
Abstract
Introduction:
Many tissues express thyroid hormone metabolizing deiodinases that both activate and inactivate thyroid hormones through conversion processes. Many believe that the primary role of thyroid hormone deiodinases is the activation of the prohormone thyroxine (T4) to the active hormone triiodothyronine because athyreotic humans can be treated with T4 alone. In our hands a nonspecific deiodinase inhibitor (iopanoic acid [IOP]) decreased cutaneous cell proliferation in vitro, so we hypothesized that topical IOP would inhibit epidermal proliferation in vivo.
Methods:
IOP was applied topically to mice. Treatments were applied daily for 1 week. Skin biopsies were either stained for 5-bromo-2-deoxyuridine or flash-frozen to assay for deiodinase activity.
Results:
Topical IOP resulted in a dose-dependent increase in epidermal proliferation. Assay revealed significant inactivating type 3 deiodinase (Dio3) activity in the epidermis but little or no activating (Dio1 or Dio2) activity. Dio3 activity was decreased 44%±21% in epidermis from mice treated with low-dose IOP and 80%±4% in epidermis from mice treated with high-dose IOP (p<0.001).
Conclusion:
We hypothesize that keratinocytes express Dio3 in vivo to maintain cutaneous health and prevent the skin from becoming hyperproliferative. Our data support the developing recognition that the primary role of thyroid hormone deiodinases in some tissues may be the degradation of thyroid hormone to protect the tissue against thyrotoxicosis.
Introduction
T3 effects on skin depend on route of delivery. In contrast to topical T3, systemically induced thyrotoxicosis in rodents results in the skin thinning and hair loss traditionally associated with excess T3 (3) along with associated collagen loss (8). There are three classic iodothyronine deiodinase enzymes (9,10). Two of the enzymes (type 1 and type 2 deiodinases, Dio1 and Dio2) are responsible for activating the pro-hormone thyroxine (T4) to the active hormone T3 and degrading T3 to its inactive by-product 3,3′-diiodothyronine (3,3′T2). Dio1 and Dio2 differ in how their activities and expression levels are regulated. Dio1 can also serve as an inactivating enzyme by converting T4 to reverse T3 (rT3). The primary role of the third enzyme, Dio3, is to convert T4 to inactive rT3. All 3 deiodinases further metabolize the tri-iodothyronines to the di-iodothyronines. Dio1 and Dio3 are located in the plasma membrane, whereas Dio2 is located in the endoplasmic reticulum.
Previous investigators showed conversion of T4 to either T3 or rT3 in skin cultures, thus demonstrating indirectly the presence of thyroid hormone deiodinases in skin (11 –13). Specific Dio2 activity has been demonstrated in cultured human fibroblasts (14). Neither Dio1 nor Dio3 was found to be active in the dermal fibroblasts. Many believe that the primary role of thyroid hormone deiodinases is the activation of the prohormone T4 to the active hormone T3 because athyreotic humans can be treated with T4 alone. While Dio1 and Dio2 are expressed in multiple tisses, it had been thought that Dio3 was expressed in a more limited fashion. Older data suggested Dio3 expression only in placenta and during major illness. Newer data have demonstrated a protective role for Dio3 in some tissues, including cochlea (15) and photoreceptors (16).
Because athyreotic humans can be treated with T4 alone, we anticipated some conversion of T4 to T3 in epidermis. Further, we previously determined that the nonspecific thyroid hormone deiodinase inhibitor, iopanoic acid (IOP), could inhibit cultured keratinocyte proliferation (17).
Therefore, we hypothesized that topical application of IOP would block epidermal proliferation in vivo. Although we and others have identified Dio3 in human skin and goat epidermis (18 –20), we predicted that the primary impact of cutaneous deiodinase inhibition in vivo would be to block proliferation based on our in vitro studies.
Materials and Methods
Animal handling
All animal experimentation was conducted in accordance with accepted standards of humane animal care in a protocol approved by the Institutional Animal Care and Use Committee at Boston University School of Medicine.
Topical IOP preparation and dosing
IOP (Sigma) was dissolved into a modestly basic stock solution to 2 final concentrations: Low-dose IOP treatment was 0.025 mM NaOH, 0.2 M NaCl, and 0.3 mM IOP. High-dose IOP treatment was 0.025 mM NaOH, 0.2 M NaCl, and 1.5 mM IOP.
The topical IOP cream was prepared by mixing the IOP into a liposome vehicle (Novasome A, IGI Inc.). Each mouse received 30 μL of the liposome vehicle applied directly to a 3×3 cm area of skin on the animal's backs. The preparation was applied daily and not removed.
