
Abstract
Background:
Decreased serum leptin has been proposed as a critical signal initiating the neuroendocrine response to fasting. Leptin administration partially reverses the fasting-induced suppression of the hypothalamus-pituitary-thyroid axis at the central level. It is, however, unknown to what extent leptin affects peripheral thyroid hormone metabolism. The aim of this study was to evaluate the effect of leptin administration on starvation-induced alterations of peripheral thyroid hormone metabolism in mice.
Methods:
Three types of experiments were performed: (i) mice were fasted for 24 hours while leptin was administered twice (at 0 and 8 hours, 1 μg/g body weight [BW]), (ii) mice were fasted for 24 hours and, subsequently, leptin was given once at 24 hours (killed at 28 and 32 hours), and (iii) mice were fasted for 48 hours. All groups had appropriate controls. Serum triiodothyronine and thyroxine, liver type 1 deiodinase (D1), type 3 deiodinase (D3), thyroid hormone receptor (TR)β1, TRα1 and α2 mRNA expression, and liver D1 and D3 activity were measured.
Results:
Twenty-four hours of fasting decreased liver TRβ1 mRNA expression, while liver TRα1, TRα2, and D1 mRNA expression and activity did not change. In contrast, 24 hours of fasting increased liver D3 mRNA. Leptin administration after fasting restored liver D3 expression, while serum thyroid hormone levels and liver TRβ1 expression remained low.
Conclusion:
Leptin administration selectively restores starvation-induced increased hepatic D3 expression independently of serum thyroid hormone concentrations. The present study shows that fasting-induced changes in mRNA expression of genes involved in hepatic hormone metabolism are influenced not only by decreased serum thyroid hormone levels but also by serum leptin.
Introduction
In humans, the effects of fasting include changes in peripheral thyroid hormone metabolism, such as decreased conversion of thyroxine (T4) into triiodothyronine (T3), decreased reverse T3 (rT3) degradation, and serum thyroglobulin concentrations (4,5). In rodents, liver type 1 deiodinase (D1) decreases during fasting probably as a result of decreased serum T3 levels (6), while maximal hepatic binding capacity of the thyroid hormone receptor (TR) decreases (7). Together, the observed changes suggest decreased thyroid hormone action in the liver during fasting, which may help in decreasing energy expenditure.
It is unknown at present whether leptin modulates the fasting induced alterations in hepatic thyroid hormone metabolism. It has been shown that leptin administration during fasting partly prevents the fasting-induced changes in serum corticosterone and T4 concentrations (2), and a more recent study by Araujo et al. reported that leptin administration during restricted feeding in rats partly restores D1 activity in liver and kidney, but not in the thyroid gland (8).
Leptin exerts its action via the long form of the leptin receptor (Ob-RL), a member of the class 1 family of cytokine receptors. The Ob-RL is predominantly expressed not only in specific nuclei of the hypothalamus but also in the liver and other peripheral organs, making a functional role of leptin in peripheral organs during fasting plausible (9 –11).
At this stage, it is largely unknown to what extent leptin affects the fasting-induced changes in hepatic thyroid hormone metabolism. The aim of the present study was to evaluate fasting-induced alterations in thyroid hormone metabolism in the mouse liver and to study the effect of leptin administration on these changes. Liver thyroid hormone metabolism was characterized by measuring TRβ1, TRα1, and TRα2 mRNA expression, as well as D1 and type 3 deiodinase (D3) mRNA expression and activity.
Materials and Methods
Animals
Female Balb/c mice (Harlan Sprague Dawley, Horst, The Netherlands) weighing ∼20 g were used at 6–12 weeks of age. The mice were kept in 12 hours of light/12 hours of darkness cycles (lights on from 07:00 to 19:00 hours), in a temperature-controlled room (22°C). Food and water were available ad libitum. A week before and during the experiment, the mice were housed in groups according to the experimental setup. We performed two types of experiments that were approved by the Local Animal Welfare Committee of the Academic Medical Center, University of Amsterdam. An additional starvation experiment (exp 2) was performed with male B6129S6F1 mice (∼22 g and 12 weeks of age) at the Gorlaeus Laboratory Leiden. The animals were kept under a 12/12 light/darkness cycle (light on from 07:00 to 19:00 hours) in a temperature-controlled room (23°C). Water and food were available ad libitum. This experiment was approved by the Local Animal Welfare Committee of the University of Leiden, the Netherlands.
Experimental protocols
Experiment 1: Two groups of mice (n=6/group) were fasted for 24 hours. One group received 20 mg leptin/0.5 mL 0.01 M NaHCO3 intraperitoneally (recombinant mouse leptin, obtained from Dr. Parlow, Harbor-UCLA Medical Center, Torrance, CA). Leptin was administered twice (at 09:00 hours [t=0 hours] and 17:00 hours [t=8 hours]) in this period. The other group received the vehicle at the same time points. Both groups were compared with control mice (n=6) that received vehicle and had free access to food. Experiment 2: One group of mice (n=12/group) was fasted for 48 hours, and one group had free access to food. Experiment 3: Two groups of mice (n=6/group) were fasted for 24 hours and, subsequently, leptin was given at 09:00 hours (t=24 hours, intraperitoneally 20 mg/0.5 mL 0.01 M NaHCO3) while the mice continued fasting. One group was killed at 28 hours, and the other was killed at 32 hours. Both groups were compared with the fed control mice that also received leptin at 24 hours and were killed at 28 and 32 hours.
