
Abstract
Background:
The type 2 deiodinase gene (DIO2) encodes a deiodinase that converts the thyroid prohormone, thyroxine, to the biologically active triiodothyronine. Thyroid hormones regulate energy balance and may also influence glucose metabolism. Therefore, we hypothesized that variations in DIO2 could contribute to obesity or type 2 diabetes mellitus (T2DM) in Pima Indians.
Methods:
Sequencing of the DIO2 gene in DNA from 83 Pima Indians identified 12 single-nucleotide polymorphisms (SNPs). Several of these SNPs were in perfect genotypic concordance among the 83 samples that were sequenced, and all 12 could be divided into five linkage disequilibrium groups. One representative SNP from each group (Thr92Ala, rs225011, rs225015, rs6574549, and a rare 5′ flanking SNP) was selected for further genotyping for association analyses. In this study, the five selected variants in DIO2, as described above, were genotyped in three groups of Pima Indians: (i) a case (n=150)/control (n=150) group for early-onset T2DM (onset age <25 years); (ii) a case (n=362)/control (n=127) group for obesity; (iii) a large (n=1,311, cases n=810/controls n=501) family-based group, of which 256 nondiabetic subjects had undergone detailed metabolic phenotyping.
Results:
The Thr92Ala variant common in Pima Indians, rs225011, and rs225015 were modestly associated with early-onset T2DM (p=0.01–0.04) in the case–control study, but were not associated with obesity in the obesity case–control study, nor associated with T2DM (at any age) or body–mass index (BMI; as a quantitative trait) in the family-based analysis. Thr92Ala, rs225011, rs225015, and rs6574549 were also nominally associated with hepatic glucose output (p=0.02). rs6574549 was associated with fasting insulin (p=0.02), insulin action (p=0.04), and energy expenditure (p=0.02). None of these nominal associations remained statistically significant after corrections for multiple testing.
Conclusions:
We propose that variation in DIO2 may have a subtle role in altering metabolic processes that lead to early-onset T2DM, but this gene does not have a large impact on T2DM at older ages, nor does DIO2 influence BMI in the Pima Indian population.
Introduction
In addition, glucose homeostasis can be affected by thyroid status (8,9). Experimental or spontaneous forms of thyroid dysfunction are associated with impaired glucose tolerance (10). Increase in both hepatic insulin resistance (11,12) and insulin-mediated glucose uptake is observed in hyperthyroidism as compared with euthyroid conditions (11). Thyroid hormones influence insulin action in skeletal muscle and adipose tissue in part by upregulating the expression of the muscle/fat-specific glucose transporter 4 (13,14). In contrast, studies in hypothyroid conditions show a decrease in both hepatic insulin resistance (12) and insulin-mediated glucose uptake in muscles (10,12). In humans, subclinical hypothyroidism is associated with hyperinsulinemia (15).
Deiodinases are selenoenzymes that convert the prohormone thyroxine (T4) into active triiodothyronine (T3) or inactive reverse triiodothyronine (rT3) (16). Type 1 deiodinase (D1), which is predominantly expressed in liver, kidney, and thyroid gland, can function as an outer- (T4 to T3) or inner-ring (T4 to rT3) deiodinase (16,17). Type 2 deiodinase (D2), which is an outer-ring deiodinase (T4 to T3), is found in thyroid gland, cardiac and skeletal muscle, brown adipose tissue, placenta, pituitary, central nervous system (CNS), and at low levels in kidney and pancreas (16 –18). Type 3 deiodinase, an inner-ring deiodinase, converts active T3 to inactive rT3 and is typically expressed in liver, skin, placenta, CNS, and fetal tissues (16,17). Although it was thought that circulating T3 was mostly generated by D1, and that D2 was more important for intracellular T3 generation (19), it has been recently suggested that D2 is the main source of circulating levels of T3 in euthyroid subjects (20). Differences in D2 activity might affect not only circulating and peripheral T3 concentrations but also intracellular T3 availability to interact with thyroid hormone nuclear receptors.