Fifteen hairless mice (SKH-1, Charles River) were used in the first experiment. To control for hairless mouse strain-specific findings, a second experiment was done with 15 CD-1, normally haired, mice (Charles River). The 15 mice in each strain were divided into three treatment groups of five mice each. Mice were treated with either low-dose IOP, high-dose IOP, or control vehicle with no drug.
On day 1, mice were biopsied and treated with their respective interventions. Topical treatments were applied once daily for 1 week. At the end of the week, the mice were injected with 5-bromo-2-deoxyuridine (BrdU; Boehringer Mannheim) and biopsied 3 hours afterward. BrdU staining was performed on slides prepared from the skin biopsies using immunohistochemistry.
Protocol to control for systemic IOP action
In a third experiment, additional SKH-1 hairless mice were randomized into three groups. The first group was treated with intraperitoneal (IP) IOP, the second group was treated with the topical IOP cream, and the third group was treated with topical vehicle alone.
Like the first experiment, on day 1, mice were wounded, biopsied, and treated with their respective interventions. Treatments were given once daily for 1 week.
Evaluation of hair number and epidermal proliferation
Animals were biopsied with full thickness dorsal skin samples taken. Skin samples were fixed in formalin and embedded in paraffin. From each paraffin block, 5 μm sections were made and stained with hematoxylin and eosin. For each histology section, the number of hair shafts that opened up to the epidermal surface was quantified in random 1-mm segments of epidermis. This was repeated 10 times per histological section. In addition, 5 μm skin histology sections from the paraffin blocks were stained for BrdU. BrdU data are reported as the number of cell nuclei stained/mm epidermis.
Type 3 iodothyronine deiodinase assays
Biopsied mouse skins were flash frozen in liquid nitrogen and homogenized in five volumes of 1.0M sucrose, 10 mM dithiothreitol in phosphate EDTA buffer (pH 6.9). Each deiodination reaction included 3 to 150 μg of homogenized tissue sonicate, 200,000 cpm of 3,3′,[125I]5′-triiodothyronine (New England Nuclear), 10 mM dithiothreitol, and various concentrations of unlabeled T3. The reaction was stopped by the addition of methanol. The labeled substrate and deiodination products were quantified with high performance liquid chromatography.
Type 1 and 2 deiodinase assays
Each deiodination assay required 25 μg of tissue sonicate, 100,000 cpm of 3,3′,[125I]5′-triiodothyronine and 3,5,3′[125I]5′-triiodothyronine (New England Nuclear) for Dio1 and Dio2 assays, respectively. In addition, 10 mM dithiothreitol along with a dose range of concentrations of cold unlabeled T3 were added to the reaction mix.
The master-mixes included nonradiolabeled rT3 as substrate for the Dio1 assay and cold T4 as substrate for the Dio2 assay with one part EDTA buffer for each sample. Individual samples were vortexed and incubated overnight at 37°C for 22 hours. Free radiolabeled iodothyronines were quantified the next day in a gamma counter.
Statistical analysis
Where applicable, statistical analysis was performed with Student's t-test (for paired comparisons) and analysis of variance (for dose–response comparisons). All data are reported as the mean result for each group of animals with error bars representing standard error of the mean.
Results
Topical treatment with a nonspecific deiodinase inhibitor-stimulated epidermal proliferation in mice in vivo
We used BrdU incorporation into DNA as a surrogate for epidermal proliferation. Were keratinocytes to express primarily activating deiodinases (e.g., Dio2), inhibition of proliferation would be predicted to occur with topical IOP treatment. However, there was a significant increase in proliferation (Fig. 1) in the topical IOP-treated groups versus controls, as would be predicted if metabolism of T3 were inhibited (86.9±16 BrdU stained nuclei/mm vs. 8.6±1.79, p<0.01).
