
Abstract
Background:
The occurrence of familial papillary thyroid cancer (FPTC) is well established but no susceptibility genes for this disease have been discovered. Our group has recently demonstrated that patients with FPTC have shorter telomeres, not associated with mutations in telomerase reverse transcriptase, gene than patients with sporadic papillary thyroid cancer (SPTC), healthy subjects (HS), and unaffected family members (UFMs). Several diseases, however, have short telomeres associated with mutations in the telomerase RNA component (TERC) gene or in the shelterin complex (POT1, RAP1, TIN2, TPP1, TRF1, and TRF2) genes. The objective of the present study was to verify whether short telomeres observed in FPTC patients were related to mutations in TERC or shelterin genes.
Methods:
Sixty-six patients with FPTC, 46 UFMs, 111 patients with SPTC, and 153 HS were analyzed by polymerase chain reaction followed by denaturing high performance liquid chromatography analysis and direct sequencing for the presence of TERC or shelterin gene mutations. When present, single-nucleotide polymorphisms were tested by χ 2 analysis at the genotypic, allelic, and haplotypic levels.
Results:
The entire sequence of the TERC gene was analyzed with particular attention to known mutations known to be associated with short telomeres. All samples appeared to be homozygous wild type for A-771G, C-99G, G305A, G322A, C323T, C408G, G450A, T467C, G508A, A514G, G623A, and C727G substitutions and for the 378Δ→3′ deletion in the TERC gene. In addition, upon analysis of all samples for shelterin proteins, we observed a significant decrease in POT1 and RAP1 protein expression in the blood of FPTC patients compared with SPTC subjects. However, no mutations or polymorphisms were found when in the coding sequences of both genes.
Conclusions:
To our knowledge this is the first study of TERC mutations or alterations in the shelterin complex in relation to FPTC. Shorter telomeres observed in FPTC are not linked to mutations or polymorphisms in TERC, POT1, or RAP1 genes.
Introduction
Short telomeres and inherited or acquired genetic defects in telomere maintenance have been associated with an increased risk of developing familial diseases such as dyskeratosis congenita (DC) syndrome (14) and familial forms of cancer such as head, neck, lung, breast, and renal cancers (15). Several mutations in telomerase RNA component (TERC) gene are associated with benign and malignant diseases such as DC (16), aplastic anemia (17,18), systemic sclerosis (19), myelodysplastic syndrome (18), and generally with a reduction in mean telomere length (20). Similarly, recent studies have highlighted an important role of the shelterin complex in cancer development (21). This complex is formed by six proteins that normally contribute to shape and safeguard human telomeres. In particular three shelterin subunits, TRF1, TRF2, and POT1, directly recognize the TTAGGG repeats. They are, moreover, interconnected by three additional shelterin proteins, TIN2, TPP1, and RAP1, forming a complex that allows cells to distinguish telomeres from extra-telomeric sites of DNA damage. Without the protective activity of shelterin, telomeres are no longer hidden from the DNA damage control mechanisms and chromosome ends are inappropriately processed by DNA repair pathways (22).
Based on these observations, the aim of the present work was to determine whether there are variations in TERC gene in the peripheral blood of patients with FPTC, perhaps explaining the telomere shortening observed in these patients. Shelterin proteins were also investigated to determine whether there were alterations in mRNA expression or the presence of gene mutations.
Patients and Methods
After informed consent in accordance with local ethical committee guidelines, blood samples for DNA analysis were collected from 66 patients with FNMTC (referred here as FPTC from the moment that all display the papillary histotype of the disease) (belonging to 38 kindreds), 46 unaffected family members (UFMs) (belonging to 23 kindreds), 111 sporadic papillary thyroid cancer (SPTC) patients, and 153 healthy subjects (HS). FPTC was defined as the presence of at least one first-degree relative with DTC. The HS were selected from volunteers of the “Blood donor centre” of the hospital of Siena (Italy). In these subjects, autoimmune diseases, cardiovascular diseases, and diabetes were excluded. Patients and HS with a history of radiation exposure and malignancies were excluded.
