
Abstract
Background:
Comprehension of the regulatory mechanism involved in the sodium iodide symporter (NIS) expression is of great relevance for thyroid cancer. In fact, restoration of NIS expression would be a strategy to treat undifferentiated thyroid cancer. Previous in vitro findings suggest that the cyclic AMP–response element (CRE) modulator (CREM) is involved in control of NIS expression. In this work, we examined the expression of CREM in a series of thyroid cancer tissues and its action on NIS promoter in human thyroid cancer cells.
Methods:
Expression of mRNA levels for CREM, PAX8 and NIS was measured by quantitative reverse transcriptase–polymerase chain reaction (RT-PCR) in 6 normal thyroid tissues, 22 papillary, 12 follicular and 4 anaplastic thyroid cancers. The effect of CREM on transcriptional activity of the NIS promoter was investigated by transient transfection of human thyroid cell lines.
Results:
Compared to normal tissues, NIS and PAX8 mRNA levels were significantly reduced in all types of thyroid cancer. As expected, the maximal decrease was detected in anaplastic thyroid cancer. Conversely, CREM mRNA levels were increased in all types of thyroid cancer, reaching statistical significance for follicular and anaplastic thyroid carcinoma (p=0.0157 and 0.0045, respectively). Transfection experiments showed an inhibitory effect of CREM on NIS promoter activity in various thyroid cancer cell lines.
Conclusions:
These data demonstrate that CREM expression is increased in thyroid cancer tissue and may play a role in the downregulation of NIS expression in thyroid cancer acting at the transcriptional level.
Introduction
Several factors participate in the regulation of NIS expression in a tissue-specific way (7,8). Analysis of human NIS has revealed many binding sites for transcription factors in the upstream region of the gene involved in its transcriptional regulation (9 –13). To our knowledge, however, in only few of them (AP2, SP1, PAX8) has a parallel analysis of their expression levels, or activities in thyroid tumor tissues, been documented (13,14). Particular attention has been focused on the activation of the NIS upstream enhancer (NUE) region (15), located between −9847 and −8968 in the human promoter, containing an essential PAX8 site and a cyclic AMP–response element (CRE)–like site, able to bind transcriptional complexes including elements of the CRE-binding protein (CREB) family (11). Among them, we have recently demonstrated an essential role played by a CRE modulator protein (CREM) in normal thyroid cells stimulated by thyrotropin or forskolin (16).
Based upon this knowledge, we hypothesized that the decrease of NIS expression in thyroid tumors could be caused by modification of expression of transcription factors regulating activity of the NUE region. Thus, in the present work, we examined the expression of CREM in a series of thyroid cancer tissues and its action on NIS promoter in human thyroid cancer cell lines.
Materials and Methods
Thyroid tumor tissues
A series of 38 thyroid cancer tissues, including 22 papillary thyroid cancers (PTC), 12 follicular thyroid cancers, (FTC) and 4 anaplastic thyroid cancers (ATC) were obtained at thyroidectomy from patients operated on for sporadic thyroid cancer at the University Hospital of Rome. These cases have been the subject of other analyses in previous reports by our group (17). The samples were snap-frozen in liquid nitrogen and stored at −80°C. All samples were reviewed by a single pathologist, who confirmed the diagnosis of thyroid cancer, identified the histological variant of the tumor, and excluded the presence of normal tissue in the specimen. The clinicopathological characteristics of the patients are described in Table 1. As normal tissues, six contralateral normal lobes of a different series of subjects undergoing thyroidectomy were utilized. The project was approved by the local ethical committee.
This item applies only to papillary thyroid cancer.
AJCC, American Joint Committee on Cancer; ATA, American Thyroid Association.
Quantitative reverse transcriptase–polymerase chain reaction
Quantitative polymerase chain reaction (PCR) analysis of the expression of the mRNA for NIS (NIS mRNA) was performed as previously described (18). Briefly, total RNA from cell lines was extracted with RNeasy protect mini kit (Qiagen, Hilden, Germany). One microgram of total RNA was reverse transcribed to single-strand cDNA using random exaprimers and 200 U MMLV reverse transcriptase (Invitrogen Corp., Carlsbad, CA) in a final volume of 20 μL at 42°C for 50 minutes followed by heating at 70°C for 15 minutes. Real-time PCRs were performed using the ABI Prism 7300 Sequence Detection System (Applied Biosystems, Foster City, CA). A 25 μL reaction mixture containing 5 μL cDNA template, 12.5 μL TaqMan Universal PCR master mix (Applied Biosystems), and 1.25 μL primer probe mixture was amplified using the following thermal cycler parameters: incubation at 50°C for 2 minutes and denaturation at 95°C for 10 minutes, then 40 cycles of the amplification step (denaturation at 95°C for 15 seconds and annealing/extension at 60°C for 1 minute). The ΔCT method, by means of the SDS software (Applied Biosystems), was used to calculate the mRNA levels. Oligonucleotide primers and probes for NIS and PAX8 were purchased from Applied Biosystems as Assays-on-Demand Gene Expression Products. Oligonucleotide primers and probes for CREM (Note: gene names are in italics) and the endogenous control β-actin are described in Table 2. Oligonucleotides for CREM detection are located on exons 4 and 5 of CREM (No. ENST00000374722 Ensembl), at the level of the encoding sequence of the Q2 domain, the glutamine-rich region present in the CREMt2α (see below).
