
Abstract
Background:
We have previously demonstrated that expression of COX-2 is upregulated by hepatocyte growth factor in thyroid papillary carcinoma (TPC) cells and is associated with increased invasiveness of tumor cells. COX-2 upregulation was associated with downregulation of KAI-1/CD82, a metastasis suppressor molecule that has been associated with the metastatic potential of several solid tumors. In the present study, we have investigated the possibility that downregulation of KAI-1/CD82 may contribute to the invasiveness of papillary carcinoma cells.
Methods:
Expression of KAI-1/CD82 and its relation to COX-2 levels were investigated in 6 primary cultures of TPC, in 2 tumor cell lines (TPC-1 and K1), and in 55 tumor samples of TPC. The biological role of KAI-1/CD82 in regulating tumor invasiveness was investigated in TPC cell lines and primary cultures transfected with a pCDNA3.1/Hygro.KAI-1; transfected cells were tested in functional studies of cell migration and invasiveness. Finally, the role of KAI-1/CD82 in influencing TPC metastasis was investigated in vivo using nu/nu mice injected with K1-transfected cells.
Results:
We provide evidence that COX-2 and KAI-1/CD82 are inversely regulated in TPC primary cultures and in TPC-1 tumor cells. In fact, inhibition of COX-2 with NS398 is associated with a 2–9-fold upregulation of KAI-1/CD82 RNA. Moreover, a possible relation between COX-2 and KAI-1/CD82 was confirmed by the presence of a statistically significant inverse correlation in the expression of the two genes in 55 tumor samples of TPC (r=−0.513; p=0.001). In 36 of 55 cases, tumor areas contained lower levels of KAI-1/CD82 RNA as compared with the corresponding normal tissue. Low expression of KAI-1/CD82 RNA in the tumor area was associated with extrathyroid extension of the disease in 16 of 19 cases (p<0.04) and with lymph node metastasis in 11 of 14 cases (not significant). KAI-1/CD82 re-expression in tumor cells was associated with a significant decrease in their migratory (50–76% reduction) and invasive (46–65% reduction) capacity, even after hepatocyte growth factor stimulation. Finally, nu/nu mice injected with KAI-1/CD82–transfected K1 TPC cells developed fewer and smaller metastasis as compared with mice injected with vector-transfected K1 cells (p=0.016).
Conclusion:
Our findings raise the possibility that downregulation of KAI-1/CD82 in TPC cells is one of the molecular mechanisms regulating their invasive and metastatic potential.
Introduction
The mechanism of action of KAI-1/CD82 as a metastasis suppressor is under investigation. Tetraspanins contain no intrinsic catalytic activity, and it is currently believed that they serve as master regulators of membrane organization. In fact, they regulate a variety of cellular events, including signaling, cell adhesion, migration, survival, and endo- and exocytosis through interactions with other surface molecules (2,3). It has been shown that KAI-1/CD82 modulates the activity of beta-1 integrins (13 –15), alpha-v/beta-3 (16), and of the growth factor receptors EGFR (17) and c-Met (15,18 –20). Through these interactions, KAI-1/CD82 profoundly reduces the migratory capacity of Met-positive tumor cells derived from lung, prostate, and pancreatic carcinomas, even after hepatocyte growth factor (HGF) stimulation (15,18 –20).
Thyroid papillary carcinoma (TPC) is characterized by a high expression of Met protein, the high affinity HGF receptor in >95% of cases (21). HGF stimulation of TPC cells increases their invasiveness and migratory activity (22), and causes an upregulation of COX-2 (23). In the present study we provide evidence that inhibition of COX-2 is associated with a significant upregulation of KAI-1/CD82 and with increased invasiveness of TPC cells.
Materials and Methods
Patients and tissue samples
This study was performed according to the informed consensus law of Italy. Fifty-eight thyroid specimens involved by TPC were collected between June 2004 and July 2009, and were stored in the tumor tissue bank of the Pathology Unit, Ospedale Sant'Andrea, Rome, Italy. The patients were 15 men and 43 women with a mean age of 47 years (range 20–77 years). Fragments of tumor tissue and of normal thyroid were snap-frozen in liquid nitrogen, and stored at −80°C until sectioning; the remaining thyroid tissue was formalin-fixed and paraffin-embedded for conventional histology.
Expression levels of KAI-1/CD82 and GAL3 proteins were investigated in paraffin sections using immunohistochemistry and a monoclonal antibody anti-KAI-1 (Clone G2; 1:50 dilution; Santa Cruz Biotechnology) and a monoclonal antibody anti-GAL3 (1:200 dilution; kindly provided by Dr. Armando Bartolazzi [Department of Clinical and Molecular Medicine “Sapienza” University, Sant'Andrea Hospital, Rome, Italy]); paraffin sections of a breast carcinoma were used as a positive control. Immunohistochemistry was performed with the automatic immunostainer Link 48 (Dako), using the EnvisionTM Flex+, mouse, High pH (LinK) kit (Dako).