A nonsynonymous amino acid substitution, Thr92Ala, in D2 has previously been reported to be associated with insulin resistance in obese Caucasian women (21). This study found that the frequency of the “risk” (Ala) allele was 0.35 in Caucasians, but further noted that this risk allele was much more common (AF=0.75) among Pima Indians. This same study provided evidence of an interaction between the Thr92Ala variant in D2 and a Trp64Arg variant in the β-3-adrenergic receptor (21). Nominal associations were reported between the Thr92Ala variant and the serum insulin curve during oral glucose tolerance test (OGTT) in a large Danish cohort (22). The Thr92Ala was also reported to be associated with insulin resistance in patients with type 2 diabetes in a Brazilian cohort (23). A recent meta-analysis in ∼11,000 subjects confirmed the association of Thr92Ala with increased risk for type 2 diabetes (24). However, there are also studies suggesting that the associations with insulin resistance and/or type 2 diabetes are inconsistent in some populations (25 –27). In the current study, we analyzed associations of other single-nucleotide polymorphisms (SNPs) in addition to the common Thr92Ala variant of the type 2 deiodinase (DIO2) gene for association with type 2 diabetes and obesity in the Pima Indian population.
Materials and Methods
Subjects
All subjects are participants of our ongoing longitudinal study of the etiology of type 2 diabetes among the Gila River Indian Community in Arizona (28). DIO2 was sequenced in DNA from 83 non–first-degree-related morbidly obese Pima Indian subjects (mean body–mass index [BMI]=60.0 kg/m2 ranging from 50.5 to 79.6 kg/m2). Genotyping was done in three groups of subjects: (i) an independent case–control group of full-heritage, non–first-degree-related Pima Indians for analysis of early-onset diabetes (29); (ii) an independent case–control group for analysis of obesity (30); (iii) a family-based association study in which the subjects are participants of the ongoing longitudinal study of the etiology of type 2 diabetes among the Gila River Indian Community in Arizona (28,31). About 5% of Pima Indians aged 15–24 years have earlier onset of type 2 diabetes (29,32). The case–control group for early-onset type 2 diabetes consisted of 150 diabetic subjects with an age of onset <25 years and 150 nondiabetic subjects who were determined to be nondiabetic at their last examination, which was after the age of 45 years. The case–control group for obesity consisted of 362 severely obese subjects, as defined by a maximum BMI >45 kg/m2, and 127 control nondiabetic, nonobese subjects selected for a BMI <30 kg/m2 at an age >35 years and not having diabetes. All of these subjects were full-blooded Pima Indians and none were first-degree relatives. The large family-based group consisted of 1,311 subjects from 322 nuclear families, of which, 810 subjects had type 2 diabetes as determined by an OGTT that is interpreted according to the criteria of the World Health Organization (33). Among the family-based group, 256 nondiabetic, full-heritage Pima subjects had undergone metabolic characterization as inpatients in our Clinical Research Center. All subjects provided written informed consent prior to participation. All studies were approved by the Tribal Council of the Gila River Indian Community and the Institutional Review Board of the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK).