Due to the diurnal variation of thyroid hormone-related genes, all the experiments were performed using the same time schedule starting at 09:00 hours (t=0). In all experiments, mice were anesthetized with isoflurane, blood was taken by cardiac puncture, and then, the mice were killed by cervical dislocation. Serum was stored at −20°C until it was analyzed. The liver was isolated and stored immediately in liquid nitrogen. We measured suppressor of cytokine signaling (SOCS) mRNA expression in order to check our experimental set up, because SOCS3 is activated by leptin (12).
RNA isolation and RT-polymerase chain reaction
Liver mRNA was isolated on the Magna Pure (Roche Molecular Biochemicals, Mannheim, Germany) using the Magna Pure LC mRNA tissue kit and ∼10 mg of liver tissue. The protocol and buffers supplied with the corresponding kit were applied. cDNA synthesis was performed using the First Strand cDNA Synthesis Kit for reverse transcriptase (RT)–polymerase chain reaction (PCR) with oligo d(T) primers (Roche Molecular Biochemicals). Real-time PCR was performed for quantitation of hypoxanthine phosphoribosyl transferase (HPRT), D1, D3, TRβ1, TRα1, and TRα2 mRNAs using the Lightcycler480 and Lightcycler480SybrGreen I Master mix (Roche Molecular Biochemicals). Primer pairs for HPRT, D1, D3, TRβ1, TRα1, and TRα2 were previously described (13 –15), We designed primer pairs for suppressor of cytokine signaling (SOCS3): SOCS3-F 5′-GTG CGC CAT GGT CAC CCA CA-3′, SOCS3-R- 5′-GGT CTT GAG GCG CAG GCT GG-3′, annealing temperature 68°C. Primers were intron spanning or genomic DNA contamination was tested using a cDNA synthesis reaction without the addition of Reverse Transcriptase. Quantification was performed using the LinReg software (16). The mean of the efficiency was calculated for each assay, and samples that had a difference greater than 0.05 of the efficiency mean value were not taken into account (0%–5%). Samples were corrected for their mRNA content using HPRT as a housekeeping gene.
Deiodinase activity
Liver D1 and D3 activities were determined as described earlier (17). In short, frozen tissue samples were homogenized on ice in 10 volumes of phosphate-EDTA (PE) buffer (0.1M sodium phosphate, 2 mM EDTA pH 7.2) using a Polytron (Kinematica, Luzern, Switzerland). 20 mM dithiotreitol was added to the PE buffer for D1 activity measurement and 50 mM, for D3. Homogenates were snap frozen in aliquots and stored at −80°C until further analysis. Protein concentration was measured with the Bio-Rad protein assay using bovine serum albumin (BSA) as the standard following the manufacturer's instructions.
D1
Liver D1 activity was measured in duplicate, using 50 μL 100–500 times diluted homogenate incubated in a final volume of 0.1 mL with 0.1 μM rT3 with the addition of ∼1×105 cpm [3′5′-125I]rT3 in PED10 buffer (0.1 M sodium phosphate, 2 mM EDTA, and 10 mM dithiothreitol pH 7.2). Reactions were stopped by adding 0.1 mL of 5% BSA on ice. The protein-bound iodothyronines were precipitated by the addition of 0.5 mL 10% (w/v) trichloroacetic acid. After centrifugation, 125I− was separated from the supernatant by chromatography on Sephadex LH-20 columns with a bed volume of 0.25 mL, equilibrated, and eluted with 0.1 M HCl. Released 125I was counted using the Packard Cobra Auto-Gamma Counting System (Canberra Packard, Zürich, Switzerland) in the eluate. D1 activity was expressed as 125I pmol released per minute per mg liver protein.
D3
D3 activity was measured by duplicate incubations of homogenates (∼100 μg protein) for 120 minutes at 37°C with 1 nM [3′-125I]T3 (200,000 cpm) in a final volume of 0.15 mL PED50 buffer in the absence or presence of 500 nM unlabeled T3 to inhibit D3 activity. Reactions were stopped by the addition of 0.15 mL ice-cold ethanol. After centrifugation, 0.125 mL of the supernatant was added to 0.125 mL 0.02 M ammonium acetate (pH 4), and 0.1 mL of the mixture was applied to the 4.6×250 mm Symmetry C18 column connected to an Alliance HPLC system (Waters, Etten-Leur, The Netherlands). The column was eluted with a linear gradient of acetonitrile (28%–42% in 15 minutes) in 0.02 M ammonium acetate (pH 4.0) at a flow of 1.2 mL/minute. The activity of T3 and 3,3′-diiodothyronine (T2) in the eluate was measured online using a Radiomatic Z-500 flow scintillation detector (Packard, Meriden, CT). Since incubation with 500 nM T3 saturates D3, the activity measured with the incubation with 1 nM T3 minus the incubation with 500 nM T3 represents true D3 activity. D3 activity was expressed as fmol generated 3,3′T2 per hour per mg tissue.