DNA extraction
Genomic DNA was extracted using salting out procedures. For each sample DNA concentration was assessed by spectrophotometry and stock solutions of 200 ng/50 μL were prepared and used for following experiments.
Search for TERC mutation
To determine the presence of TERC mutations, 25 μL/samples of AmpliTaq Gold PCR Master mix (Applied Biosystems) was been added to 1.5 mM MgCl2, 400 μM dNTPs, and 300 nM of specific primers (primer sequences, annealing temperatures, and denaturing high performance liquid chromatography [DHPLC] conditions are available upon request) designed to sequence the entire gene in a final volume of 50 μL. Polymerase chain reaction (PCR) products were visualized with ethidium bromide in a 2% agarose gel. Samples were subsequently denatured (10 minutes at 95°C followed by 5 minutes at 56°C) and analyzed with the DHPLC (Transgenomic Wave Nucleic Acid Fragment Analysis System-Transgenomic Inc.) technique to confirm the presence/absence of mutations using specific temperatures and applying gradient conditions as calculated by the Navigator™ software (Vers. 1.6.4). Samples with different elution profile were subjected to direct pyrosequencing (99.5% accuracy) (Primm).
Shelterin protein expression
RNA was extracted from fresh blood of 51 FPTC and 82 SPTC patients using QIAamp RNA Blood Mini Kit (Qiagen). One microgram of each sample was retrotranscribed into complementary cDNA using iScript cDNA Synthesis Kit (Biorad) and 200 ng/μL was evaluated by real-time PCR using the MJ Mini Thermocycler (Biorad) in a mix containing 2× iQ™ Supermix (Biorad) and 20× TaqMan primer/probes (Applied Biosystems) in a final volume of 25 μL. Annealing temperature was 60°C for 35 cycles. Each sample was run in duplicate and for each run efficiency of real time PCR (RT-PCR) (E), slope values, and correlation coefficients (R 2) were determined. The expression level was calculated as ΔΔCt and reported as 2−ΔΔCt against beta-actin and RPL13 chosen as reference genes by two different software programs for the selection of optimal control genes in qRT-PCR studies, Normfinder and GenNorm. The GenNorm provides a ranking of the tested genes, based on their expression stability, determining the two most stable reference genes or a combination of multiple stable genes for normalization. NormFinder identified the optimal normalization genes among a set of candidates according to their expression stability value in a given sample set and a given experimental design.
Search for POT1 and RAP1 point mutations
To determine the presence of POT1 and RAP1 point mutations, specific primers were designed using Vector NTI Software® to cover all the coding sequence of the genes (see Ensembl database). For each sample (66 FPTC, 111 PTC, 46 UFMs, and 153 HS), 200 ng of DNA was amplified in a final volume of 50 μL containing 2× AmpliTaq Gold PCR Master mix (Applied Biosystems) and 300 nM of specific primers (primers and PCR conditions are available upon request). Samples were subsequently denatured and analyzed with the DHPLC (Transgenomic Wave Nucleic Acid Fragment Analysis System-Transgenomic Inc.) technique to confirm the presence/absence of mutations. Samples with different elution profiles were subjected to pyrosequencing (Primm) (DHPLC conditions are available upon request).
Statistical analysis
Statistical analysis was performed using the software package SPSS version 13.0 (IBM Company). Interaction with single-nucleotide polymorphisms (SNPs) was tested by χ 2 analysis at genotypic, allelic, and haplotypic levels. p<0.05 was considered statistically significant. Student's t test was used to calculate the difference in the expression levels of shelterin proteins using StatView for Windows, ver.5.00.1 (SAS Institute).
Results
Search for TERC mutations
Samples were tested for known, generally disease-associated, TERC variations (Table 1) together with a complete sequencing of the entire gene. All of the subjects analyzed (FPTC patients, UFMs, SPTC patients, and HS) were homozygous wild type for A-771G, C-99G, G305A, G322A, C323T, C408G, G450A, T467C, G508A, A514G, G623A, and C727G substitutions and for the 378Δ→3′ deletion.
TERC, telomerase RNA component.