Metabolic characterization of nondiabetic subjects
Volunteers were admitted to the Clinical Research Unit where they consumed a weight-maintaining diet (containing 50% of calories as carbohydrates, 30% as fat, and 20% as protein) for 2–3 days prior to clinical testing. Body composition was measured by dual energy X-ray absorptiometry (DXA) using a total body scanner (DPX-L; Lunar Radiation, Madison, Wisconsin) as described previously (34). The OGTT was performed, and blood for plasma glucose and insulin measurements was drawn before ingesting glucose, and at 30, 60, 120, and 180 minutes thereafter. Subjects underwent a two-step (low and high insulin dose) euglycemic-hyperinsulinemic clamp to assess insulin-mediated glucose disposal as described previously (35). The rate of basal endogenous glucose output was measured before insulin infusion and during the last 40 minutes of the low-dose insulin infusion using tritiated glucose and calculated from the Steele equation (36). All measurements derived from the glucose clamp were normalized to estimated metabolic body size (EMBS, which equals fat-free mass+17.7 kg). The measurement of energy expenditure in the respiratory chamber has previously been described (37). The rate of energy expenditure was measured continuously, calculated for each 15-minute interval, and then averaged for a 24-hour interval (24-EE). Sleeping metabolic rate (SMR) was defined as the average energy expenditure of all 15-minute periods between 11:30
Identification and genotyping of polymorphisms
DIO2 was sequenced in genomic DNA from 83 Pima Indians. All four exons, the exon-intron junctions, the 5′ and 3′ untranslated regions, and ∼2.0 kb of the upstream (putative promoter) region were sequenced using Big Dye Terminator (Applied Biosystems), and the samples were analyzed on an automated capillary sequencer (model 3730; Applied Biosystems). Variants were genotyped by allelic discrimination using TaqMan Assays on Demand (Applied Biosystems) for all the SNPs except rs6574549, which was genotyped by SNPplex following the manufacturer's protocol (Applied Biosystems).
Statistical analysis
Statistical analyses were performed using the procedures of the Statistical Analysis System software (SAS Institute). The odds ratio was used to determine the strength of the association between the prevalence of the at-risk genotype and affection status. Associations were calculated under three different models: one assuming that the common allele is dominant to the less common allele, one assuming that it is recessive, and one assuming an additive relation between alleles and prevalence of disease. For continuous variables, the general estimating equation procedure was used to adjust for covariates that account for the correlation among family members (i.e., siblings). In the case–control study for early-onset type 2 diabetes, the association was assessed by logistic regression and controlled for sex and heritage. p-Values of <0.05 were considered significant. These nominal p-values were adjusted using the multiple-testing correction Sidak formula. We conducted the multiple comparisons correction assuming 5 SNPs and 5 trait domains (diabetes, BMI/fat mass, insulin resistance, hepatic glucose output, and SMR) for a correction factor of 25 total comparisons. We assumed 5 effective traits rather than 10 because many of these metabolic traits are highly correlated.
For the early-onset diabetes case–control study, power is 52% to detect an odds ratio of 1.50 at p<0.05 assuming a minor allele frequency (MAF)=0.2. For the obesity case–control study, power is 62% to detect an OR=1.50 at p<0.05 assuming an MAF=0.2. For the metabolic traits (n=256), power is 36% to detect an allele explaining 1% of variance at p<0.05. For the BMI linkage family-based study, we typically analyzed siblings only (n=1,085), rather than the full cohort of n=1311. With this assumption, power is 91% to detect an allele explaining 1% of the variance at p<0.05.
D′ was calculated as a measure of allelic association and r 2 as a measure of concordance. The analysis of interaction between Thr92Ala in DIO2 and Trp64Arg in β-3-adrenergic receptor (ADRB3) was performed as described previously (21).
Results
Sequencing of the DIO2 gene in DNA from 83 Pima Indians identified 12 SNPs (Fig. 1). These SNPs included a novel rare SNP (G/A) in the 5′ flanking region (−2,035 bp upstream of the translation start site), four intronic SNPs (rs225010, rs225011, rs225012, and rs225013), one coding SNP (rs225014 predicting Thr92Ala), and 6 SNPs in the 3′ untranslated region (rs6574549, a novel T/C SNP, rs225015, rs225016, rs225017, and rs225018). Several of these SNPs were in perfect genotypic concordance among the 83 samples that were sequenced, and all 12 could be divided into five linkage disequilibrium groups (defined as r 2=1 and D′=1; Fig. 1, SNPs grouped A–E). One representative SNP from each group (Thr92Ala, rs225011, rs225015, 6574549, and the rare 5′ flanking SNP) was selected for further genotyping for association analyses.