Thyroid hormone levels
Serum T3 and T4 were measured with in-house radio immunoassays (RIAs) (18). All samples of one experiment were measured within the same run (intra-assay variability T3: 3.6% and T4: 6.6%).
Statistical analysis
Data are presented as mean±standard error of the mean. Variations between fasted mice, fasted mice treated with leptin, and control-fed mice (exp 1) were evaluated by analysis of variance (ANOVA) with one grouping factor (treatment) and—if significant—followed by Tukey's test for pair-wise comparisons. Variations between fasted mice treated with leptin and control-fed mice treated with leptin (exp 3) were evaluated by ANOVA with two grouping factors (time and treatment) and their interaction followed by Tukey's test for pair-wise comparisons. p-values in the figures represent the statistical significance tested by ANOVA, and symbols represent the differences between groups. In case of abnormal distribution of the data, data were rank transformed before the ANOVA. All analyses were carried out in SPSS 11.5.1 (SPSS, Inc., Chicago, IL). p-values<0.05 were considered statistically significant.
Results
Fasting-induced alterations in liver thyroid hormone metabolism and the effect of leptin administration during fasting
SOCS3 mRNA expression was measured in the liver in an attempt to address the question whether leptin has a direct effect on the liver (12), and, indeed, leptin administration during fasting resulted in a marked increase of SOCS3 mRNA expression (SOCS3 relative expression, fasted: 0.09±0.01 vs. fasted/leptin: 0.26±0.05, p<0.05).