We found two polymorphisms indicated as polymorphism #1 (SNP rs2293607) and polymorphism #2 (SNP rs35073794) of unknown pathogenic significance (Ensembl database). Polymorphism #1 consisted of n501 (T>C) substitution (forward strand), and was observed in the heterozygous form (TC) in 48/153 (31.4%) of HS, 26/66 (39.4%) of FPTC patients, 17/46 (37%) of UFMs, and 35/111 (31.5%) of SPTC patients (Table 2). The homozygous variant (CC) was detected in 16/153 (10.5%) of HS, 4/66 (6.1%) of FPTC patients, 6/46 (13%) of UFMs, and 9/111 (8.1%) of sporadic PTC patients (Table 2). The different distribution of this polymorphism in the four groups was not statistically significant (Pearson chi-square p=0.714).
T=Wt allele and C=polymorphic allele for polymorphism #1; G=Wt allele and A=polymorphic allele for polymorphism #2.
FPTC, familial papillary thyroid cancer.
Polymorphism #2 was more rare and consisted of n701 (G>A) (forward strand) substitution. It was found as heterozygous (GA) in 4/153 (2.6%) of HS, 4/66 (6.1%) of FPTC patients, 3/46 (6.5%) of UFMs, and 2/111 (1.8%) of SPTC patients (Table 2). The homozygous variant (AA) was detected only in 1/153 (0.7%) of HS (Table 2). Also in this case, polymorphism frequency was not statistically different among groups (Pearson chi-square p=0.504). In addition, the diplotype distribution obtained by considering the combination of polymorphisms #1 and #2 was not different in the various groups (Table 3) as well as the single allele frequency of both polymorphisms (Table 4, p value of 0.576 for polymorphism #1 and a p value of 0.417 for polymorphism #2, respectively) and the association between single alleles (Table 5, p value of 0.640).
When TERC copy number variations were analyzed, we found one sample belonging to the UFMs with two insertions and two deletions (Fig. 1) not reported before. These insertions concern nucleotides 1418 (1418insA) and 1437 (1437insT) while the deletions affected nucleotide 1440–1442 (1440_1442delCTG) and 1461–1467 (1461_1467delGGAAAAA). We did not find two other variations reported in genome databases (i.e., Ensembl) and indicated as CN_996453 and CN_996452. When the search was extended to the entire gene sequence, no mutations or polymorphisms were found.
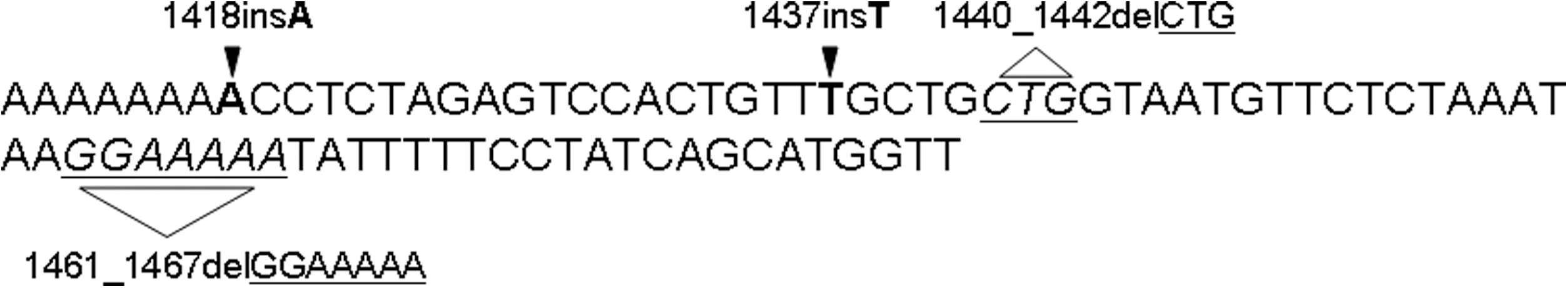
Schematic representation of telomerase RNA component insertions and deletions found in one sample belonging to one unaffected family member.
