
Abstract
Background:
Thyroid hormones (THs) are known to regulate protein synthesis by acting at the transcriptional level and inducing the expression of many genes. However, little is known about their role in protein expression at the post-transcriptional level, even though studies have shown enhancement of protein synthesis associated with mTOR/p70S6K activation after triiodo-
Methods:
INS-1E cells were maintained in the presence or absence of T3 (10−6 or 10−8 M) for 12 hours. Thereafter, insulin concentration in the culture medium was determined by radioimmunoassay, and the cells were processed for Western blot detection of insulin, eukaryotic initiation factor 2 (eIF2), p-eIF2, eIF5A, EF1A, eIF4E binding protein (4E-BP), p-4E-BP, p70S6K, and p-p70S6K.
Results:
It was found that, in parallel with increased insulin generation, T3 induced p70S6K phosphorylation and the expression of the translational factors eIF2, eIF5A, and eukaryotic elongation factor 1 alpha (eEF1A). In contrast, total and phosphorylated 4E-BP, as well as total p70S6K and p-eIF2 content, remained unchanged after T3 treatment.
Conclusions:
Considering that (i) p70S6K induces S6 phosphorylation of the 40S ribosomal subunit, an essential condition for protein synthesis; (ii) eIF2 is essential for the initiation of messenger RNA translation process; and (iii) eIF5A and eEF1A play a central role in the elongation of the polypeptidic chain during the transcripts decoding, the data presented here lead us to suppose that a part of T3-induced insulin expression in INS-1E cells depends on the protein synthesis activation at the post-transcriptional level, as these proteins of the translational machinery were shown to be regulated by T3.
Introduction
It is important to note that, besides p70S6K, there are many other proteins engaged in the initiation and ending processes of mRNA decoding. These are known as translational factors. The initiation factors promote coupling of the transcripts with the ribosomal subunits, whereas the elongation factors insert amino acids into the nascent polypeptide chain (7,8).
With regard to the initiation factors, the eukaryotic initiation factor 2 (eIF2) is one of the most extensively studied. This protein forms a ternary complex with guanosine
mRNAs are coiled molecules that need to be unwound before their coupling with the 43S complex, a process that requires specific translational proteins. One of these proteins is the eIF4E, which is attached to specific cytoplasmic proteins, known as eIF4E binding proteins (4E-BPs). This attachment inhibits eIF4E and impairs protein synthesis initiation. Therefore, 4E-BP phosphorylation has been recognized as an essential step in mRNA decoding, as its phosphorylation leads to dissociation between eIF4E and 4E-BP, a process that releases the translation factor 4E which initiates protein synthesis (11).
After ribosome formation, the next step is the elongation of the polypeptide chain, which depends on the binding of the eukaryotic elongation factor 1 alpha (eEF1A) to the aminoacyl tRNAs. This complex is transported to the ribosomes, and the anticodon sequence of the aminoacyl tRNA is coupled to the adequate codon of the mRNA molecule, allowing the insertion of amino acids to the nascent polypeptide chain (12,13).
It is known that THs rapidly increase the amount of eEF1A anchored to cytoskeleton proteins, which may improve mRNA stability and translation (14). Besides eEF1A, other proteins also participate in the elongation process, such as the eIF5A. Despite being acknowledged as an initiation factor, eIF5A is also involved in the elongation phase of protein synthesis. In fact, the elongation factor 2 (eEF2) inhibitor, sordarin, blunts the effect of eIF5A on yeast growth, indicating that eIF5A might function together with eEF2 to promote ribosomal translocation (15,16).
Although studies from our laboratory have shown that INS-1E cells subjected to T3 treatment did not have altered proinsulin mRNA content (17), our preliminary data is that insulin in this cell type increases after T3 treatment. However, the mechanism of this is unknown.