Primary cultures and papillary carcinoma cell lines
Primary cultures of neoplastic thyroid cells were established as previously described (22). In brief, fragments of 6 papillary carcinomas were digested with a collagenase–hyaluronidase mixture (Sigma) for 2 hours at 37°C. Cells were washed three times with phosphate buffered saline (PBS) and were plated on Primaria plates (Falcon) at a density of 1×106 cells/75 mL. Primary cultures were maintained in Dulbecco's modified Eagle's medium (DMEM), supplemented with 10% fetal bovine serum (FBS; Life Technologies), and were split every 15 days for a period of 90 days. Two human papillary thyroid carcinoma cell lines, TPC-1 and K1 (kindly provided by Dr. Salvatore Sciacchitano), were cultured in DMEM and DMEM:Hamm's F12:MCDB 105 medium, respectively (Gibco) containing 10% FBS (Life Technologies). K1 and TPC-1 are authentic thyroid cancer cell lines as demonstrated by Schweppe et al. (24).
Primary cultures and tumor cell lines were stimulated with the COX-2 inhibitor NS-398 at different concentrations (0–50 μM; Sigma) or with a dimethylsulfoxide-containing control medium for 20 hours. NS-398 was dissolved in dimethylsulfoxide (Sigma) and then diluted in the growth medium to the working concentration. In some experiments, TPC-1 and K1 cells were starved in DMEM containing 0.5% FBS for 18 hours, and then stimulated with HGF (100 ng/mL; R&D Systems).
Real-time polymerase chain reaction array analysis
Quantitative real-time polymerase chain reaction (RT-PCR) array analysis was carried out using an RT2Profiler PCR array purchased from SuperArray Bioscience Corporation. Briefly, pooled total RNA (3 μg) obtained from primary cultures and tumor cell lines stimulated or not with the COX-2 inhibitor NS-398 was converted into first-strand cDNA using RT2 First Strand Kit (SuperArray). The cDNA template was mixed with the ready-to-use RT2 Real Time SYBR Green PCR Master Mix (Superarray) containing a panel of 84 primer sets for pathway-focused genes (Human Invasiveness and Metastasis; Superarray APH5028) plus five housekeeping genes and three RNA and PCR quality controls. The mixture was aliquoted into each well of the same plate containing predispensed gene-specific primer sets, and PCR was performed on iCycler Biorad. Data were analyzed using the PCR Array Data Analysis Web Portal. Results were expressed as relative levels of each gene mRNA under the effect of NS398 referred to the expression of this gene in unstimulated cells (that were chosen to represent 1× expression of each gene). Results were considered significant when mRNA expression was threefold higher or lower than that of unstimulated cells. Experiments were performed in triplicate.
Transfections
TPC-1 and K1 cell lines and two primary cultures were transfected with the plasmid construct pCDNA3.1/Hygro.CD82 (Invitrogen) using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. TPC-1, K1, and primary cultures at 5×104 cells for a 35-mm-diameter plate were transfected with 2 μg of vector+CD82 and 0.2 μg of a fluorescent reporter green fluorescent protein. About 4 hours after transfection, the culture medium was changed to a regular medium with supplements. About 24 hours later, the cells were tested for transfection efficiency at RNA level by real-time PCR and at protein level with immunofluorescence. For stable transfection, the culture medium containing Hygromycin-resistant cells was maintained 10 days later and screened for CD82 expression. Cells created by transfection in a similar fashion with the empty vector were used as controls.
Immunofluorescence
Cells before and after transfection were allowed to attach overnight to chambered glass slides. The cells were then rinsed in PBS; fixed in ice-cold methanol; washed; blocked in a PBS-containing globulin-free bovine serum albumin (BSA; 1%), Triton X-100 (0.3%), and goat serum (3%); incubated with primary antibodies (monoclonal anti-CD82, clone 5B5 1:50 dilution; Thermo Scientific) in a humidified chamber for 1 hour at room temperature; washed in PBS; and incubated again for 1 hour in the dark with an fluorescein isothiocyanate–coupled secondary antibody (1:50 dilution). After washing, the cell lines were covered with DAPI solution (Roche). Slides were coverslipped and examined by fluorescence microscopy.