CREM, cyclic AMP–response element modulator.
Cell lines and transient transfections
TPC-1 and BCPAP cell lines, derived from PTC, WRO derived from FTC and FRO cells, derived from human undifferentiated thyroid carcinoma, all authenticated for their thyroid origin (19), were grown in Dulbecco's modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (Invitrogen Corp.). Suberoylanilide hydroxamic acid (SAHA) was purchased from Cayman Chemical (Ann Arbor MI); valproic acid (VPA) was purchased from USP (Rockville, MD). SAHA and VPA treatment was performed for 48 hours at the dose of 4 μM and 6 mM, respectively. Transfections were performed by Lipofectamine 2000 reagent (Invitrogen Corp.) according to the manufacturer's instructions. The pGL3 promoter vector with NUE region (−9847 to −8968, NIS-NUE) is described elsewhere (11).
In the NIS-NUE construct, firefly luciferase is the reporter gene. Cells were grown in six-well plates to about 90% confluence. Unless otherwise noted, 1.3 μg firefly luciferase reporter construct was transiently cotransfected with 350 ng CREMα or CREMt2α expression vectors and 26 ng pRL-CMV (Renilla luciferase expression plasmid; Promega, Fitchburg, WI) to normalize the transfection efficiency among replicates. After incubation for 24 hours, the medium was replaced with a fresh medium. Treatment of cells with forskolin (10 μM) was performed for 24 hours. Firefly and renilla luciferase activities were determined 48 hours after the beginning of transfection by using the Dual-Luciferase Reporter Assay System (Promega).
Results
CREM, NIS and PAX8 gene expression in thyroid cancer tissues