There have been few studies of the effects of THs on beta cells. Knowledge on their actions is limited to the enhancement of glucose induced insulin secretion and control of beta-cell viability through the regulation of pro-mitotic and pro-apoptotic factors (18,19). Therefore, in this study, we attempted to evaluate the total and phosphorylated protein content of translational machinery in pancreatic beta cells as this might unveil some of the mechanisms by which THs regulate protein, and particularly insulin, synthesis.
Materials and Methods
Materials
T3, Triton X-100, and
Cell culture
The rat INS-1E insulinoma cell line was generously provided by Prof. Dr. Meire Sogayar (University of São Paulo, São Paulo, Brazil) and cultured in RPMI 1640 with 11 mM glucose, supplemented with 10% FBS, 10 mM HEPES, 1 mM sodium pyruvate, 2 mM
Total protein determination and electrophoresis
Cells were lysed in buffer containing 100 mM Tris (pH 7.5), 10 mM EDTA (pH 7.0), 35 mM sodium dodecyl sulfate (SDS), 100 mM fluoride, 17 mM pyrophosphate, and 10 mM orthovanadate. The homogenate was centrifuged at 12,000 rpm for 40 minutes at 4°C, and the supernatant was used for total protein determination (20).
One hundred micrograms of protein from each sample were treated with Laemmli buffer (21), submitted to electrophoresis in SDS-–polyacrylamide gel electrophoresis, and transferred to a nitrocellulose membrane by electroblotting, for 60 minutes at 15 V. Nonspecific protein binding to the membrane was reduced by preincubation with blocking buffer (5% nonfat dry milk, 2.7 mM KCl, 137 mM NaCl, 8 mM NaHPO4, 1.4 mM KPO4, and 0.1% Tween 20), overnight, at 4°C. The membrane was incubated with the specific primary antibodies in blocking buffer, for at least 3 hours at room temperature (RT), followed by incubation with the appropriate secondary peroxidase-conjugated antibody (1:5000), in 2.7 mM KCl, 137 mM NaCl, 8 mM NaHPO4, 1.4 mM KPO4, and 0.1% Tween 20 buffer, for 90 minutes at RT. The antibodies and acrylamide gel concentration are described in Table 1. After washing the membrane, band detection was carried out by using the ECL kit.
HRP, horseradish peroxidase; SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis; eIF, eukaryotic initiation factor; p-, phosphorylated; 4E-BP, eIF4e binding protein.
Blots were analyzed by scanning densitometry and quantified using the Image Master-1D-Pharmacia Biotech SW software (Pharmacia Biotech, Uppsala, Sweden). Membranes were also incubated with monoclonal anti-α-actin antibody, which was used as an internal control. The results were expressed as arbitrary units.
Radioimmunoassay for insulin determination
INS-1E cells culture medium was collected from each flask, and insulin concentration was measured by radioimmunoassay, as previously described (22).
Statistical analysis
The data were expressed as the mean±standard error of the mean and subjected to analysis of variance, followed by the Student-Newman-Keuls post-test. Differences were considered significant at p<0.05.
Results
T3 increases insulin protein expression
The insulin content was determined by Western blotting and radioimmunoassay (RIA) analysis, as described. With regard to the insulin stored in beta cells, an increase in insulin content after T3 treatment was observed as compared with the control cells (Fig. 1A,B). T3 treatment also increased insulin levels in the cell-culture medium, as shown by the RIA analysis (Fig. 1C).

Triiodo-
T3 induces phosphorylation of p70S6K
Total and phosphorylated p70S6K were evaluated. Total p70S6K remained unchanged in the T3-treated cells, when compared with the control cells; however, the phosphorylated p70S6K amount increased in the T3-treated group. These data are shown in Figure 2.