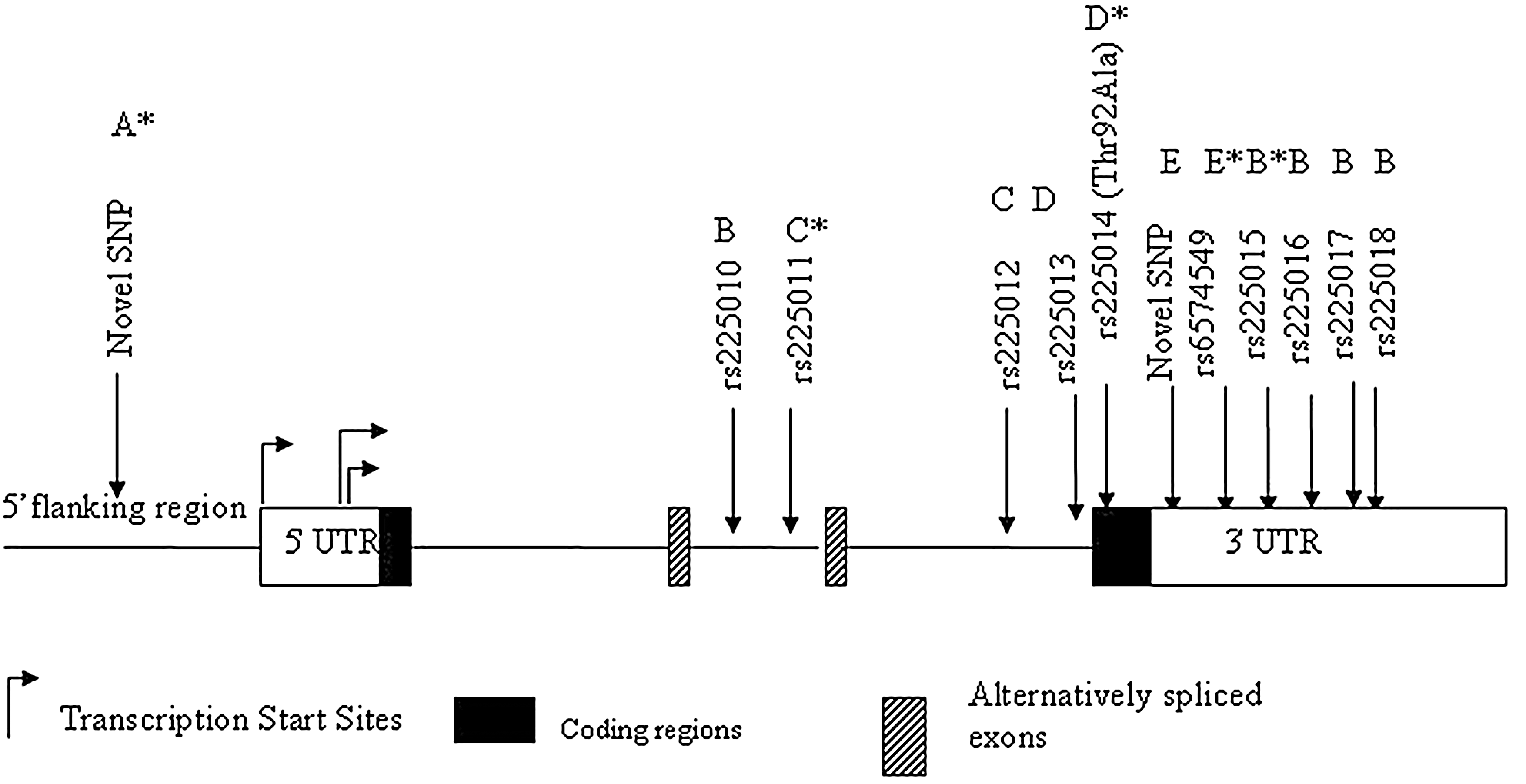
Gene structure of DIO2 and SNPs identified in Pima Indians. (A–E) SNPs grouped by perfect linkage disequilibrium (D'=1; r 2=1) among the 83 samples that were sequenced; *representative SNP from each group selected for additional genotyping. Locations of SNPs are not shown to scale. SNP, single-nucleotide polymorphism.