We first evaluated by quantitative reverse transcriptase–PCR (RT-PCR) the expression of CREM, NIS, and PAX8 genes in a series of 38 thyroid cancer tissues, including 22 PTCs, 12 FTCs, and 4 ATCs, and, as control, 6 normal thyroid tissues (Fig. 1). As expected, compared to normal tissues, in nearly all tumor samples, we found a great reduction of NIS transcript. In FTCs and ATCs, such a reduction was extremely relevant with an 800- and 3000-fold decrease, respectively (Fig. 1A). Also in the case of PAX8, tumors displayed reduced gene expression. However, in PTC and FTC mean PAX8 mRNA levels were reduced only about twofold, and a significant overlap was present when the gene expression was analyzed in single cases. In ATC PAX8 expression was very much reduced (Fig. 1B). A surprising result was observed for CREM expression. In fact, mean levels of CREM mRNA were found to be increased in comparison with normal tissues about twofold in PTC and seven- to eightfold in FTC and ATC (Fig. 1C). Analysis of the data of PTCs, divided in subgroups according to the presence of BRAF V600E mutation, did not reveal any significant difference in CREM expression levels (data not shown). Similarly, subgroups of all differentiated thyroid cancers (PTCs and FTCs), according to the risk category (low or intermediate based on the American Thyroid Association risk stratification system) (20), showed significantly lower levels of mRNA expression only for NIS and not for PAX8 and CREM in the intermediate risk category (Fig. 2).
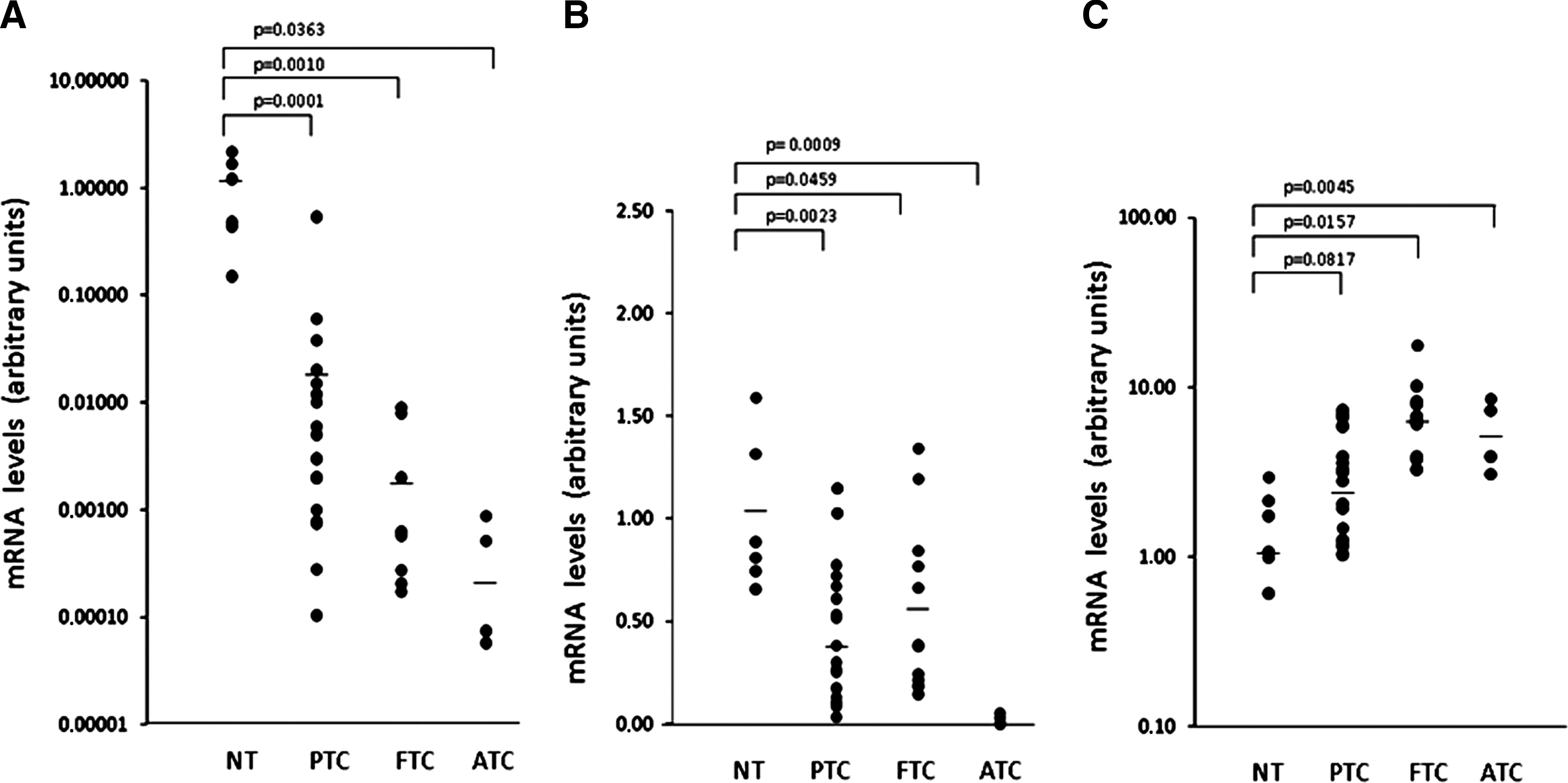
NIS, PAX8, and CREM mRNA levels in thyroid cancer. Expression of
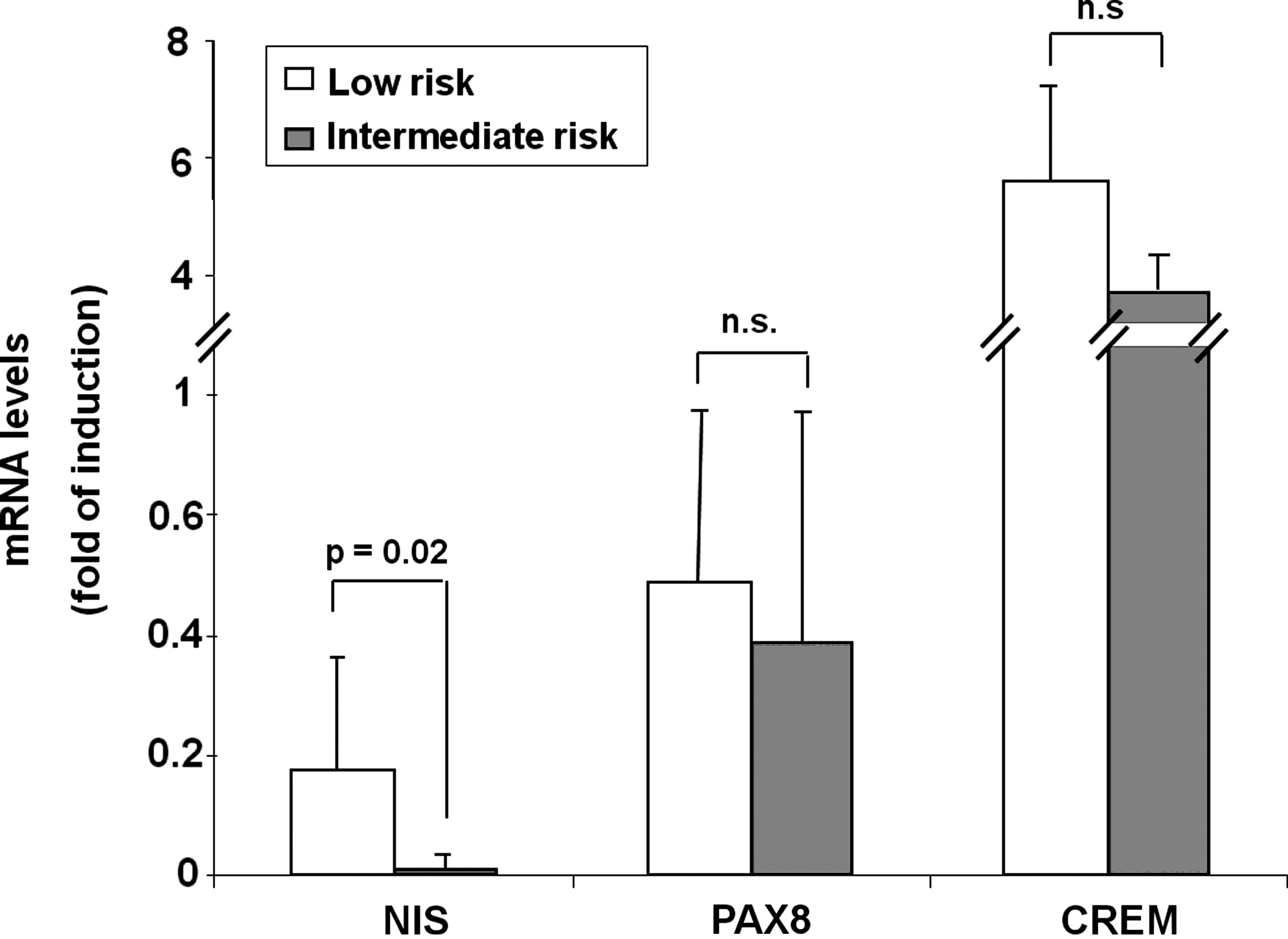
CREM mRNA levels according to risk category. All differentiated thyroid cancers were divided in subgroups according to risk category: low (n=15, white bars) or intermediate (n=15, gray bars); gene expression was evaluated by quantitative RT-PCR, as reported in Materials and Methods section. The p-values, obtained by Student's t-test, are indicated. n.s., not significant (p>0.05).