Shelterin protein complex
We extended our research to the six proteins of the shelterin complex implicated in the regulation of telomere length. We previously analyzed their expression levels in blood of 51 patients with FPTC and 82 patients with SPTC and expressed the results as 2−ΔΔCt with respect to two different reference genes (beta-actin and RPL13). Figure 2 reports the mean expression values for (Fig. 2A) TRF1 (SPTC: 9.39±2.5, FPTC: 13.05±3.2, p=0.9), (Fig. 2B) TRF2 (SPTC: 3.04±1, FPTC: 3.18±0.8, p=0.1), (Fig. 2C) TIN2 (SPTC: 6.16±2.4, FPTC: 11.9±2.7, p=0.05), (Fig. 2D) TPP1 (SPTC: 5.18±0.78, FPTC: 4.89±1, p=0.4), (Fig. 2E) POT1 (SPTC: 6.45±1.2, FPTC: 2.8±0.93, p<0.01), and (Fig. 2F) RAP1 (SPTC: 11.5±1.2, FPTC: 4.9±1, p=0.03). Only for POT1 and RAP1 did we observe a significant decrease in the expression level in patients with FPTC compared with patients with SPTC (Fig. 2). We then looked for point mutations in POT1 and RAP1 genes in the blood of 66 patients with FPTC, 111 patients with SPTC, 46 UFMs, and 153 HS. All samples were homozygous wild type in both genes. We found only the polymorphism rs4888444 in RAP1, with a frequency of 2.9%, in FPTC patients. This was considerably lower than that reported for normal population (20%) and, thus, not associated with the FPTC phenotype.
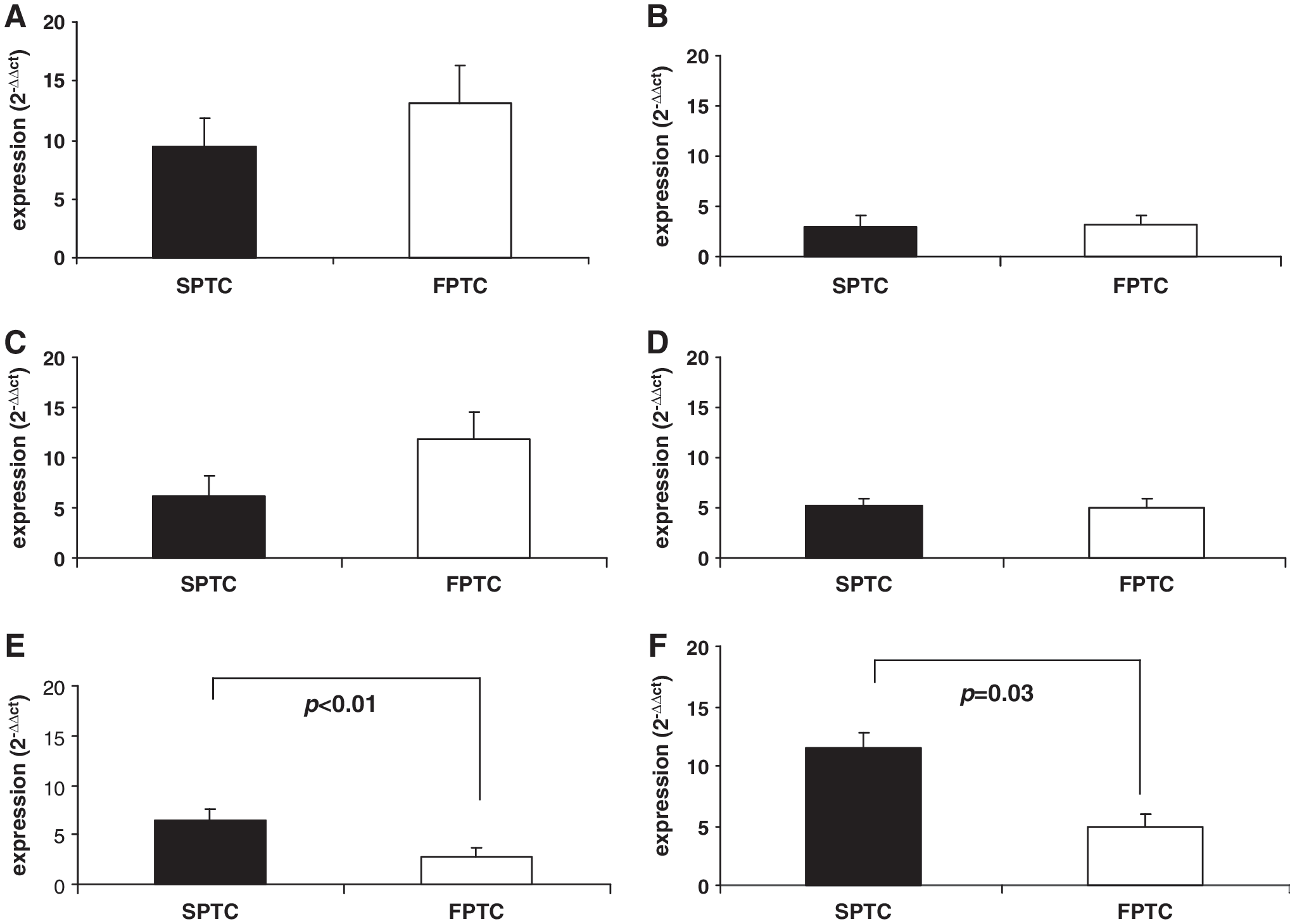
Mean expression levels of the six shelterin proteins expressed as 2−ΔΔCt and calculated in comparison to two different reference genes (beta-actin and RPL13).
Discussion
Telomeres are structures of eukaryotic chromosomes consisting of 6 bp repeats (TTAGGG) that protect chromosome ends from degradation and from end-to-end fusions (23). Telomeres are maintained by telomerase complex composed by an enzymatic component (TERT) and an RNA component (TERC) (24), which acts as template for addition of telomeric repeats. In addition, accessory proteins such as dyskerin, NHP2, NOP110, pontin/reptin, and TCAB1 are found at telomere ends (24). Dyskerin, NHP2, and NOP10 are necessary for the stability and accumulation of TERC, whereas dyskerin collaborates with pontin/reptin to allow the assembly between TERC and TERT. Telomerase cooperates in telomere lengthening together with several proteins such as the shelterin family (TRF1, TRF2, RAP1, TIN2, POT1, and TPP1) and