Real-time PCR analysis
Thyroid tissue expression of KAI-1/CD82 was investigated at RNA level. Frozen sections of cryopreserved tissues were used for RNA extraction. Tumor tissue and normal thyroid tissue were isolated using the microdissection laser system SL CUT (Nikon) and total RNA was obtained using the Pico Pure Isolation Kit (Arcturus). The integrity of the RNA was assessed by denaturing agarose gel electrophoresis and spectrophotometry. KAI-1/CD82 expression was also evaluated in RNA extracts obtained from six cases of primary cultures of papillary thyroid carcinoma (PTC), and in two cell lines (TPC-1 and K1) of papillary carcinoma of the thyroid prior or after incubation with the COX-2 inhibitor NS-398 at different concentrations (0–50 μM) and in RNA extracts obtained from KAI-1/CD82–transfected TPC-1 and K1 cells. RNA transcripts for KAI-1/CD82 were measured by real-time absolute quantitative RT-PCR, based on TaqMan methodology, using the ICycler System (Biorad). Gene-specific primers and probes were as follows: KAI-1/CD82, forward 5-CCG ACA AGA GCA GTT TCA TCT CT-3, reverse 5-AAG ACA TAG GCC CCC ATC CT-3, and probe 5-CCT GCA AAC CTC CTC CAG CTC GCT-3. To normalize the amount of total RNA present in each reaction, we amplified the housekeeping gene β-actin. Measurements were performed in triplicate.
Western blot and immunoprecipitation
Expression of KAI-1/CD82 protein was examined using Western blot analysis. Briefly, lysates from TPC1 cells were obtained using a lysis buffer containing 1 mM ortho-vanadate and a cocktail of protease inhibitors (Sigma), and maintained in lysis buffer at 4°C for 15 minutes. Lysates were washed with an ice-cold lysis buffer, eluted, and denatured by heating for 5 minutes at 95°C in reducing Laemmli buffer. Proteins were resolved on sodium dodecyl sulfate polyacrylamide gel electrophoresis (8% SDS-PAGE) and transferred onto nitrocellulose filters. Filters were blocked with 5% BSA for 1 hour and probed with an anti- KAI-1/CD82 monoclonal antibody (G-2 clone 1:250; Santa Cruz Biotechnology). The reactions were revealed by an ECL Western blot detection system (Amersham Pharmacia Biotech). To normalize the amount of total protein present in each sample, filters were probed with an anti-Vinculin monoclonal antibody (1:1000; UCS Diagnostics).
To evaluate the effect of KAI-1/CD82 re-expression on Met tyrosine-phosphorylation confluent, K1 and K1/KAI-1/CD82–transfected cells cultured on 10 cm dishes were starved of serum for 24 hours and treated with different concentrations of HGF (0, 25, and 100 ng/mL; RD Systems) for 10 minutes. For immunoblots, 1 mg of cell lysate was immunoprecipitated with DQ13 anti-Met antibody, resolved by SDS-PAGE, transferred to a nitrocellulose membrane (Bio-Rad), and incubated with PY20 anti-phosphotyrosine antibody and re-blotted with DL21 anti-Met antibody. Protein bands were visualized using horseradish peroxidase–conjugated secondary antibodies and an enhanced chemiluminescence reagent (Amersham Pharmacia Biotech).
In vitro migration and invasiveness
The effects of KAI-1/CD82 transfection on the migratory and invasive capacities of TPC cells were evaluated using the BioCoat Invasion Chamber system (BD Biosciences). The Matrigel invasion chambers, containing an 8-μm-pore-size PET membrane, were treated with Matrigel Basement Membrane Matrix (for invasion test) or with BSA (for migration test). About 1.5×105 KAI-1/CD82–transfected or vector-transfected cells diluted in 0.2% FBS-DMEM were added to the upper compartment; 2.5 mL of 10% FBS-DMEM or 100 ng/mL HGF (RD Systems) was added to the lower compartment. The migration assay was performed for 24 hours in a humidified tissue culture incubator at 37°C and in a 5% CO2 atmosphere. After incubation, nonmigrating cells were removed by scrubbing, and migrating cells present on the lower surface of the membrane were stained with Diff-Quick; cells present in 10 high powder field were counted in each filter at 400×. The data are given as mean±SD of triplicates filters.
Rac1 mRNA, protein expression, and activation assay
Equal numbers of K1 CD82/KAI-1–transfected and vector-transfected cells (3×105 cells per 60 mm culture dish; 60% confluency) were left untreated in the medium at 37°C in 5% CO2. After 1 day, total RNA was isolated using Trizol reagent (Amersham Pharmacia Biotech). Two micrograms of total RNA was reverse transcribed to synthesize cDNA using an AccuPower RT-PCR Mix kit (Bioneer). The PCR amplification was performed with 10 μL of RT reaction mixture using the following thermocycling profile: 1 cycle at 94°C for 5 minutes; 30 cycles at 94°C for 1 minute, 55°C for 1 minute, and 72°C for 2 minutes; and then 1 final cycle at 72°C for 10 minutes. β-Actin was used as an internal control. The primers used were Rac1 forward 5′-ATC TCG AGA TGC AGG CCA TCA AGT GTG TG-3′ and Rac1 reverse 5′-CCG AAT TCT TAC AAC AGC AGG CAT TTT CT-3′. The PCR products were electrophoretically resolved on a 1% agarose gel and then analyzed under UV light after staining with ethidium bromide.