Genotyping of the five representative SNPs in the early-onset type 2 diabetes case–control subjects showed that three of the representative SNPs were associated with early-onset type 2 diabetes (Table 1). These included the Thr92Ala variant (p=0.01), as well as rs225011 and rs225015 (p=0.04 and 0.03, respectively). However, after corrections for multiple testing, none of the associations remained statistically significant.
The major allele is represented as “1” and the minor allele as “2.” Significant p-values (before multiple testing corrections) and odds ratios are shown in bold.
SNP, single-nucleotide polymorphism; CI, confidence interval.
None of the five representative SNPs were associated with obesity based on analysis of genotypes in the obesity case–control subjects (data not shown), and none of the SNPs were associated with BMI or type 2 diabetes (at any age) based on genotypes from the family-based study (data not shown).
The D′ and r 2 values between these five representative SNPs, based on genotypic information from the 1,311 family-based subjects, are shown in Figure 2. All five SNPs were in high linkage disequilibrium (D′≥0.9). The concordance rates (r 2) were also high among the Thr92Ala variant, rs225011, and rs225015 (MAF=0.18–0.19), but the r 2 differed for the less-frequent variants rs6574549 and the novel 5′ flanking SNP. This is consistent with what was observed with the early-onset type 2 diabetes association, where Thr92Ala, rs225011, and rs225015 all showed similar significant levels of association to this disease (before multiple testing correction).
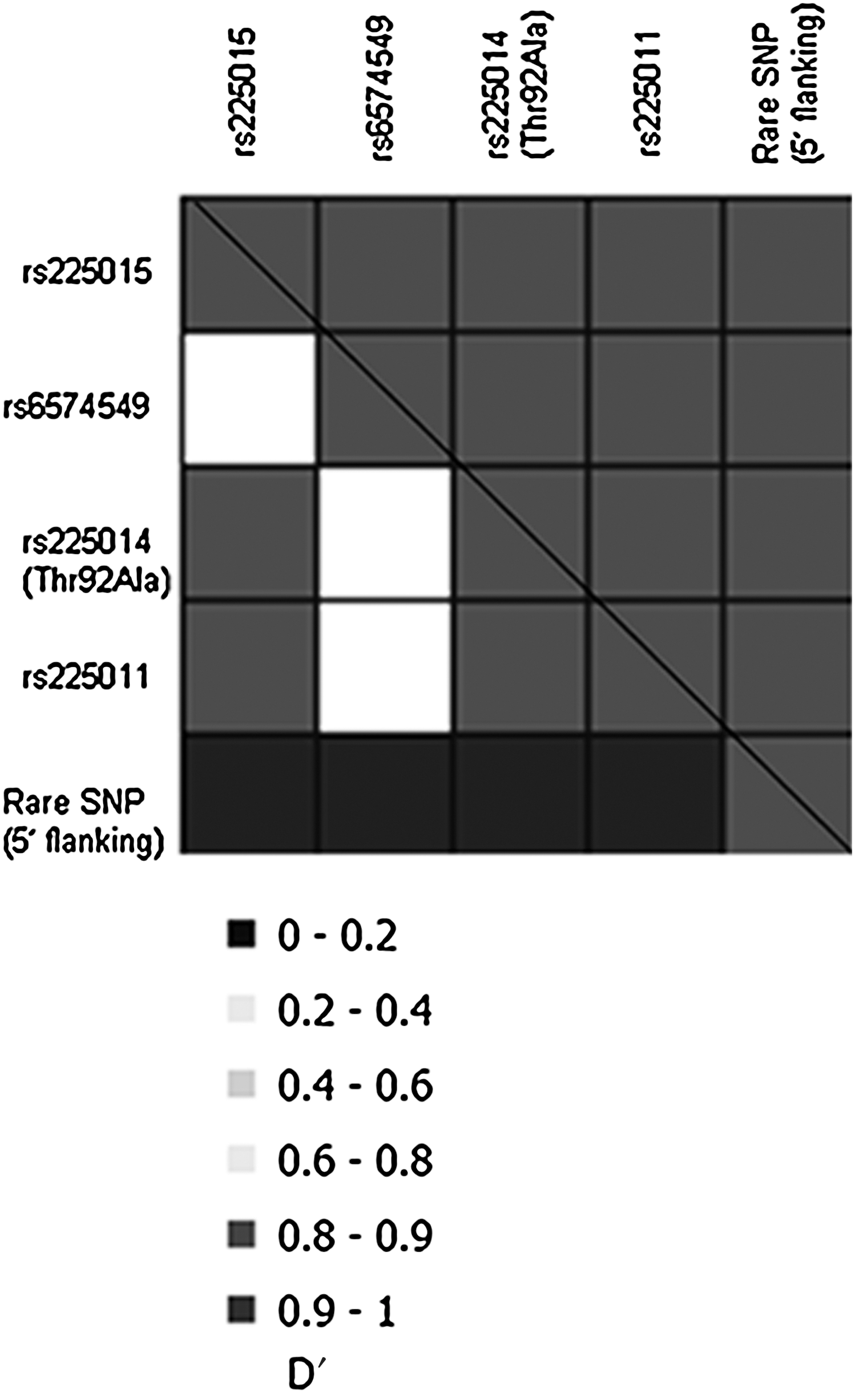
Pairwise linkage disequilibrium between genotyped SNPs. D' values are represented by upper right half and r 2 values are represented by the lower left half of the box. Genotypes of the five representative SNPs in DIO2 that were typed in 1,311 subjects (family-based study) were used to determine pairwise linkage disequilibrium.