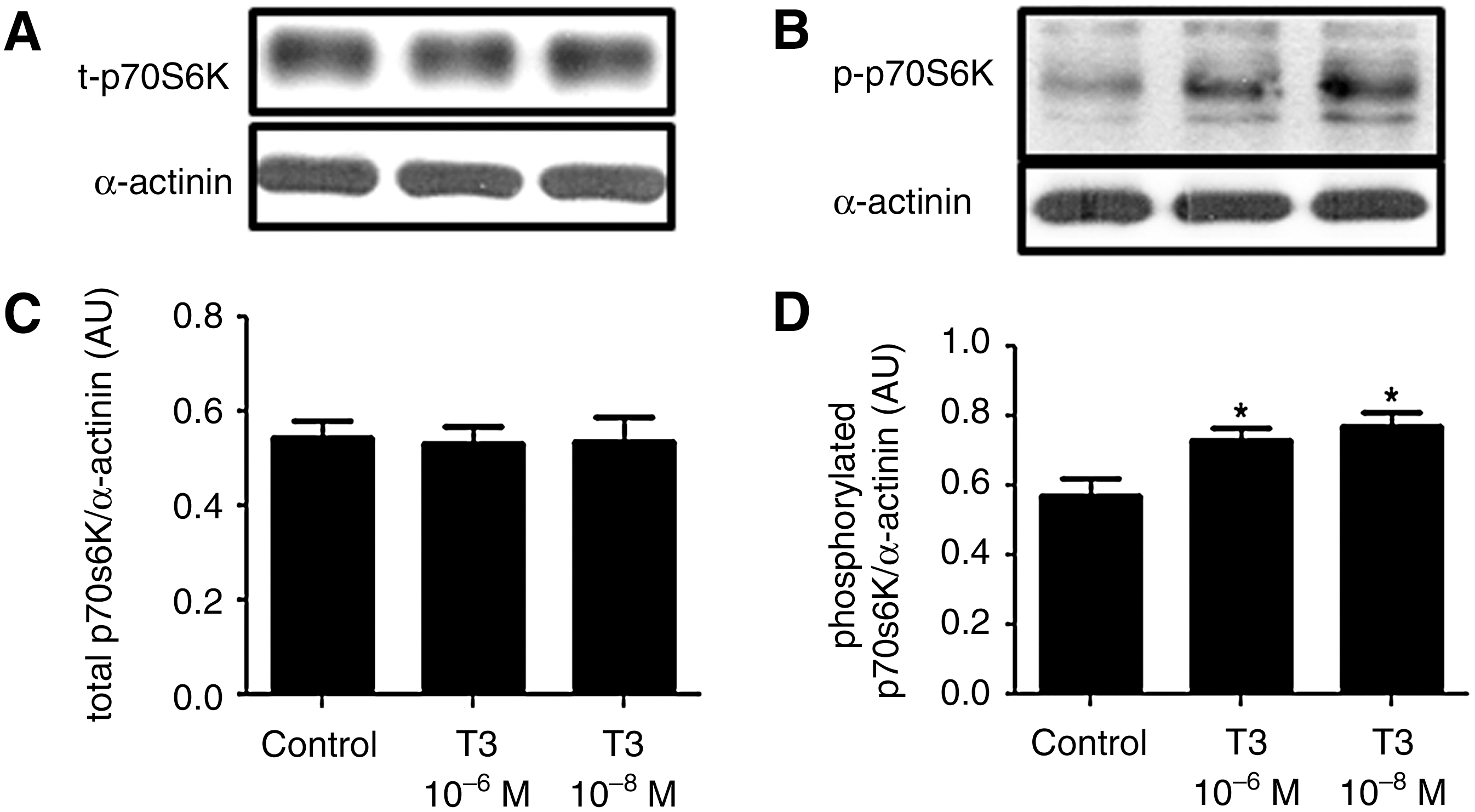
T3 induces phosphorylation of p70S6K. Representative blots of total
T3 increases eIF2 protein expression and decreases the ratio between phosphorylated and total eIF2
The amount of both total and phosphorylated eIF2 was evaluated. T3 treatment led to an increase of total eIF2 content in beta cells compared with the control group (Fig. 3A,C). On the other hand, the phosphorylated eIF2 content remained unaltered in T3-treated cells versus the control cells (Fig. 3B,D); however, the ratio between phosphorylated and total eIF2 in the T3-treated group was shown to be decreased versus the control (Fig. 3E).