Levels of RAC1 protein were evaluated in vector- and KAI-1/CD82-transfecetd cells by Western blot analysis (see Western Blot and immunoprecipitation section) using a rabbit monoclonal antibody anti-RAC-1 (1:500; Novus Biologicals).
Intracellular Rac1 activity was measured using a StressGen's StressXpress1 Rac1 Activation Kit (StressGen) according to the manufacturer's instructions.
In vivo lung metastasis assay
All procedures involving animals and their care are in compliance with national and international laws. Female CD-1 nude (nu/nu) mice (6–8 weeks old and 22–24 g in body weight) were purchased from Charles River Laboratories. Each experimental group included four mice. Experimental lung metastases were obtained by injecting mice with 6.5×106 viable cells (KAI-1/CD82 transfected or vector transfected) into the tail vein (intravenous) and killing mice 60 days after tumor implant. Lungs were excised and divided into two portions: one half was fixed in Bouin's solution and the presence of metastatic nodules was evaluated using a dissecting microscope. All the lung lesions evidenced by microscopy after staining of lungs with Bouin's, regardless of their size, were considered positive. For histopathological determination of the number and size of lung metastases, the other half of the lung from mice were formalin-fixed, paraffin-embedded, and stained with hematoxylin and eosin and with a specific human thyroid marker (GAL3).
To assess the differences in the lung metastatic ability between different mice groups, the Student's t-test for unpaired data (two-sided) was used. A p-value <0.05 was considered statistically significant.
Statistical analysis
Patient data were analyzed by Statistical Package for Social Sciences (SPSS for Windows, version 14.0). The association among all data was obtained using the Fisher's exact test or the Spearman's correlation test. A p-value <0.05 was considered statistically significant. Statistical analysis of the in vitro experiments was carried out using the Student's t-test or the Mann–Whitney U-test. A p-value <0.05 was considered statistically significant.
Results
Expression levels of COX-2 and KAI-1/CD82 are inversely correlated in TPC tumor cells