Genotypic data from the five representative SNPs were further analyzed for association with metabolic predictors of type 2 diabetes among 256 nondiabetic subjects. Data from the novel 5′ flanking SNP were not analyzed because it was too rare (among 256 subjects, only 2 heterozygotes were identified). Measures of insulin action from both an OGTT and hyperinsulinemic-euglycemic clamp were analyzed in addition to measures of energy expenditure from a human respiratory chamber. All four of the representative variants were nominally associated with the rate of basal endogenous glucose output (Table 2; only data for Thr92Ala and rs6574549 are shown since Thr92Ala provided similar results to rs225011 and rs225015). For the SNPs represented by Thr92Ala, rs225011, and rs225015 variants, the risk allele for early-onset type 2 diabetes is associated with a lower mean basal endogenous glucose output rate. For the SNPs represented by rs6574549, the risk allele (A) was similarly associated with a lower basal endogenous glucose output rate. Although individuals homozygous (AA) for the risk allele of rs6574549 had lower basal endogenous glucose output rates and increased rates of sleeping energy expenditure, they had higher fasting insulin levels (37 vs. 30 μU/mL) and lower glucose disposal rates (high dose) (8.2 vs. 9.2 mg/kg EMBS/min) compared with the heterozygous individuals (AC) (Table 2). However, after corrections for multiple testing, none of the associations remained statistically significant.
Values for fasting and 2-hour plasma insulin concentrations and glucose disposal rate during low-dose insulin infusion were log transformed before analyses to approximate a normal distribution. Data for these variables are presented as geometric means (95% CI, low-high). All other variables are mean±standard error.
Analyzed under a recessive model (AA vs. AC + CC) due to the low frequency (n=1) of subjects with CC genotype.
Adjusted for age, sex, and nuclear family membership.
Adjusted for age, sex, %fat, and nuclear family membership.
Adjusted for age, sex, fat mass, fat-free mass, and nuclear family membership.
Fewer subjects underwent the respiratory chamber measurements.
p Δ, p-value corrected for multiple testing; EMBS, estimated metabolic body size.
We further looked for an interaction of the Thr92Ala polymorphism in DIO2 with the Trp64Arg polymorphism in the ADRB3 gene in determining BMI in case–control and family-based datasets in the Pima Indians. No significant association was identified (Table 3).