Fasting for 24 hours resulted in significantly decreased serum T3 (p<0.01) and T4 (p<0.01) levels. T3 and T4 concentrations were slightly but significantly higher in the leptin treated group compared with starved mice, but still significantly lower compared with fed control mice (data not shown).
Liver TRβ1 mRNA expression decreased (p<0.01), while liver TRα1 and α2 mRNA expression did not change during fasting. Leptin administration was given twice during the 24 hours of fasting in order to evaluate whether fasting-induced changes can be prevented by leptin administration. Leptin administration did not prevent the fasting-induced decrease of liver TRβ1 mRNA expression, while it decreased liver TRα1 mRNA expression (p<0.05). TRα2 mRNA expression was not altered by any of the experimental conditions (Fig. 1).
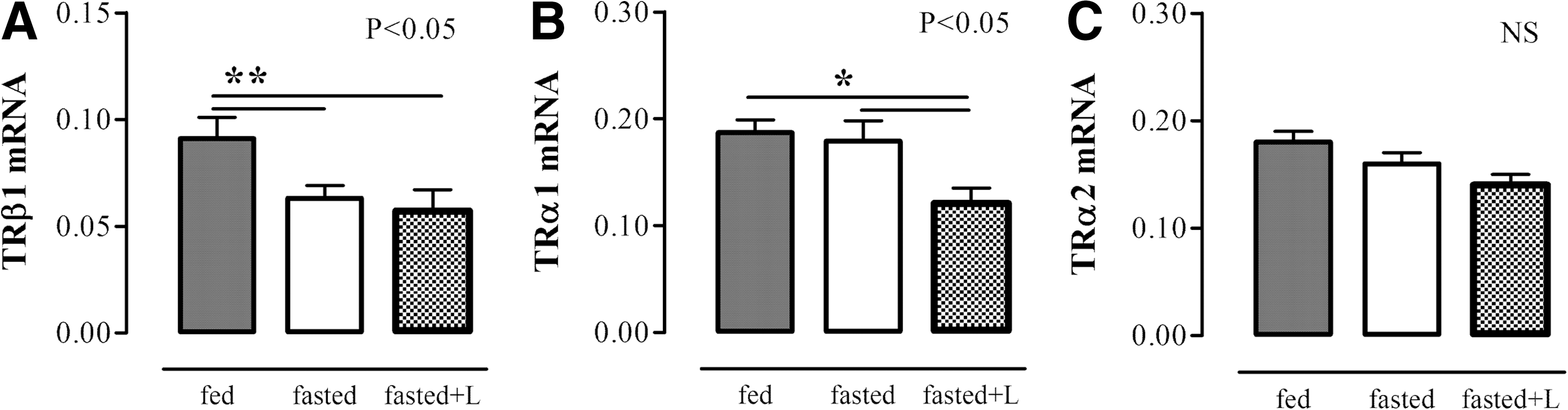
Liver TRβ1 mRNA
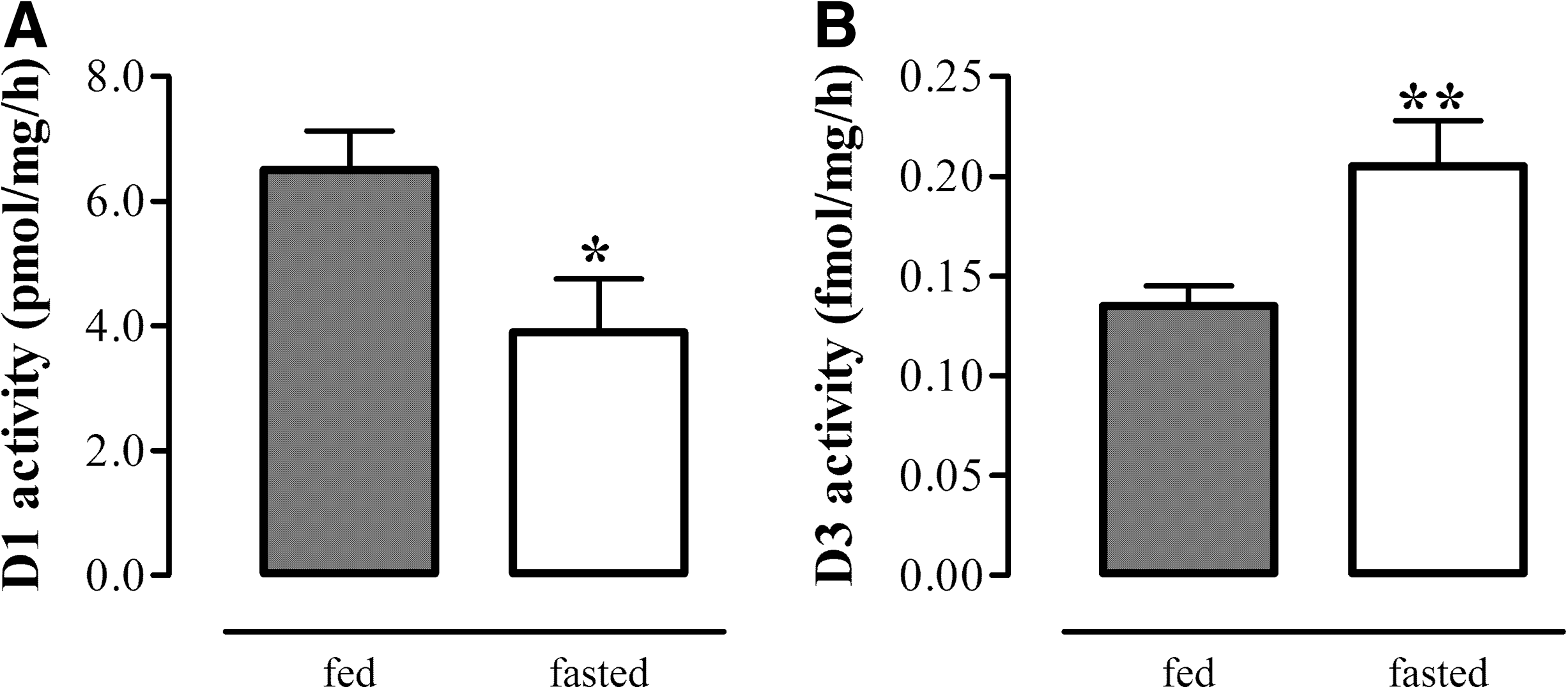
Liver D1 mRNA expression and activity did not change after 24 hours of fasting. By contrast, liver D3 mRNA expression increased significantly compared with fed control mice (p<0.01), and D3 activity tended to increase (p=0.06) (Fig. 2). Liver D1 mRNA expression and activity were not affected by leptin treatment during the 24 hours of fasting. By contrast, the fasting-induced increase in liver D3 mRNA expression was partly restored by leptin administration (p<0.05), although liver D3 activity was still high in the leptin-treated fasted mice (Fig. 2). A prolonged period of fasting (48 hours) resulted in significantly decreased liver D1 and increased liver D3 activity levels (Fig. 3). Fasting for 48 hours also resulted in significantly decreased liver TRβ1 mRNA expression, while liver TRα1 and TRα2 mRNA expression was not affected (data not shown).