In a recent publication, we have demonstrated that expression of COX-2 is upregulated in TPC cells (23). In order to evaluate the downstream effects of COX-2 activation, we have blocked COX-2 with NS398, and we have tested the RNA in a microarray real-time assay measuring the expression of 84 genes involved in invasiveness and metastasis. It was found that COX-2 inhibition was associated with a dramatic upregulation of KAI-1/CD82 transcription (Supplementary Table S1; Supplementary Data are available online at
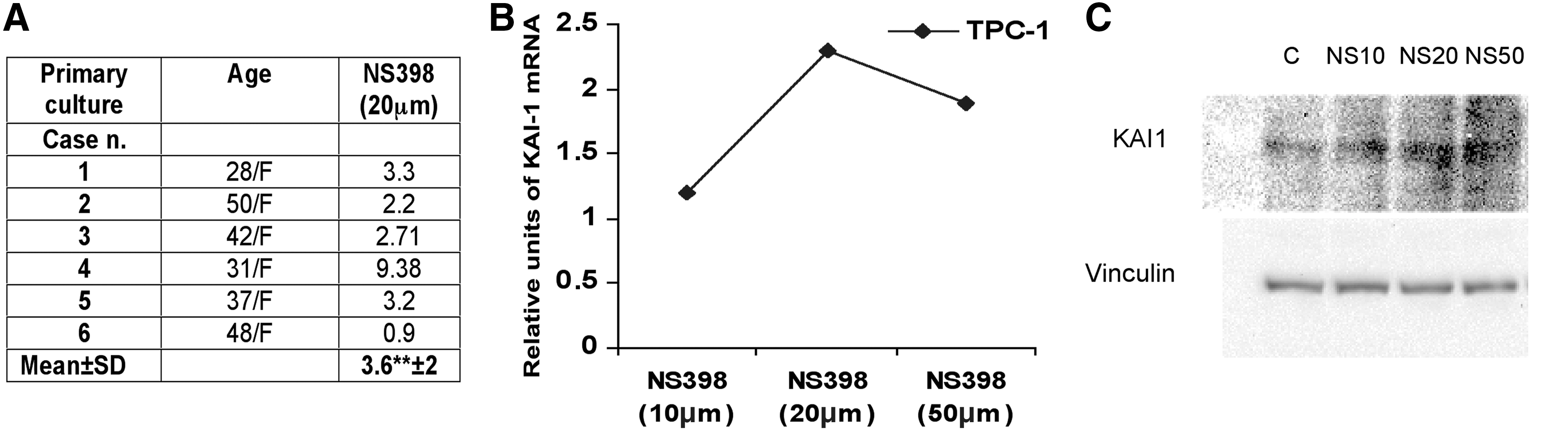
NS398 treatment causes an increase of KAI-1/CD82 mRNA expression in primary cultures of TPC and in the papillary carcinoma cell line TPC-1. Primary cultures of TPC cells
The levels of expression of KAI-1/CD82 RNA were investigated in frozen sections of 55 samples of tumor tissue in which total RNA was extracted from the tumor area and from the surrounding normal thyroid tissue. It was found that in 36 of 55 cases (65%) the tumor areas contained lower levels of KAI-1/CD82 RNA as compared with the corresponding normal thyroid tissue; these cases were classified as Low Tumor-to-Benign Ratio; the remaining 19 cases were classified as High Tumor-to-Benign Ratio. In Table 1, the clinicopathologic features of the two groups (Low Tumor-to-Benign Ratio vs. High Tumor-to-Benign Ratio) are compared; it was found that low expression of KAI-1/CD82 RNA was significantly associated (p<0.04) with extrathyroid extension on the disease, and was more often present in those cases with lymph node metastasis (30% vs. 15%, n.s.). Low expression of KAI-1/CD82 in tumor tissue was confirmed also by immunohistochemistry since PTC cells of most cases exhibited only weak and focal membrane immunostaining for KAI-1/CD82 (Fig. 2). Finally, we have previously reported that TPC cells in tumor sections express higher levels of COX-2 as compared with the corresponding normal tissue (23). The existence of a possible relation in the expression of COX-2 and KAI-1/CD82 in each single case was investigated at RNA level in 58 tumor samples (Fig. 3); it was found that there is a statistically significant inverse correlation (r=−0.513, p=0.001) in the expression of the two genes.
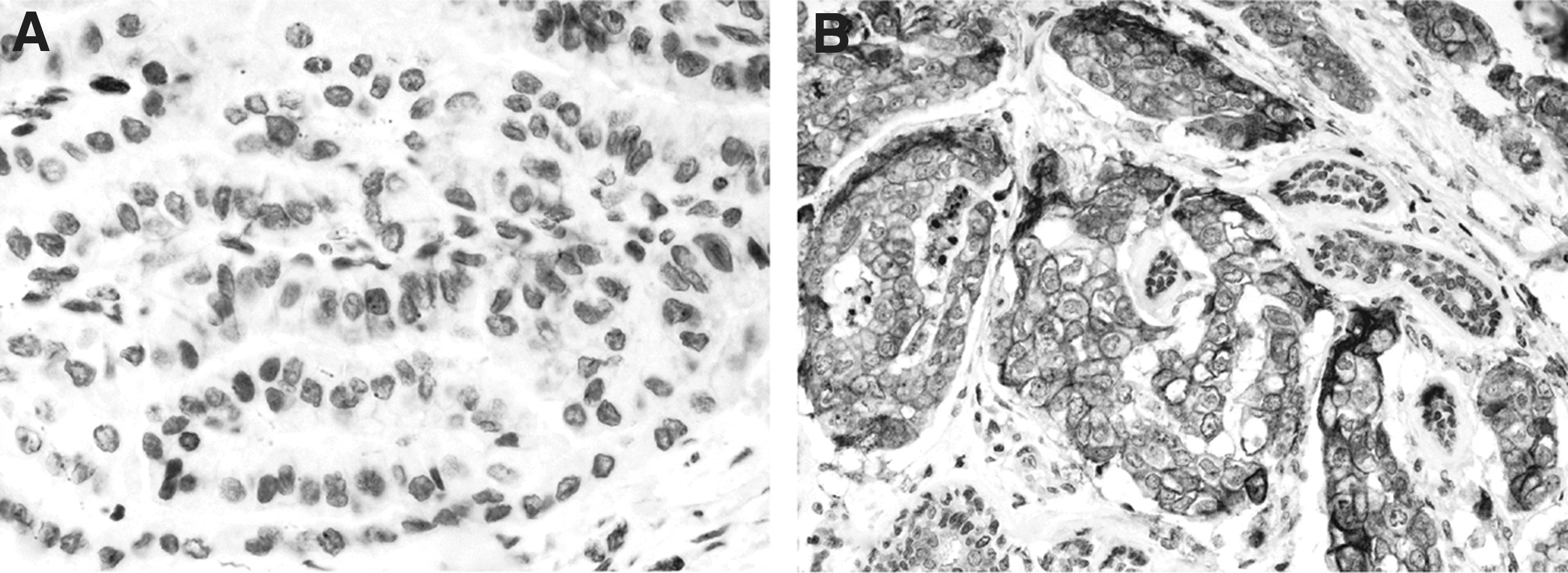
Expression of KAI-1/CD82 in tumor tissue.