CREM action on NIS promoter
Findings shown in Figure 1 suggest that CREM may act as a repressor of NIS expression in thyroid tumor cells. To investigate the role of CREM in the control of NIS expression in thyroid cancer cells, we next performed cotransfection experiments, in which an expression vector for CREMt2α was cotransfected with a construct in which the transcription of the reporter LUC gene is controlled by the basal NIS promoter plus the NUE (NIS-NUE). Four different thyroid cancer cell lines were evaluated: BCPAP, TPC-1, WRO, and FRO. As shown in Figure 3, expression of CREMt2α induced an inhibitory effect in all the thyroid cancer cell lines examined, independently from the different basal levels. Thus, our data support the notion that in human cancer thyroid cell lines, CREM acts as a repressor of NIS expression.
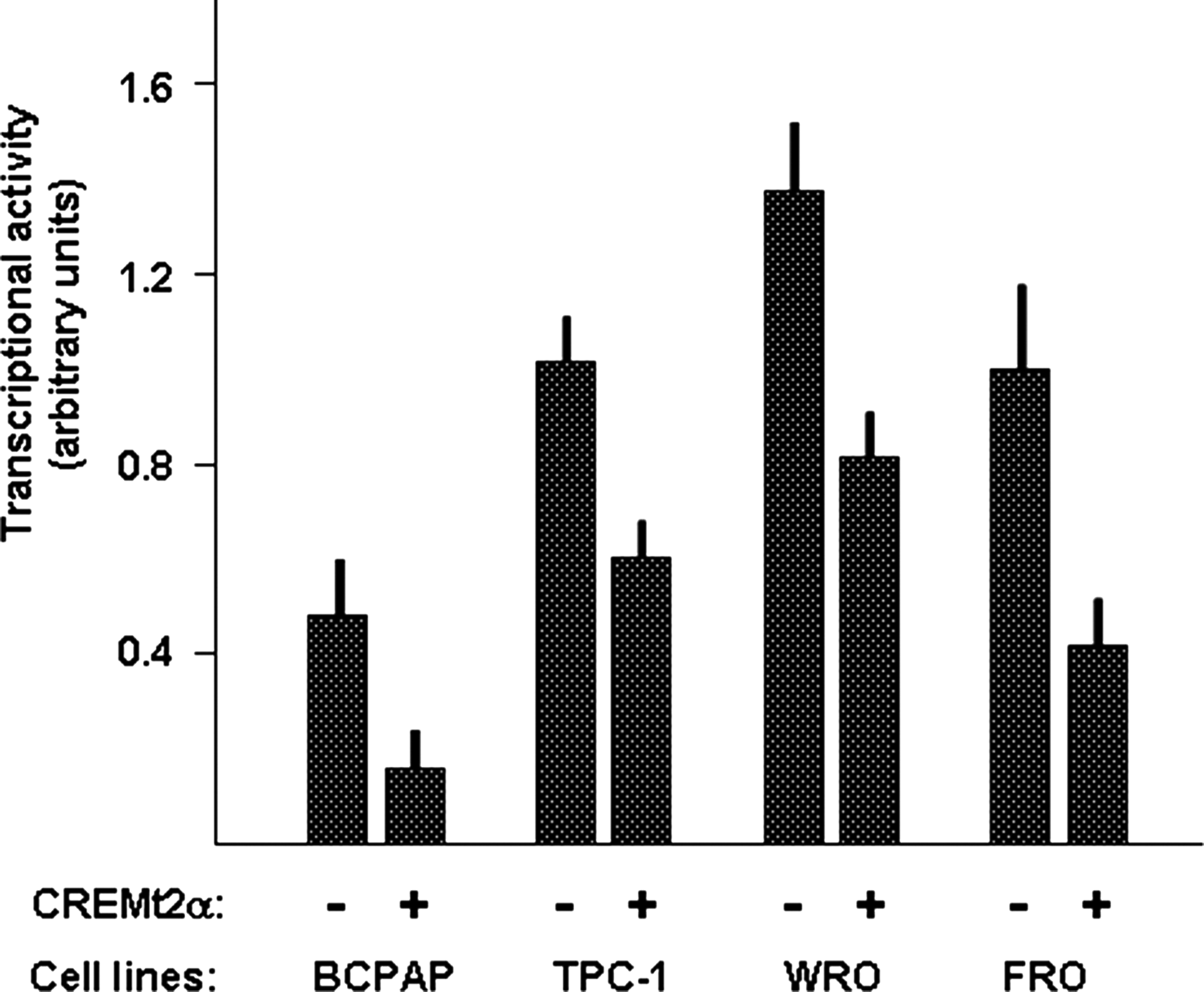
Effect of CREM on NIS promoter activity in human thyroid cancer cell lines. The indicated cell lines were cotransfected with the NIS-NUE reporter construct and with/without the expression vector for CREM (CREMt2α). Expression of Renilla luciferase was utilized for normalization of transfection efficiency among replicates. Each bar represents the mean value±SD of three independent transfections. NUE, NIS upstream enhancer.
