussion
Historically, the Notch system was first described in neurogenesis (23) and later, in angiogenesis (24). It is now accepted that receptors and ligands of the Notch family are widely expressed in normal, reactive, and neoplastic tissues, in endothelial and nonendothelial structures (7,25 –27). In this study, we report for the first time the presence and the regulation of the Notch receptors, Notch1 and Notch4, as well as of their ligand DLL4 in the thyroid. Our data demonstrate that the three components are overexpressed in thyrocytes of thyroid cancers compared with normal and hyperplastic thyroid tissues, but also in hyperplastic tissues compared with normal tissues. The detection of DLL4 and Notch receptors in both endothelial cells and thyrocytes suggests a role not only in thyroid cell signaling, but also in neoplastic and non neoplastic angiogenesis.
The role of Notch has not yet been fully elucidated in the thyroid. Ferretti et al. reported the presence of Notch1, -2, and -3 in normal thyrocytes (22) and suggested a role for Notch1 in thyroid cell differentiation. The expression of Notch1 and -3 mRNAs was decreased in anaplastic carcinomas, but similar to that of the control in PC and FC. In the current study, Notch1 is also expressed in the NT, but in contrast with Ferretti et al., its expression is increased in PC. This could be due to the fact that different inclusion criteria were used in each study, notwithstanding the variability usually observed among cancers. We also took care while analyzing well-differentiated cancers and focused our research on the link between angiogenesis and DLL4 expression. Since we found an increased expression of DLL4 and of its receptors in well-differentiated carcinomas, it is possible that this pathway influences oncogenic processes in the thyroid.
As recently reported, the Delta-Notch pathway may play a role in tumor angiogenesis (6). As suggested by our observations in GD, this could also be the case in non neoplastic angiogenesis. The differences in DDL4 expression, especially in endothelial cells of small capillaries, may infer different regulations when non-neoplastic and neoplastic growths are considered. The expression of DLL4 is low in the normal adult vasculature, but strongly upregulated in tumor vasculature as reported for instance in renal, bladder, and breast carcinomas (24,26,27). Of note, in our study, DLL4 expression was high in epithelial cells, but in fact lacking in small vessels of neoplastic tissues in deep contrast with hyperplastic benign tissues. Although the exact function of DLL4 is not yet fully understood, evidence