molecular chaperones such as HSP90 (25). The TERC gene maps on chromosome 3 and is characterized by one exon of 438-bp long (Ensembl database). As reported in aplastic anemia (AA) (26) and DC (14, 26), several mutations in TERC gene are associated with telomere shortening particularly those involving the pseudoknot domain (26). In DC, TERC mutations and short telomeres are responsible for the familial form (14, 27). In a previous study we have demonstrated that patients with FPTC have significantly shorter telomeres compared with patients with SPTC, UFM, and HS not related to mutations in TERT gene (12). In this report, we search for known/new TERC gene mutations as possible implicated factor for telomere shortening in FPTC. All samples analyzed, however, were homozygous wild type for TERC mutations with the exception of two SNPs (T501C and G701A) of unknown pathogenetic significance. Statistical analysis of the distribution of the two single polymorphisms, of the combination of both polymorphisms, and of the association between single alleles of polymorphisms #1 and #2 among FPTC patients, SPTC patients, and HS yielded no statistically significant difference. We then extended our research to accessory proteins involved in telomere maintenance. We excluded from this analysis proteins such as dyskerin that has been clearly associated with DC development (28) or hoyeraal-hreidarsson syndrome (29) completely absent in our patients (Genecards database) and focused our attention on shelterin complex. We found a significant reduction in mRNA expression of POT1 and RAP1, not linked to known mutations in both genes.
To our knowledge this is the first report in which mutations in TERC, POT1, and RAP1 have been searched in FPTC and our results exclude that variations of these genes are responsible for telomere shortening observed in familial form of PTC. However, a possible implication of shelterin proteins needs to be further investigated (i.e., miRNA regulation) from the moment that FPTC patients display a decrease in POT1 and RAP1 mRNA expression.
Footnotes
Acknowledgment
Funding: This research did not receive any specific grant from any funding agency in the public, commercial, or not-for-profit sector.
Disclosure Statement
The authors declare that no conflict of interest exists.