Subjects homozygous and heterozygous for each variant are reported together.
Discussion
In our study of Pima Indians, we found that several variants in DIO2, including the Thr92Ala, are weakly associated with early-onset type 2 diabetes with an onset age of less than 25 years, but this association is not significant for type 2 diabetes as defined by onset at any age. We have previously shown that type 2 diabetes is more heritable at younger ages (38) where there are fewer confounding long-term environmental influences, and this could explain our ability to detect a subtle association in the more powerful young-onset case–control group but not in the large family-based study. Further, we did not detect an association with these variants and either obesity or BMI as a continuous trait, which was somewhat unexpected since thyroid hormone action has a role in energy homeostasis. Thyroid hormones interact synergistically with the adrenergic system in adaptive thermogenesis, and variations in the β-3-adrenergic receptor (ADRB3) are associated with body weight gain (39) and time of onset of type 2 diabetes (40). It has previously been reported that there is an association with body weight and BMI in subjects carrying both the DIO2 Thr92Ala variant and the ADRB3 Trp64Arg variant (21). This interaction was not replicated in our study of the Pima Indians or in another large cohort of more than 7000 Danish white subjects (22).
It has also been shown that carriers of Ala/Ala genotype of Thr92Ala have higher fasting insulin levels as well as estimated HOMA-IR in other populations (21,23). Hence, we analyzed associations of the Thr92Ala and other variations with direct measurements of insulin-mediated glucose uptake that may support the risk for young-onset diabetes. None of the variations that were associated with young-onset diabetes were significantly associated with either fasting insulin levels or rates of insulin-mediated glucose uptake. However, the representative SNP rs6574549 that was in high D′ but lower r 2 with these other variants had statistically significant (nominal, but not corrected p-values) associations with these insulin-related phenotypes. This SNP was also nominally associated with sleeping energy expenditure. Surprisingly, rs6574549 was not itself associated with early-onset type 2 diabetes. The low frequency of this variant (minor allele=0.06) may account for this inconsistency and the associations with measures of insulin action and energy expenditure may be false positives, or alternatively, the lack of association with this variant and early-onset type 2 diabetes may be a false negative.
All the variants in DIO2 were weakly associated (statistically significant nominal, but not corrected p-values) with endogenous rates of hepatic glucose output with the carriers of the risk alleles having lower rates. It has been demonstrated that elevated levels of plasma T3 can stimulate hepatic gluconeogenesis (41). There could be lower hepatic gluconeogenesis in subjects carrying the diabetes risk alleles if they had lower physiologically active plasma or intracellular T3. In this study we did not have measures of thyroid-stimulating hormone (TSH), T3, or T4 for the genotyped subjects. However, in a separate group of nondiabetic Pima Indian subjects (n=89), neither Thr92Ala nor rs6574549 was significantly associated with measured plasma levels of TSH, free T3, free T4, or T3/T4 ratio (42).
We cannot exclude the possibility that the weak associations we observed in this study may be due to linkage disequilibrium with a functional SNP in the introns of DIO2, another gene, or a regulatory element in the region. In summary, our data support a minor role of DIO2 in contributing to insulin resistance, thereby increasing susceptibility to early-onset type 2 diabetes, but we do not believe that this is a major determinant for either type 2 diabetes or obesity among this Native American population.
Footnotes
Acknowledgments
The authors gratefully acknowledge the volunteers and leaders of the Gila River Indian Community, whose cooperation made these studies possible. The authors also acknowledge Dr. Joy Bunt and the nurses of the Clinical Research Center, and Dr. Arline Salbe and the Metabolic Kitchen staff, for the care of the research volunteers. The authors sincerely thank Dr. Robert Hanson for performing the power analyses, as well as help in conducting the multiple testing corrections in this study. This research was supported by the Intramural Research Program of the National Institute of Health, NIDDK.
Disclosure Statement
No competing financial interests exist.