suggests that it may act downstream to VEGF through negative feedback to avoid uncontrolled endothelial proliferation, while maintaining the integrity of newly formed blood vessels (5,15 –17). Thus, our observation that DLL4 is absent in small vessels of neoplastic thyroids along with a high VEGF expression (not shown) is consistent with a loss of finely tuned angiogenesis in malignant lesions. Conversely, in hyperplastic benign thyroids (GD), although the vasculature is also growing up, but not in an anarchic way, DLL4 expression is present in microvessels, thus suggesting that now, the growing vasculature is preserved in a “healthier” status (Fig. 4).
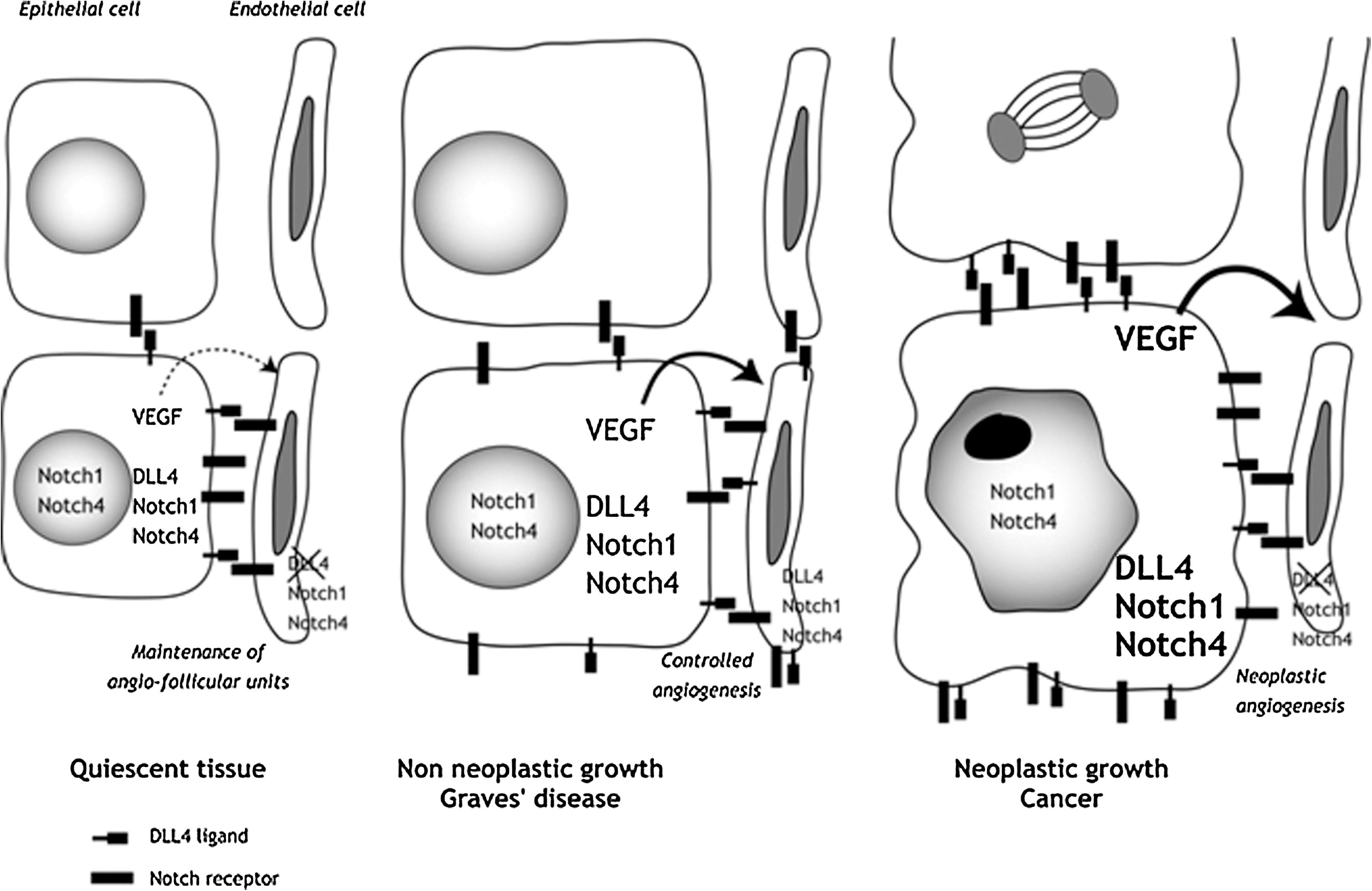
Schematic and hypothetical representation of the Notch-DLL4 signaling pathway in normal (quiescent), non neoplastic but hyperplastic (Graves' disease), and neoplastic tissues (cancer). See the text for comments.
Having in mind that the thyroid gland is composed of angio-folicular units with tight relations between epithelial and endothelial cells based on several growth and vasoactive factors, the discovery of the Delta-Notch pathway is not really a surprise (18 –21). The thyroid microcirculation is indeed thoroughly involved in the maintenance of proper thyrocyte function (18 –20). In NTs, the quiescence and fenestration of angio-follicular unit capillary networks is ensured by the secretion of small amounts of VEGF by follicular cells. Even though DLL4 is not expressed, Notch1 and 4 are detected in endothelial cells. Hence, it is possible that direct physical links between epithelial (that express DLL4) and endothelial cells (that express Notch1 and Notch4) occur through the DLL4/Notch system, thereby contributing to the maintenance of angio-follicular units. In conditions of non neoplastic growth and angiogenesis as in GD, coherent cross talk between cells from the two tissue compartments lead to controlled angiogenesis and again safeguard the integrity of angio-follicular units. Here, enhanced VEGF secretion by follicular cells is associated with and perhaps controlled by DLL4 expressed in endothelial cells, inducing proper differentiation and modeling of the vascular network. Conversely, in carcinomas, when these cross talks are disrupted because DLL4 is no more adequately expressed in endothelial cells of capillaries, high levels of “uncontrolled” VEGF may lead to perverted and uncontrolled angiogenesis, the integrity of angio-follicular units being then irrevocably disrupted (see Fig. 4).
Footnotes
Disclosure Statement
The authors declare that no competing financial interests exist.