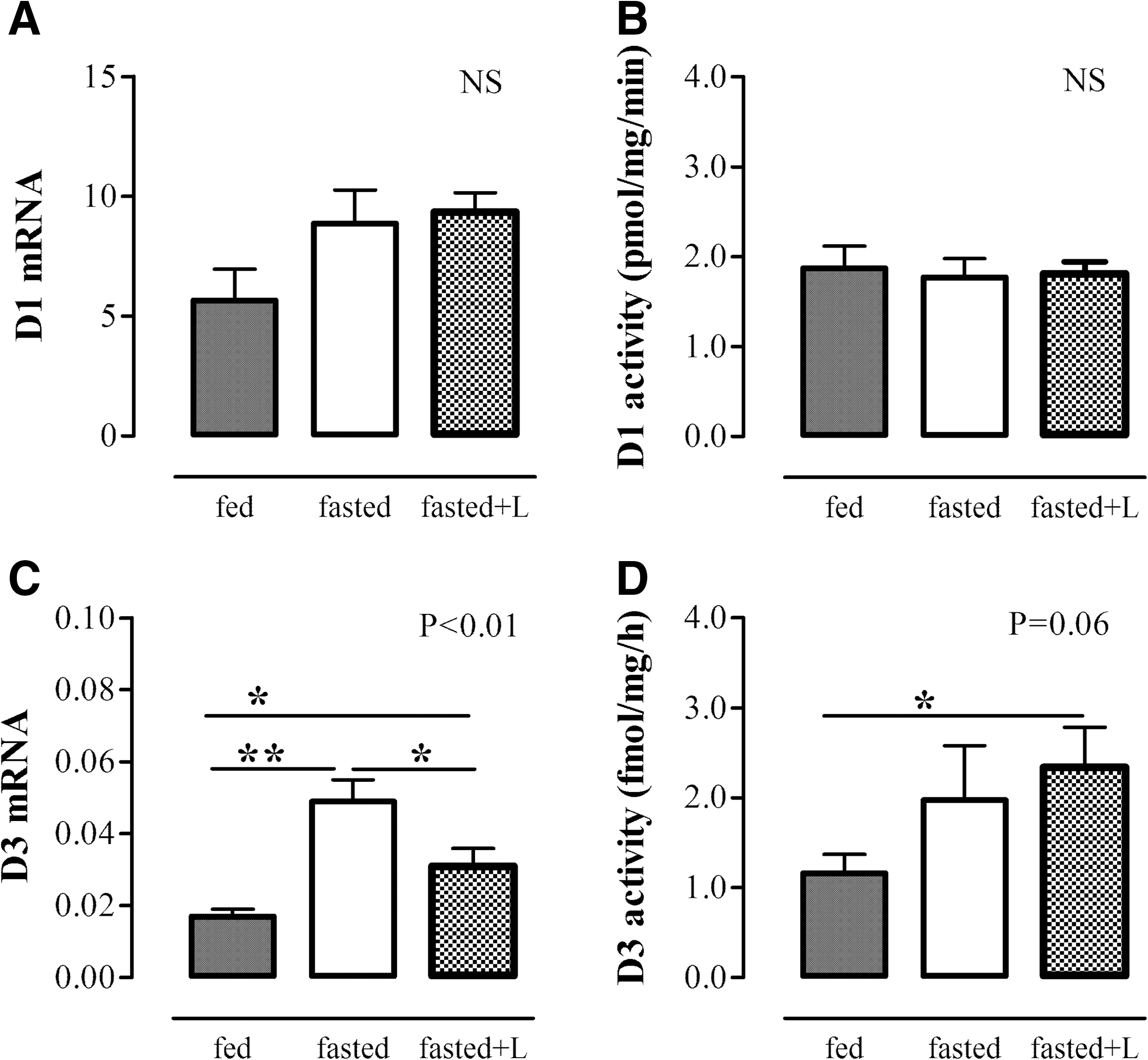
Liver D1 mRNA expression
Liver D1 activity
Effect of leptin administration on liver thyroid hormone metabolism after fasting
Liver SOCS3 mRNA expression was higher in leptin-treated mice at 28 hours (0.21±0.03 [arbitrary units], p<0.05) compared with those at 24 hours (0.09±0.01).
The fasting-induced decrease of serum T3 and T4 at 28 and 32 hours, respectively, was not affected by leptin treatment at 24 hours (data not shown).
Leptin was administered after a fasting period of 24 hours in order to evaluate whether fasting-induced alterations can be restored by leptin administration. Mice remained fasted after leptin administration and were compared with fed mice that received a single injection of leptin. We determined liver mRNA expression 4 and 8 hours after administration, because leptin serum levels declined within 8 hours after the injection to 25% of the fed state (19). Mice were fasted for 28 and 32 hours, respectively. Due to the diurnal variation of thyroid hormone-related genes, fasted leptin-treated groups were compared with fed leptin-treated groups at the same time point. TR expression and deiodinase expression and activity levels measured in the fed group varied and reflected the diurnal rhythm throughout the day. Fasting resulted in decreased TRβ1 mRNA expression compared with fed mice. Leptin administration at 24 hours did not affect the fasting-induced decrease of TRβ1 mRNA at 28 and 32 hours; it remained low compared with the fed leptin-treated mice (ANOVA, p<0.01, Fig. 4). Liver TRα1 and TRα2 mRNA expression were not different between the fasted and fed mice treated with leptin (Fig. 4).