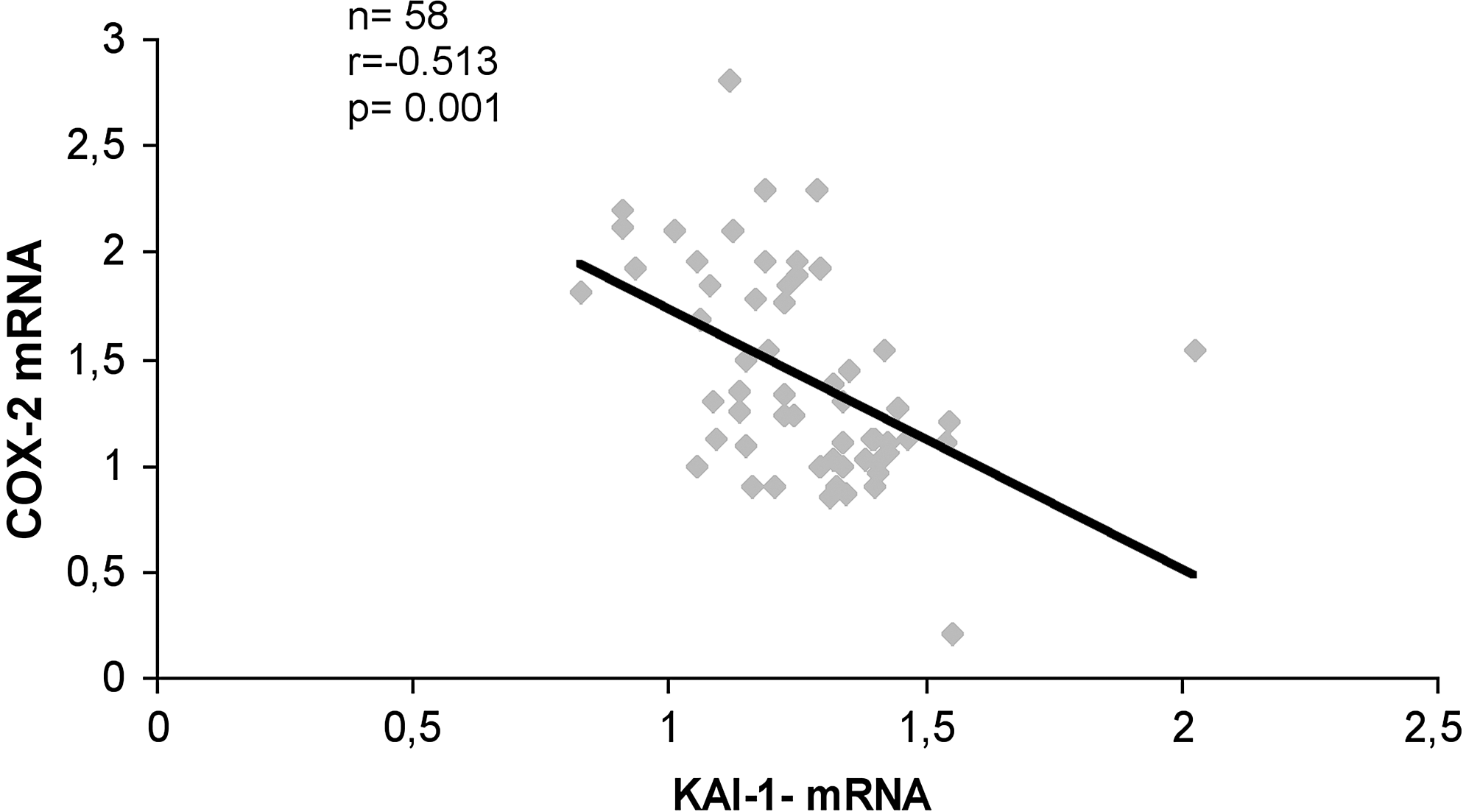
Expression levels of KAI-1/CD82 and COX2 mRNA are inversely correlated in tumor tissue. Total RNA was extracted from frozen sections of 58 tumor tissue samples of papillary thyroid carcinoma. The KAI-1/CD82 and COX-2 expression levels were determined by quantitative real-time polymerase chain reaction comparative Ct method. Spearman correlation coefficient r=−0.513, p=0.001.
Total RNA was extracted from tissue samples obtained from the tumor and from the corresponding normal thyroid of the same patients in 55 cases of papillary thyroid carcinoma. RNA transcripts for KAI-1/CD82 were measured by quantitative real-time polymerase chain reaction (see Materials and Methods).