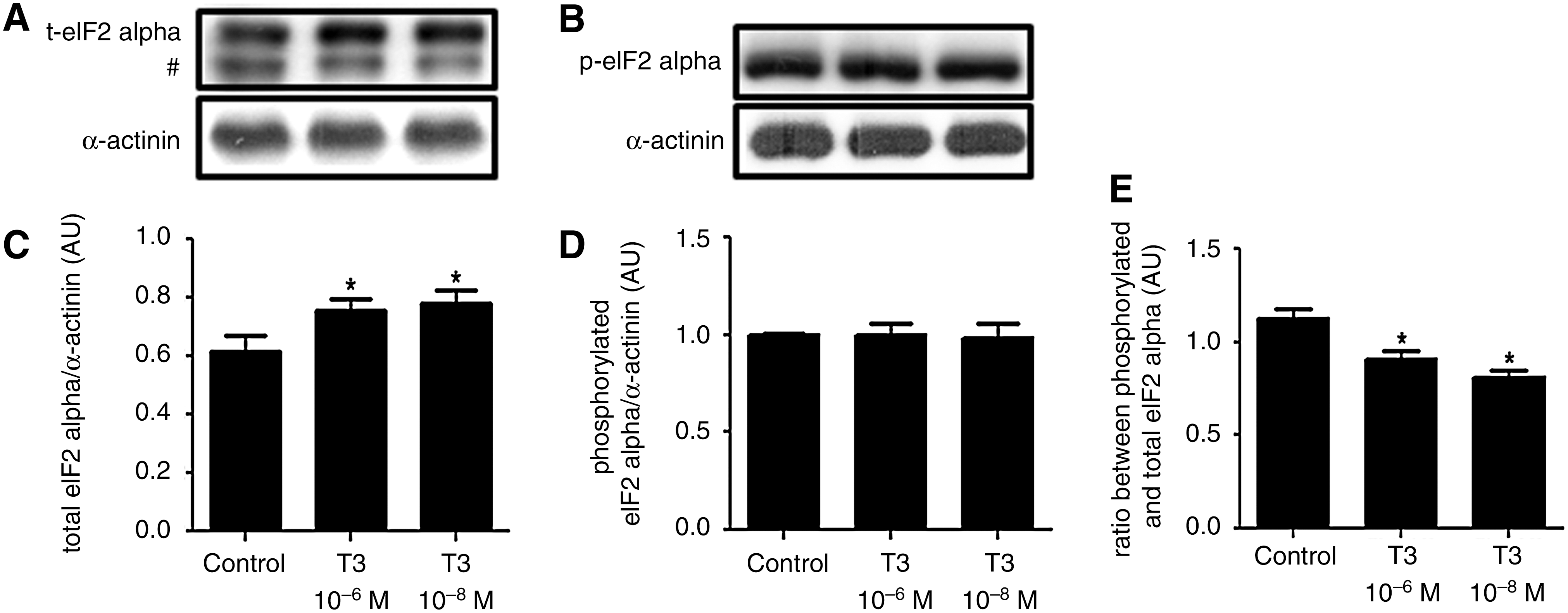
T3 increases eukaryotic initiation factor 2 (eIF2) protein expression and decreases the ratio between phosphorylated and total eIF2. Representative blots of total
T3 does not affect the total and phosphorylated 4E-BP protein content
Cells that had been incubated with T3 did not show alterations in either total or phosphorylated 4E-BP amount when compared with the control cells. These data are shown in Figure 4.