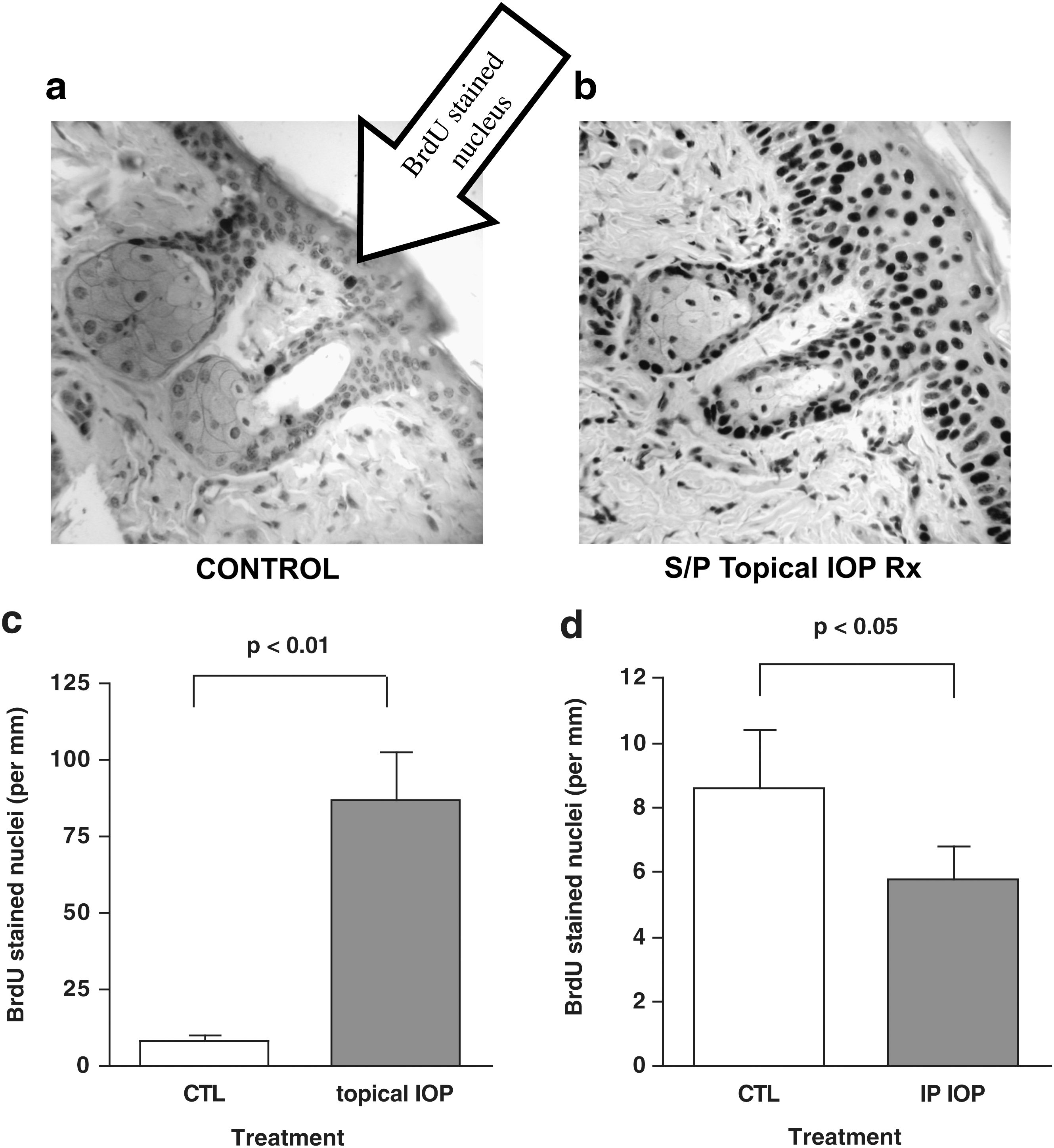
Increased proliferation evident in intraperitoneal iopanoic acid (IOP) –treated epidermis.
Systemic IOP resulted in decreased epidermal proliferation
To control for nonspecific systemic effects of IOP, we undertook a study to establish the impact of systemic IOP (via IP injection) relative to control mice to mice receiving topical IOP treatment. In humans, systemic IOP causes hypothyroidism by blocking T4 to T3 conversion.
Indeed, when we measured serum thyroid hormone levels with standard immunoassay kits (ICN), we found decreased T4 and T3 concentrations in mice treated with IP IOP. By contrast T4 and T3 concentrations were not significantly altered in mice receiving topical IOP treatment (T4 levels for control mice 7.4±1.5 μg/dL, for IP IOP-treated mice 5.7±3.8, and for topical IOP-treated mice 8.5±2.3; T3 levels for control mice 66±14 ng/dL, for IP IOP-treated mice 12±31, and for topical IOP-treated mice 64±7.5).
As expected, epidermal keratinocyte proliferation was decreased in the IP IOP group relative to controls (5.8±1.0 BrdU stained nuclei/mm versus 8.6±1.79, p<0.05). In addition, it was noteworthy that the systemic effect was opposite to the topical treatment effect.