It is known that increase of cyclic AMP levels in nontumorigenic rat thyroid cell lines by forskolin treatment greatly induces the NIS-NUE transcriptional activity (11). Thus, we tested whether forskolin treatment was able to modify the inhibitory effect of CREM on NIS-NUE in the context of tumor cell lines. As shown in Figure 4, in both TPC-1 and FRO cell lines, forskolin treatment increases the transcriptional activity of NIS-NUE but does not modify the inhibitory effect exerted by expression of CREMt2α; in both cell lines, CREMt2α exerts inhibition either in the absence or in the presence of forskolin.
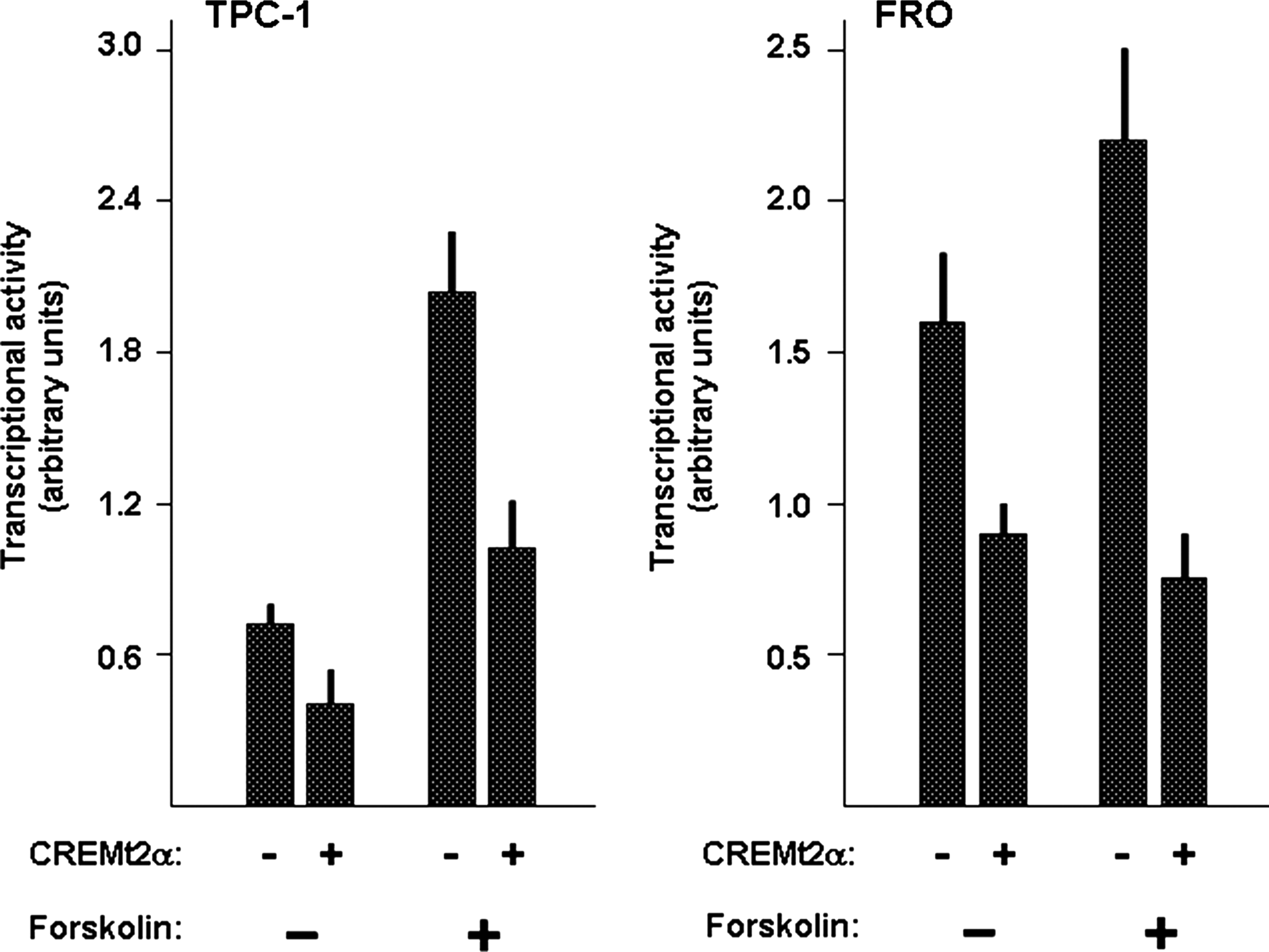
Effect of CREM on NIS promoter activity in the presence of forskolin. TPC1 and FRO cell lines were cotransfected with the NIS-NUE reporter construct and with/without the expression vector for CREM (CREMt2α) in the presence or absence of forskolin (10 μM) for 24 hours as described in the Materials and Methods section. Normalization for efficiency of transfection was performed as described in legend of figure 2. Each bar represents the mean value±SD of three independent transfections.