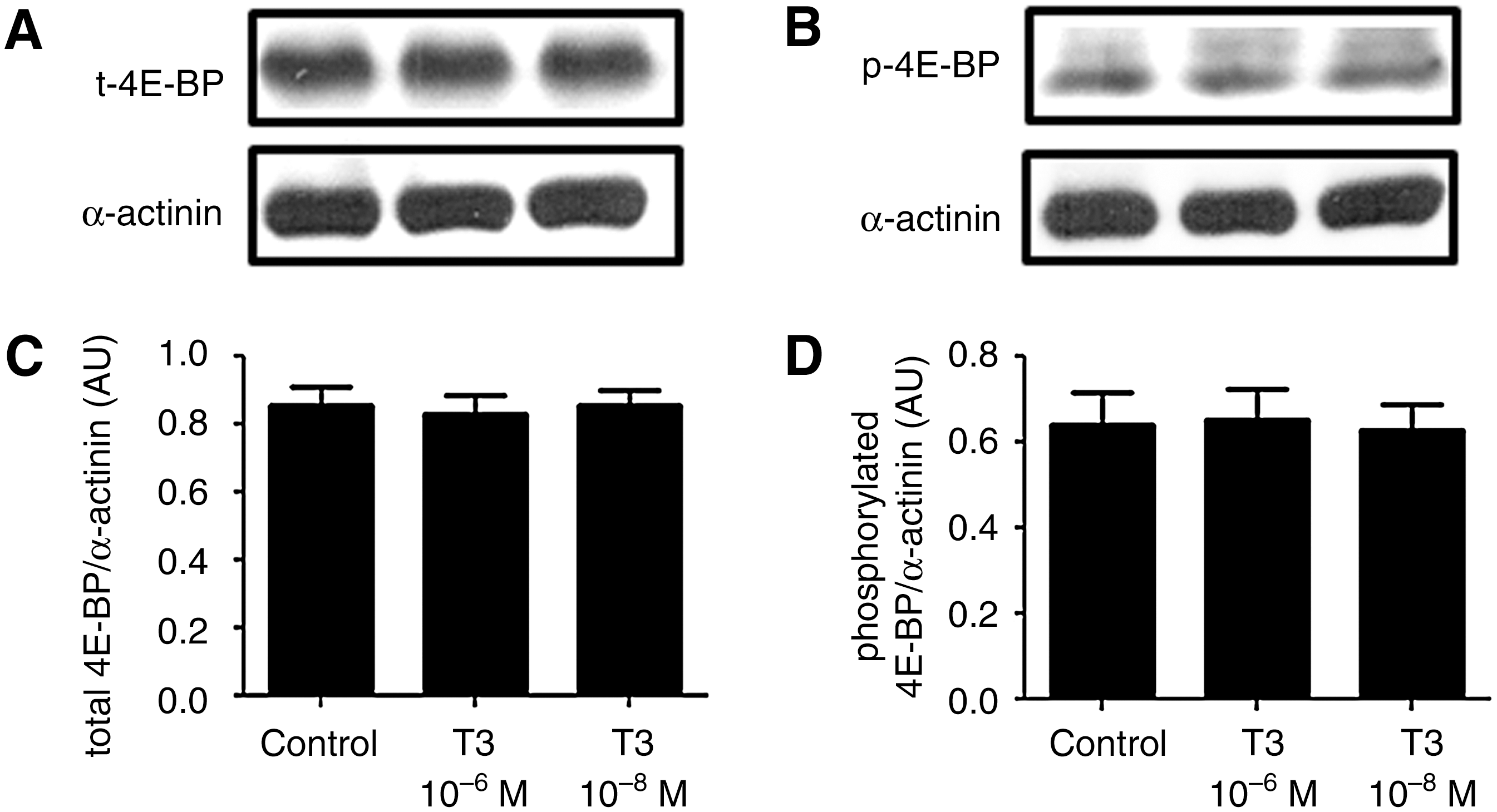
T3 does not regulate eIF4E binding protein (4E-BP) expression and phosphorylation. Representative blots of total
T3 increases eEF1A and eIF5A protein expression
Compared with the control, the T3-treated cells showed an increase in both eEF1A and eIF5A translational factors, which are committed to the elongation phase of protein synthesis. These data are shown in Figure 5.