Patients were grouped into two cohorts: High Tumor-to-Benign Ratio and Low Tumor-to-Benign Ratio. The tumor-to-benign ratio (TTBR) was calculated by comparing KAI-1 expression in papillary thyroid carcinoma vs. KAI-1expression in corresponding peritumoral normal thyroid tissue. TTBR>1 was considered as high, while TTBR<1 was considered as low.
Correlation analysis between KAI-1 expression levels and clinicopathological parameters was carried out using Fisher's exact test. p-Values <0.05 were considered statistically significant.
Thirty of 55 patients were not clinically suspected of having lymph node metastases, whereas in 25 patients enlarged cervical lymph nodes were dissected; of these, lymph node metastases were detected only in 14 cases.
The biological role of KAI-1/CD82 in TPC cells
To characterize the biological role of KAI-1/CD82 in papillary carcinoma of the thyroid, tumor cells of K1, TPC-1 cell lines, and of two primary cultures of TPC cells were stably or transiently transfected with a KAI-1/CD82 cDNA. Vector-transfected clones were generated as controls. Transfection efficiency was evaluated at RNA level and was confirmed with immunofluorescence. Stable KAI-1 transfection caused a 30–107-fold increase of KAI-1/CD82 mRNA levels, as compared with vector-transfected TPC-1 and K1 cells; this event was associated with a bright immunofluorescence staining of tumor cells at membrane level in both stably and transiently KAI-1/CD82–transfected cells (Fig. 4).

Immunofluorescent staining for CD82 in stably and transiently transfected TPC cells. The efficiency of transient
KAI-1/CD82–transfected and vector-transfected cells were tested for migratory and invasive capacity using the BioCoat Invasion Chamber system (Fig. 5). In repeated experiments, it was found that KAI-1/CD82 re-expression was consistently associated with a marked and significant reduction in the migratory (50–76%) (Fig. 5A) and in the invasive capacity (46–65%) (Fig. 5B) of TPC tumor cells. These findings were confirmed also using TPC cells obtained from two primary cultures.

Migratory and invasive capacity of KAI-1/CD82–transfected TPC cells. The effect of KAI-1/CD82 re-expression was tested using the TPC1 and K1 cell lines and TPC cells obtained from two primary cultures of papillary carcinoma of the thyroid. KAI-1/CD82–transfected or nontransfected TPC cells, diluted in 0.2% FBS-DMEM, were added to the upper compartment of the BioCoat Invasion Chamber; in the lower compartment, 2.5 mL of 10% FBS-DMEM was added as chemoattractant. The migration
It has been reported that KAI-1/CD82 re-expression reduces the HGF-induced migratory capacity of Met-positive tumor cells (15,18 –20). We have investigated this possibility in the experiments reported in Figure 6A, where it was found that K1 cells transfected with KAI-1/CD82 were no longer responsive to stimulation with 100 ng/mL HGF, whereas vector-transfected cells were still able to migrate and to invade Matrigel. Reduced motility and invasiveness of KAI-1/CD82–transfected cells were associated with low levels of c-Met tyrosine-phosphorylation HGF stimulation (Fig. 6B); this finding suggests that KAI-1/CD82 re-expression reduces or abolish HGF-induced Met activation.

KAI-1/CD82 re-expression reduces HGF-induced Met-activation.
Cell motility requires cytoskeleton reorganization. It has been shown that KAI-1/CD82 inhibits expression and activation of RAC1, a small GTP-binding protein involved in the regulation of various cellular functions, including cytoskeletal reorganization (18). In the experiments reported in Figure 7, the levels of expression and of activation of RAC1 were evaluated in vector-transfected or KAI-1/CD82–transfected K1 and TPC-1 cells. It was found that KAI-1/CD82–transfected cells and vector-transfected cells expressed comparable amounts of RAC-1 at protein level and at RNA level (Fig. 7A, B); however, the active form of RAC-1 was dramatically reduced in KAI-1/CD82–transfected cells (Fig. 7C). Our findings are consistent with the possibility that re-expression of KAI-1/CD82 profoundly interferes with HGF-induced GTP-mediated intracellular signaling and cytoskeleton organization.