Topical IOP treatment resulted in increased hair counts
We used hair growth as a second surrogate for epidermal proliferation. There was a modest but significant dose–response increase in hair growth for SKH-1 mice treated with topical IOP relative to controls (3.2±0.2 hair shafts/mm for mice receiving high-dose IOP, 2.8±0.2 hair shafts/mm for mice receiving the low-dose IOP, and 2.1±0.2 for mice in the control group, p<0.005, Fig. 2).
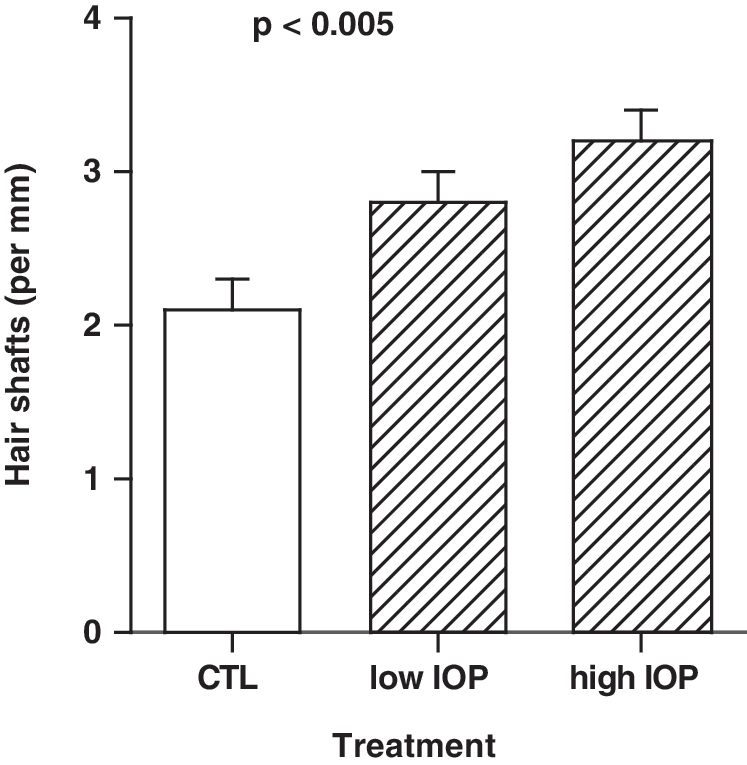
Increased hair growth demonstrated in IOP-treated animals. Topical IOP resulted in a significant dose-dependent increase in hair shaft number (p<0.005, ANOVA). ANOVA, analysis of variance.
Significant type 3 deiodinase activity was observed in the hairless mouse strain
To determine if thyroid hormone metabolism could play a role in epidermal proliferation, we assayed for deiodinase activity. Type 3 deiodinase activity was detected in SKH-1 mouse epidermis (1.03±0.27 fmols/mg/minute). By contrast, neither Dio1 nor Dio2 activity was detected in SKH-1 mouse epidermis.
IOP inhibited deiodinase activity
Dio3 activity was decreased 44%±21% in epidermis from mice treated with low-dose IOP and 80%±4% in epidermis from mice treated with high-dose IOP (p<0.001, Fig. 3).

Topical IOP resulted in a significant decrease in type 3 deiodinase (Dio3) activity (p<0.001, ANOVA).
Deiodinase findings were replicated in a haired mouse strain
To control for mouse strain and to demonstrate that the deiodinase activity pattern was not SKH-1 strain specific, the topical IOP treatment experiments were repeated using a haired mouse strain (CD-1; Charles River).
In CD-1 mice, the BrdU incorporation assay and the hair count assay were consistent with increased epidermal proliferation in mice treated with topical IOP relative to control mice. Like with the hairless mice, there was a significant, dose-dependent increase in BrdU stained nuclei in mice treated with topical IOP. In addition, there was a modest but significant dose–response increase in hair growth for CD-1 mice treated with topical IOP relative to controls.
Finally, Dio3 activity was easily detected in CD-1 epidermis (3.5±0.15 fmols/mg/minute) but neither Dio1 nor Dio2 activity was detected. In mice treated with topical IOP, Dio3 activity was decreased in dose-dependent fashion (down 33%±19% for mice treated with the low IOP dose, and down 97%±7% for mice treated with the high IOP dose, p<0.001).