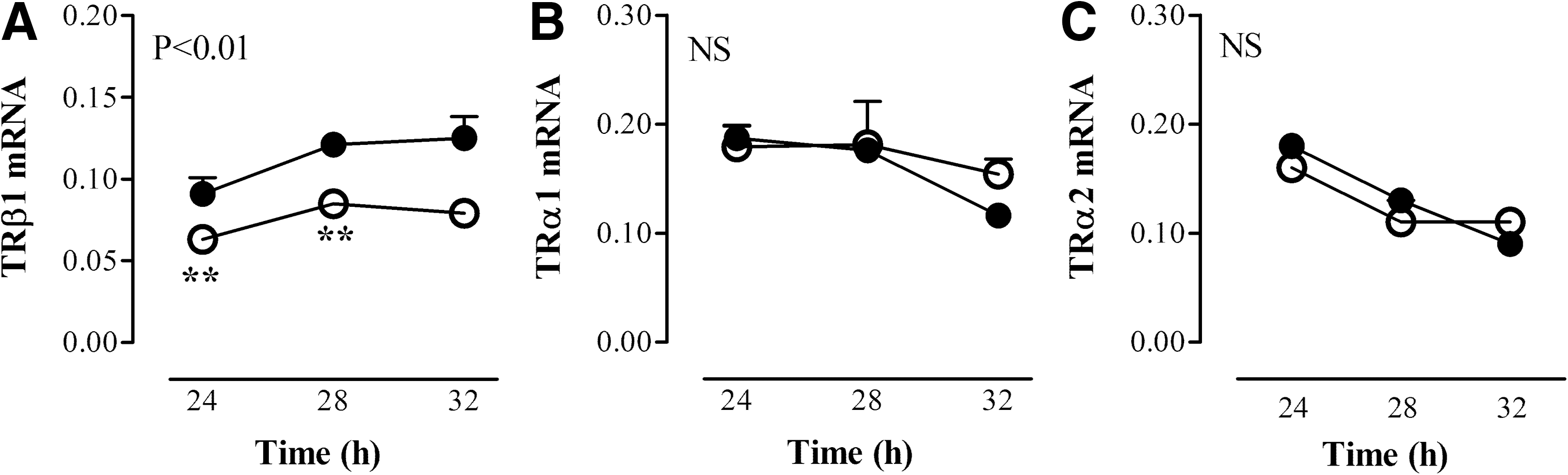
Liver TRβ1 mRNA
Liver D1 mRNA expression was significantly different in fasted mice that received a single dose of leptin compared with the fed mice that received leptin during the same period (ANOVA, p i<0.01). Comparison between the groups showed that only a fasting period of 32 hours in combination with leptin was sufficient to lower liver D1 mRNA expression (p<0.01). Leptin administration to fasted mice did not restore liver D1 mRNA expression compared with the fed mice who received leptin. Moreover, liver D1 activity was completely unaffected in the fasted mice that received a single dose of leptin compared with the control mice (Fig. 5). In contrast, the fasting-induced increase of liver D3 mRNA expression was completely restored 4 and 8 hours after leptin injection (28 and 32 hours fasting, respectively) compared with the fed mice that received leptin (ANOVA, p<0.05). Liver D3 activity was still increased in fasted mice that had received a single dose of leptin 4 hours earlier, while it normalized to control levels 8 hours after leptin administration, indicating that the effect of leptin on D3 expression might be at the transcriptional level (Fig. 5).
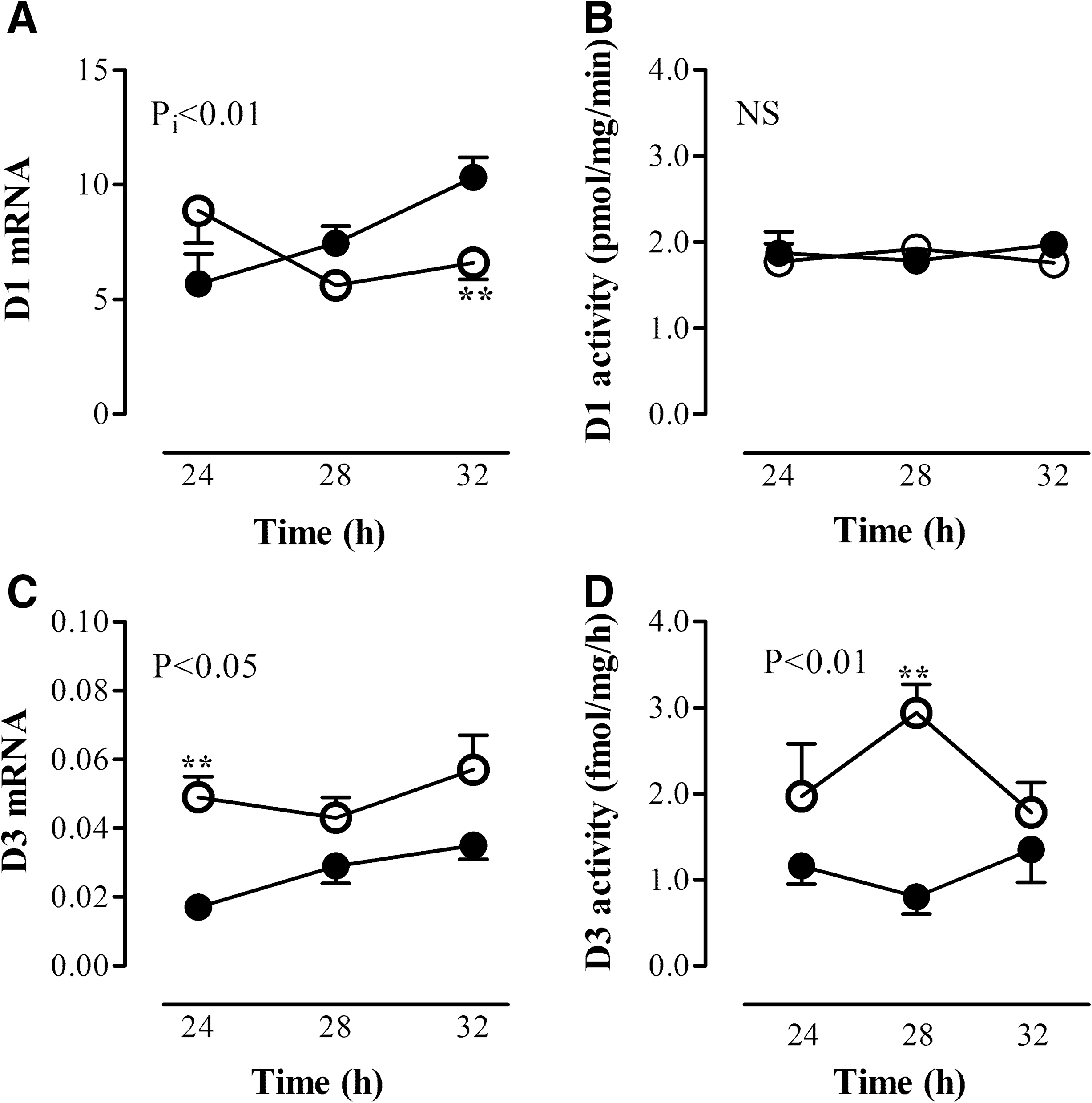
Liver D1 mRNA expression
Discussion
The aim of the present study was to evaluate fasting-induced alterations in thyroid hormone metabolism in the mouse liver as well as the effect of leptin administration on these changes. It has been suggested that the prime effect of fasting on the HPT axis in rodents might be a reduction of thyroidal secretion of serum thyroid hormone levels resulting from reduced stimulation by TSH (20). Although a prominent fall in serum thyroid hormone levels influences thyroid hormone-related gene expression by itself, the present study shows that fasting-induced changes in mRNA expression of genes involved in hepatic thyroid hormone metabolism are influenced not only by decreased serum thyroid hormone levels but also by serum leptin.