In order to gain an insight on the CREM effects on NIS-NUE construct, we used a vector able to express the CREB DNA-binding domain without the Q2 transcriptional activation domain (CREMα) that is present in the CREMt2α. Results of transfection experiments in TPC-1 and FRO cell lines are shown in Figure 5. In both cell lines, CREMα is not able to inhibit the NIS-NUE transcriptional activity. These data suggest that the presence of the Q2 domain is necessary for the CREM inhibitory effect.
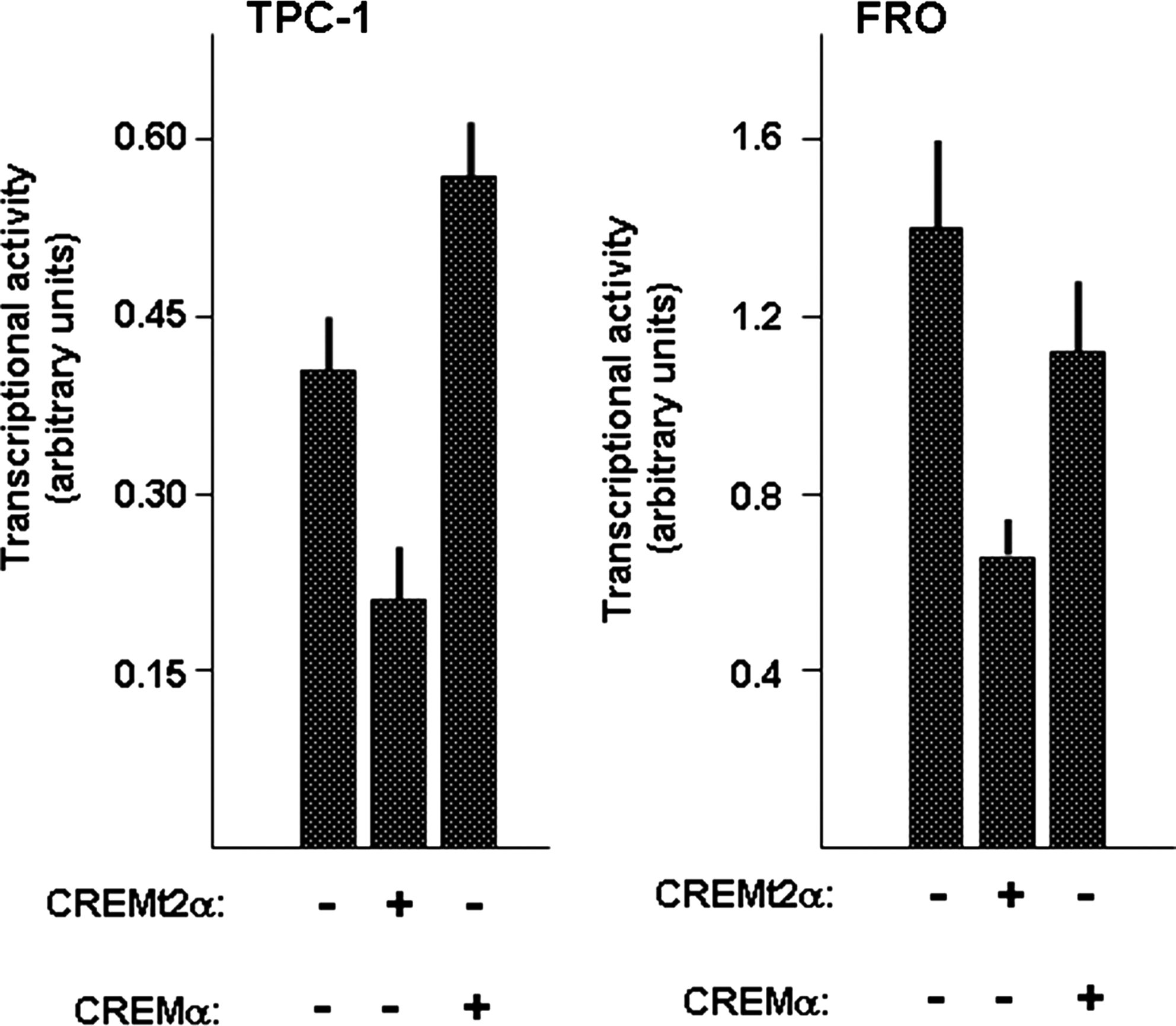
Effect of CREMt2α and CREMα on transcriptional activity of the NIS-NUE construct. TPC1 and FRO cells were cotransfected with the indicated constructs and the promoter activity was measured as described in the Materials and Methods section. Each bar represents the mean value±SD of three independent transfections.