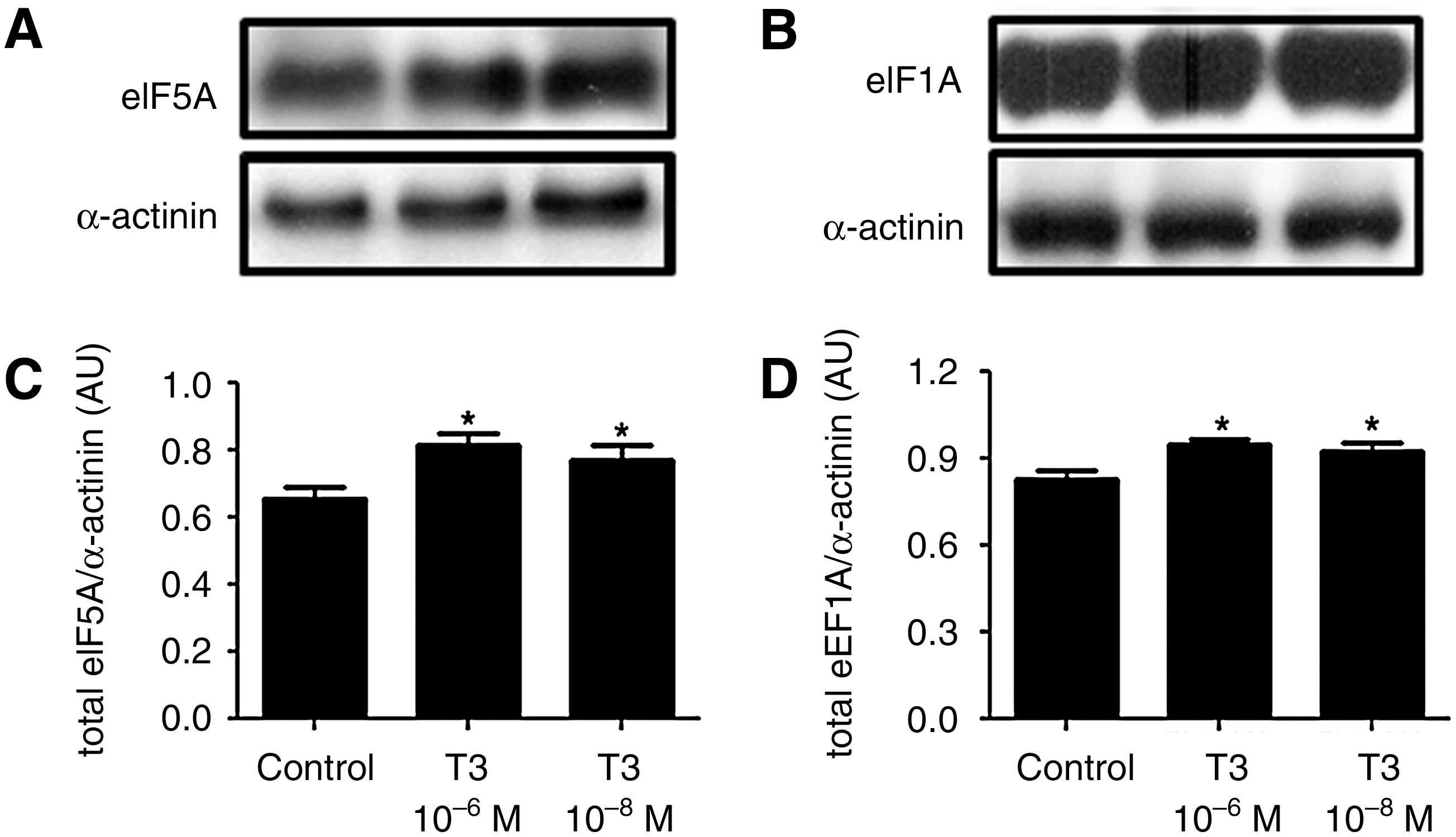
T3 increases eukaryotic elongation factor 1 alpha (eEF1A) and eIF5A expression. Representative blots of total eEF1A
Discussion
This study focuses on the modulation of the expression and activities of translational factors by TH (T3); this may relate to the stimulatory T3 action on insulin production. Insulin is the major product synthesized by pancreatic beta cells and one of the most important hormones involved in the regulation of carbohydrate metabolism (24). Insulin synthesis and secretion are regulated by several factors, including metabolic, neural, and hormonal ones. With regard to the latter, we have shown that T3 is one of the hormonal factors that is capable of regulating insulin production and secretion, as insulin content inside beta cells, as well as in the culture medium, increases after T3 administration.