Leptin, an adipocyte-derived hormone, is predominantly involved in the neuroendocrine response to fasting by signaling energy shortage to the hypothalamus. In line with this, leptin administration during fasting may restore the fasting-induced decrease of preproTRH mRNA expression in the hypothalamic PVN (3). Ahima et al. showed that leptin during fasting is also capable of partly restoring the fasting-induced decrease in serum T4 levels (2). It is unknown at present whether these effects of leptin depend completely on the central part of the HPT-axis or whether leptin directly affects peripheral organs such as the thyroid gland or the liver.
We studied the influence of peripherally administered leptin on fasting-induced alterations in peripheral thyroid hormone metabolism by giving leptin twice during the fasting period, and by the administration of a single dose of leptin after the fasting period. We compared fasted mice with fed mice that also received leptin in order to control for leptin-induced effects on metabolism irrespective of fasting. We have shown earlier that the experimental design (time-course, moment, and dose of leptin) can be critical, because a single injection of leptin after 24 hours of fasting is not sufficient to restore the fasting-induced decrease in pituitary type 2 deiodinase (D2) and TRβ2 mRNA expression within 4 hours, while leptin given during the fasting period completely prevents the observed changes (19). In the present study, we measured liver SOCS3 mRNA expression in order to control for the experimental setup as leptin activates SOCS3 mRNA expression (12). Indeed, SOCS3 mRNA expression increased in the liver on leptin administration given both during and after the fasting period. From these observations, we concluded that our experimental approach was adequate to study the role of leptin in liver thyroid hormone metabolism, although we cannot fully exclude the effects of leptin-induced changes at the central level on liver thyroid hormone metabolism.