We and others have reported that treatment of thyroid tumor cells with histone deacetylase (HDAC) inhibitors greatly increases NIS mRNA levels (21). Thus, if CREM inhibits NIS gene transcription, a decreasing effect of HDAC inhibitors on CREM expression could be expected. This hypothesis was tested by treating FRO cells with two different HDAC inhibitors, SAHA and VPA. Indeed both HDAC inhibitors increased NIS mRNA levels while decreasing CREM mRNA levels (Fig. 6).
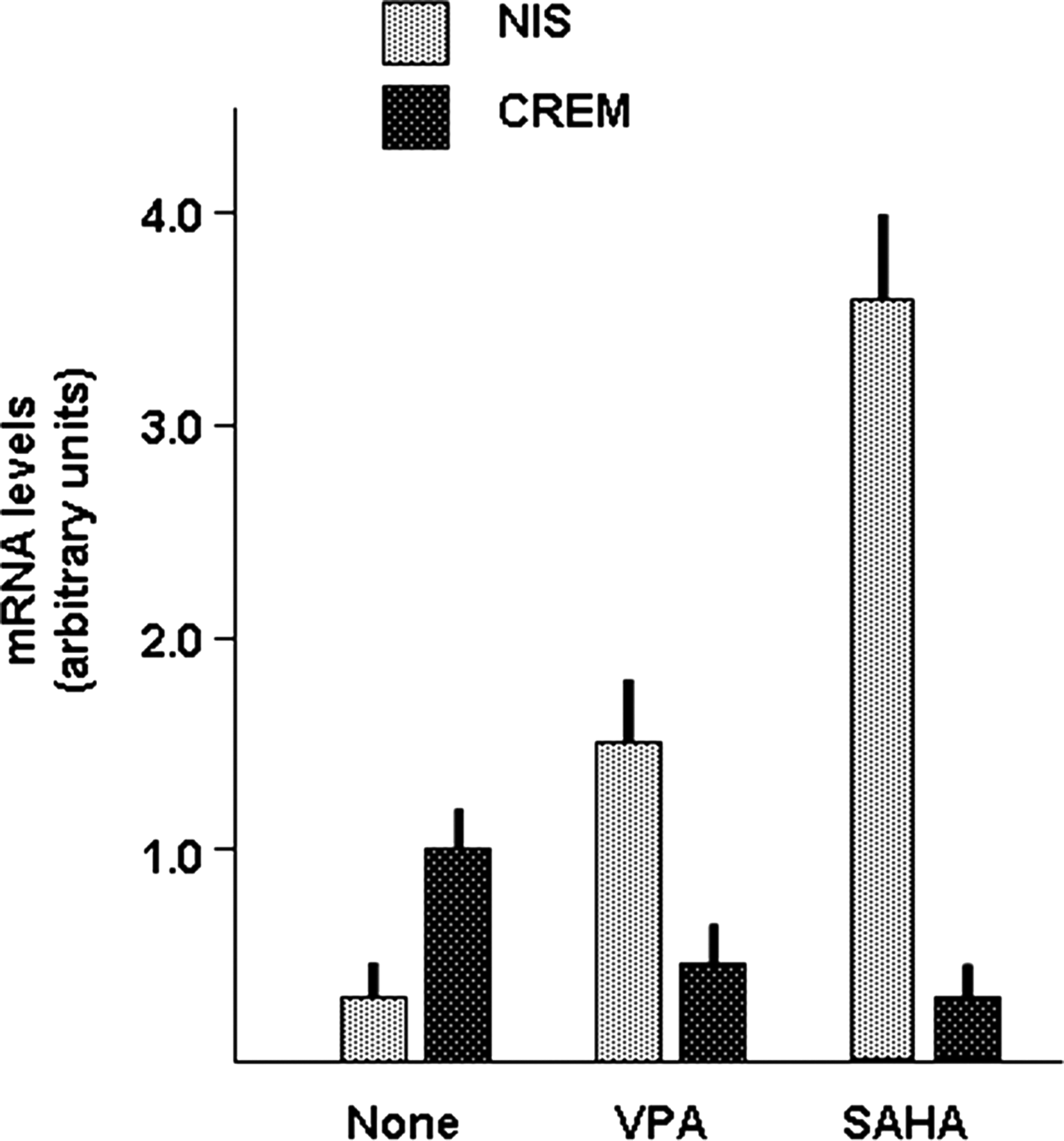
Effect of histone deacetylase inhibitors on NIS and CREM expression in FRO cells. The FRO cell lines was treated for 48 hours with valproic acid (VPA) (6 mM) or suberoylanilide hydroxamic acid (SAHA) (4 μM) and then NIS mRNA levels were measured as described in the Materials and Methods section. Each bar represents the mean value±SD of three independent measures.