Discussion
Thyroid hormone action on epidermis is fundamentally dependent on the relative intracellular concentration of the active thyroid hormone, T3. The predominant circulating thyroid hormone is the pro-hormone, T4. While some tissues respond directly to circulating serum T3, many tissues (including epidermis and dermis) depend on local T4 conversion to the active thyroid hormone T3 by iodothyronine deiodinases (9,10). In addition, individual tissues express the iodothyronine deiodinases that metabolize both T4 and T3 to inactive metabolites.
As classically understood, the primary role of Dio3 is to protect tissues against the increased metabolism that comes with increased thyroid hormone levels. Dio3 is known to be expressed in sick animals with the expectation that it serves to protect tissues from significant demand during a weakened state. Dio3 is also expressed in placenta where it is presumed that it protects the developing fetus against potential excess thyroid hormone from the mother.
For years, it was presumed that Dio3 only served in the roles noted and that there was no significant Dio3 activity in the healthy adult.
We hypothesize that the data presented here support the data by Ng at al. (15,16) and others for tissue-specific expression of cutaneous deiodinases wherein the primary role of deiodinases in some tissues is to inactivate thyroid hormone. In vivo, keratinocytes, which do not have a blood supply, express the inactivating deiodinase (Dio3) instead of the activating deiodinase (Dio2) expressed in the abutting cells that separate keratinocytes from their blood supply (Fig. 4).

Model of mechanism underlying increased cutaneous proliferation in response to topical IOP.
Were the primary deiodinases expressed in epidermis activating (e.g., Dio2 and sometimes Dio1), IOP might be predicted to inhibit T4 to T3 conversion and epidermal proliferation like we observed with keratinocytes in vitro (17).
Alternatively, if the primary deiodinases expressed in epidermis were inactivating (e.g., Dio3 and sometimes Dio1), IOP might be predicted to inhibit metabolism of T3, which would maintain intracellular T3 and stimulate proliferation.
To our surprise, we demonstrated dramatic stimulation of epidermal proliferation after topical application of IOP. The stimulation contrasted with the systemic impact of IOP. The stimulation was dose-dependent. The stimulation was not mouse strain specific.
We and other investigators reported that skin might express both the type 2 and type 3 iodothyronine deiodinases (11,18 –20) and that dermal fibroblasts specifically express only the type 2 deiodinase (14).
In addition, previous investigators demonstrated that in a transformed epidermal cell line, increased Dio3 expression resulted in stimulated proliferation (21). The thought was that T3 acted to stimulate cell differentiation and the premature metabolism of T3 facilitated greater proliferation. Whether the finding could be extended to normal epidermis in vivo remained unclear.
In this in vivo study, we demonstrated significant Dio3 iodothyronine deiodinase activity in murine epidermis but negligible Dio1 or Dio2 deiodinase activity. Then we demonstrated that topical IOP treatment resulted in a dose–response decrease in the type 3 deiodinase activity. Our data suggest that neither our keratinocyte cell culture model (17) nor that of Dentice et al. (21) exactly reflects the in vivo state. Our current hypothesis is that dermal factors not present in the in vitro models play an important role in expression of deiodinases.
Our data do not completely exclude the possibility of some Dio2 deiodinase expression in murine epidermis in vivo. A recent study demonstrated Dio2 activity in murine muscle that was only detected with a modified Dio2 assay to increase fidelity (22).
The data suggest that in epidermis, the type 3 thyroid hormone deiodinase plays a rate limiting role wherein local thyroid hormone concentration is regulated by the rate of degradation to inactive metabolites.
We propose that epidermal keratinocytes express Dio3 in vivo to maintain cutaneous health and prevent the skin from becoming hyperproliferative. When the type 3 deiodinase activity is inhibited, cell proliferation can increase.
We further propose that even a nonspecific deiodinase inhibitor may be a useful therapeutic intervention in circumstances where there is narrow expression of deiodinase type and effect can be predicted.
More broadly, it is possible that Dio3 serves to buffer the body against thyrotoxicosis in general. The skin is the largest organ in the body and could serve such a significant role. Alternatively, Dio3 expression may be regulated in a tissue specific fashion throughout the body. In such a scenario, Dio3 would join with the activating deiodinase (Dio2) in providing exquisite local control over thyroid hormone levels. In the latter case, elegant symbiosis between tissues may have evolved where the epidermis maintains Dio3 activity to buffer both itself and adjacent dermis against thyrotoxicosis.
Footnotes
Acknowledgments
The authors wish to thank Ann Marie Zavacki and Rafael Arrojo e Drigo for assistance with deiodinase assays.
Disclosure Statement
The authors declare that no competing financial interests exist.