These data indicate a clear stimulatory effect of THs on insulin synthesis. This has not yet been properly characterized in other tissues and cells (25,26). It is known that T3 induces the activation of p70S6K, thus stimulating protein synthesis in many tissues and cells (5,27). This led us to postulate that proteins from translational machinery may be targets for the regulatory action of THs on protein synthesis.
Indeed, we found that T3 modulates p70S6K activity in beta cells; evidence for this was the increase in the amount of phosphorylated p70S6K, an effect that apparently does not require new protein synthesis, as the total p70S6K content remained unchanged. It is worth noting that p70S6K activates the ribosomal S6 protein by phosphorylation, enhancing the decoding of several mRNA species into proteins (6); this may explain, in part, the increase in insulin expression observed in these cells.
The translation of transcripts is a complex process involving several proteins. One of them is eIF2, which is essential for the initiation of mRNA translation. In this study, we found that eIF2 expression is under the control of THs, as T3 administration led to an increase in its content. This may accelerate insulin synthesis, as more ternary complexes are formed, allowing more transcripts to be directed to ribosomes and enabling translation into protein. Moreover, T3 did not affect eIF2 phosphorylation. This is known to reduce the activity of eIF2 and, as a consequence, protein synthesis (28). Therefore, it seems that the decreased ratio of phosphorylated eIF2 to total eIF2 as observed in T3-treated cells, compared with control cells, reinforces the stimulatory T3 effect on insulin synthesis.
Usually, the stimulation of p70S6K activities is accompanied by phosphorylation of 4E-BPs. This process leads to the inhibition of 4E-BPs and increases protein synthesis, due to the consequent release of eIF4E, which allows its interaction with specific regions of the transcripts, an important step in mRNA translation. Despite the increase in p70S6K phosphorylation, T3 treatment was not associated with a change in either 4E-BP phosphorylation or its total protein levels, indicating that, in this beta-cell line, 4E-BPs do not mediate the effect of THs on insulin expression.
Furthermore, T3 also modulates other classes of molecules that are involved in polypeptide chain elongation. One of these proteins is the eEF1A, which binds to the aminoacyl-tRNA and transports it to ribosomes, where the tRNA anticodon's triplet sequence can base-pair to one or more codons, starting the elongation stage of polypeptide chain formation (12,13). This phase is crucial to protein synthesis, and seems to be regulated by THs, as eEF1A content significantly increased after T3 treatment. Moreover, T3 increased the expression of the elongation factor eIF5A, which is thought to stimulate the ribosome to move on the mRNA molecule, as previously shown for eEF2 (15,16). Thus, the increase in eEF1A and eIF5A expression reinforces the concept that THs act on insulin production at the elongation phase, which is supported by the increase in the amount of amino acids addressed to ribosomes and the acceleration of the ribosome movement on the mRNA molecule.
In summary, the data presented here lead us to conclude that the stimulatory effect of T3 on insulin expression depends, at least in part, on activation of protein synthesis, as the expression of translational factors, as well as p70S6K phosphorylation, was modulated by T3. In addition, this study helps us understand some of the molecular mechanisms whereby T3 regulates protein synthesis. It should be noted that, even though we have focused our studies on insulin synthesis, we cannot exclude the possibility that other proteins in these cells we studied might also be similarly altered by T3 treatment.
Footnotes
Acknowledgments
This work was supported by a grant from the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP: 08/56446-9). The authors thank Leonice Lourenço Poyares for the excellent technical assistance rendered.
Disclosure Statement
The authors declare that no competing financial interests exist.