Discussion
Investigation of the mechanism of control of NIS transcription has revealed the existence, in both rat and human NIS promoter, of a NUE region, target of a complex of regulatory elements both cyclic AMP–dependent and independent (11 –15). Among them, a role of the CREB-family component CREM has been documented in our previous in vitro studies (11 –16). In normal thyroid FRTL-5 cells, CREM overexpression stimulates the transcriptional activity of the NUE only after forskolin treatment (16). Such a finding prompted us to hypothesize that an altered expression of CREM may be involved in silencing of NIS expression occurring in most thyroid cancer tissues, as already documented for the thyroid specific transcription factor, PAX8 (14). Surprisingly, analysis of mRNA levels in a series of thyroid cancer tissues revealed that, while in NIS-defective tumors, PAX8 mRNA levels were reduced in a parallel way, CREM mRNA levels were increased in all thyroid cancers examined, independent of histotype or, for the PTCs risk category. In addition, when transfected into thyroid cancer cells, CREM expression had an inhibitory action on the NIS promoter. Similar data were observed in various thyroid cancer cell lines presenting different activated oncogenic pathways. The effect of CREM expression was not observed when the CREMα construct is used, indicating that the presence of the Q2 domain is essential for the inhibitory action. The Q2 domain is a glutamine-rich region and is considered a transcriptional activator (22). According to this notion, an inhibiting action of CREM has been reported in the past only for CREM lacking the Q1 and Q2 regions (23). Our data suggest instead, that in particular cellular contexts, the Q2 domain may act as a transcriptional repressor. In agreement with our data, in several conditions it has been demonstrated that glutamine-rich domains may act as transcriptional repressors (24 –27). Indeed, by performing X-ray crystallography on the glutamine-rich domain of HDAC4, it has been proposed that a general role of glutamine-rich motifs is to establish protein–protein interactions among the components of polar interaction networks (28). Thus, we propose that while in the context of nontumorigenic cells CREM acts as a transcriptional activator of NIS, in several tumor cells CREM acts as a repressor of NIS transcription. This dual role has been described for several other transcriptional activators including SP1, c-FOS, YY1, and MYC. For these factors, in fact, it has been shown that they can induce transcriptional activation or inhibition, depending on the context (29 –32). Such a dual function has been shown to contribute to the difference existing between normal and neoplastic cells. The transcription factor NKX2.2 contains both transcriptional activation and transcriptional repression domains; by investigating Ewing's sarcoma cells, however, it has been shown that only the transcriptional repressive function is necessary and sufficient for the oncogenic phenotype (33). In our hypothesis, CREM switches from an activator to a repressor in the context of the same promoter. This behavior has been demonstrated for other transcriptional regulators. It has been reported, in fact, that the transcriptional cofactors Brg-1 and BRM can switch their mode of function at the same promoter between activation and repression (34). Moreover, by using Drosophila embryos, Chen et al. have shown that the transcription factor Hsf is converted from a repressor to an activator of the tailles gene transcription by a MAPK pathway-induced phosphorylation (35). The question is, “How is CREM converted from activator to repressor during thyroid cell transformation?” Probably, this change is due to modification of the network of transcriptional regulators bound to the proximal promoter of NIS. Accordingly, our unpublished data indicate that forskolin stimulation reduces HDAC1 bound to the proximal promoter of NIS in normal thyroid cells; in contrast, it induces the reverse effect in a thyroid cancer cell line containing RET/PTC.
The CREM-mediated inhibition of the promoter of NIS seems to better reflect what occurs in human thyroid tumors in vivo, in which CREM mRNA levels are maintained or increased, as a result of genetic (oncogene-driven) or epigenetic changes occurring after neoplastic transformation. Indeed, epigenetic alterations have been extensively reported in thyroid cancer cells, influencing NIS, as well as other differentiating thyroid-specific gene expression (36). In this regard it is noteworthy that, in FRO cells, inhibitors of HDAC that were able to restore NIS expression induce downregulation of CREM mRNA levels. The complexity of regulation of NIS gene transcription by the CRE-binding family members is also demonstrated by the finding that CREB was found functionally downregulated in hyperfunctioning thyroid adenomas (37); these are know to overexpress NIS.
Several agents have been and are currently being tested in preclinical models of thyroid cancer to determine if they restore the radioiodine concentration capacity, based on the re-expression of the NIS in thyroid tumor cells. The characterization of the complex regulatory mechanisms involved in the NIS transcription and parallel studies of their alteration in thyroid tumor tissues will help to better define the targets of novel redifferentiating therapeutic strategies for those tumors refractory to current treatment.
Footnotes
Acknowledgments
This work was supported by grants to G.D. from Associazione Italiana per la Ricerca sul Cancro (AIRC) (No. IG 10296) and MIUR (PRIN No. 20093WAPIK_003), to M.F. from the American Thyroid Association, and to S.F. from The Umberto Di Mario Foundation and from the Banca d'Italia.
Disclosure Statement
The authors declare that no competing financial interests exist.