Serum thyroid hormone levels were markedly decreased during fasting, and we observed reduced TRβ1 mRNA expression in the liver of mice fasted for 24 hours, while TRα1 and TRα2 mRNA expression did not change. Liver TR expression during fasting has not been extensively studied so far. Oppenheimer and Schwartz showed many years ago that the maximal binding capacity of the TR in liver decreased during fasting (7) suggesting reduced T3-responsive gene expression, but it was unknown whether fasting affects TR mRNA expression as a result of decreased serum T3 levels. Sadow et al. studied the regulation of TR isoforms in the liver and heart and observed a decreased expression of TRβ1 mRNA in liver during T3 treatment (21). This study suggested negative regulation of the mouse THRβ gene by T3, implicating that fasting should result in increased expression of TRβ mRNA. However, since fasting decreased TRβ1 mRNA expression in the present study, other T3-independent mechanisms should be involved. A complex role of food intake in the regulation of TR isoform expression was suggested by an earlier study reporting that the diurnal 24 hour rhythm of liver TRβ1 was affected by food intake, while the diurnal variation of TRα1 and TRα2 was related to the activity of the biological clock (22). In the present study, leptin administration during fasting decreased TRα1 mRNA expression significantly compared with fed and fasted mice, which might represent a direct effect of leptin on the TRα1 mRNA expression. This concept is supported by the observation that an intraperitoneal leptin injection resulted in activation of the AP-1 pathway in mice (23), while other studies in hepatocytes showed that cytokine-induced signal transduction pathways are involved in basal TRα mRNA expression (15).
Liver deiodinase expression appeared to be differentially affected by 24 hours of fasting, as liver D1 did not change while liver D3 increased. D1 is a T3-responsive gene, and the fasting-induced decrease in serum thyroid hormone levels should result in decreased liver D1 mRNA expression and D1 activity. However, it has been previously shown that a period of 24 hours of fasting is not enough to induce significant decreased liver D1 mRNA expression in mice despite low serum thyroid hormone levels (24). That a longer period of fasting should result in lower D1 mRNA expression levels as shown earlier in rats (6) was supported by the present study showing that 48 hours of fasting in mice significantly decreases D1 activity. Leptin administration did not result in a normalization of serum thyroid hormone levels within 8 hours. In addition, leptin administration did not normalize liver D1 mRNA in fasted mice to the levels observed in fed mice, which is in contrast to the results observed by Araujo et al. (8). Araujo et al. postulated that leptin is able to normalize decreased liver D1 after food restriction (8). The increase in serum T3 on leptin treatment in their experimental setup, however, makes it difficult to distinguish between direct effects of T3 and leptin on liver D1 expression.
To our surprise, liver D3 mRNA expression increased after 24 hours of fasting, and D3 activity was significantly increased after 48 hours of fasting, making it plausible that fasting affects D3 expression at the transcriptional level. D3 inactivates thyroid hormone and is thought to be involved in decreasing T3 local bioavailability (25). A recent study in chickens also showed that food restriction increases hepatic D3, but not D1 (26). D3 is transcriptionally stimulated by T3, but other regulatory pathways are also involved as shown in a variety of cell types (25). One of these pathways is the Raf/MEK/ERK pathway belonging to the MAPK cascade (27). Interestingly, leptin activates Jak2 and phosphorylates STAT3 and MAPK, respectively (28). Activation of MAPK signaling by leptin implicates that leptin might be able to increase D3 expression, as several studies report a positive effect of an activated MAPK pathway on D3 expression (25). However, the fasting-induced increase in D3 mRNA expression was completely abolished by leptin shortly after the injection and leptin administration during the fasting period partly prevented the fasting-induced increase in D3. This is in agreement with studies performed in a human liver cell line (HepG2 cells), showing that stimulation of HepG2 cells with an activator of the Raf/MEK/ERK cascade did not result in increased D3 mRNA expression or activity (29). Furthermore, we have shown in several animal models of illness that despite increased liver cytokine expression, implying induction of the MEK/ERK cascade (30), liver D3 does not increase (31,32). Recently, it has been shown in a rabbit model of NTI by Weekers et al. (33) that exogenous infusion of TRH and growth hormone releasing protein (GHRP)-2 reverses down-regulation of hepatic D1 activity and up-regulation of D3 activity. These data suggest that peripheral changes in liver D1 and D3 are under central control. Since leptin administration also reversed the fasting-induced changes in TRH in the PVN (19), we cannot exclude the possibility that leptin has an indirect effect on liver D3 expression. However, when we plotted liver D3 mRNA expression against TRH mRNA expression in PVN, there was no correlation between both expressions during fasting.
In conclusion, fasting results in profound changes in hepatic thyroid hormone metabolism characterized by decreased TRβ1 and D1 mRNA expression and increased D3 mRNA expression preceding increased D3 activity. The increase in D3 might be mediated by a decrease in leptin and be independent of serum thyroid hormone levels. Notably, low thyroid hormone levels might be responsible for the decrease in liver D1 mRNA expression and low liver TRβ1 mRNA expression, although it is unclear at present to what extent the THRβ gene is regulated by T3 in this setting. The overall result of these complex changes in liver thyroid hormone metabolism during fasting is likely to be the downregulation of local T3 hormone action, which is assumed to represent an energy-saving mechanism that is instrumental in times of food shortage.
Footnotes
Acknowledgments
The authors would like to thank J. Daalhuisen (Department of Experimental Internal Medicine, University of Amsterdam) and M. Platvoet-ter Schiphorst (Department of Endocrinology and Metabolism, University of Amsterdam) for expert biotechnical assistance and the staff of the University of Amsterdam's Laboratory of Endocrinology for measuring thyroid hormones.
Disclosure Statement
The authors have nothing to disclose.