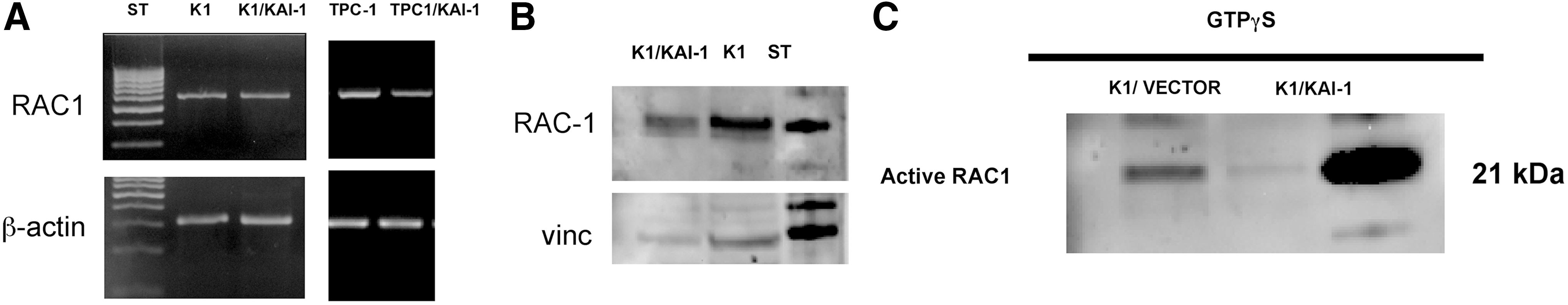
RAC-1 mRNA, Rac-1 protein, and Rac-1 activation in KAI-1/CD82–transfected and vector-transfected TPC cells. Re-expression of KAI-1/CD82 is associated with slightly reduced levels of RAC-1 mRNA and Rac-1 protein in KAI-1/CD8–transfected K1 and TPC-1 cells, as demonstrated by real-time polymerase chain reaction
KAI-1/CD82–transfected TPC cells exhibit a reduced capacity to metastasize in vivo
We evaluated the effect of CD82/KAI-1 on experimental metastasis. To this purpose two groups of four mice each were inoculated into the tail vein (intravenous) and killed 60 days after tumor injection, and the presence of lung metastases was analyzed at macroscopic and microscopic levels, by the aid of a dissecting and optical microscope. Lung metastases evaluated by immunohistochemical analysis were observed in all mice carrying vector-K1 tumors (100%), with a median number of 4.5 metastases per lung (mean 9.6±2.1), while only 2 out of 4 (50%) animals injected with KAI-1/CD82 cells show lung metastases with a median number of 0.5 metastases per lung (mean 2.5±1.9). The difference was statistically significant (p=0.016). Smaller metastatic lesions were observed in animals injected with CD82/KAI-1 cells (diameter from about 0.065 to 0.7 mm) when compared to lung nodules found in control mice (diameter from about 0.050 to 3.7 mm). Representative images of the macroscopic and microscopic lung lesions are reported in Figure 8.

Lung metastasis assay: representative images of the lung nodules in the animals injected with cells expressing KAI-1/CD-82 or control vectors.
Discussion
In the present study, we provide evidence that most cases of TPC express low levels of KAI-1/CD82, that poor expression of KAI-1/CD82 is more often present in those cases with extrathyroid extension of the disease, and that re-expression of KAI-1/CD82 in TPC cells is associated with decreased migratory and invasive capacity of tumor cells. So far, the role of KAI-1/CD82 has been poorly investigated in thyroid tumors. In an immunohistochemical study with a polyclonal antibody, it was found that KAI-1/CD82 was overexpressed in 64% of papillary carcinoma cases (25). In another study, it was reported that 33 cases of TPC had significantly lower levels of KAI-1/CD82 RNA as compared with 12 cases of benign goiter (26). In both studies, low expression of KAI-1/CD82 was associated with lymph node metastasis, capsule infiltration, and a more advanced tumor–node–metastasis status (25,26). Our findings are consistent with these previous reports since we have observed that 64% of TPC tumors had lower levels of KAI-1/CD82 as compared with the corresponding normal thyroid tissue, and that low expression of KAI-1/CD82 is more often associated with extrathyroid extension of the disease. Thus, thyroid tumors appear to be similar to those of other organs, including prostate, lung, breast, and pancreas, where decreased expression of KAI-1/CD82 is often associated with increased invasiveness and with a less favorable prognosis (5 –10).
In a previous study (23), we have reported that HGF stimulation of TPC cells is followed by upregulation of COX-2 in vitro, and that expression levels of MET RNA and COX-2 RNA are directly correlated in tissues involved by TPC. In the present study, we provide evidence that COX-2 inhibition is followed by upregulation of KAI-1/CD82, and that the expression levels of COX2 and KAI-1/CD82 are inversely correlated in tumor tissue. Taken together, our findings suggest that HGF activation of tumor cells overexpressing Met receptor causes upregulation of COX-2 and downregulation of KAI-1/CD82. Since COX-2 and KAI-1/CD82 have opposite effects on tumor invasiveness, it is not surprising that these two molecules are inversely coregulated perhaps by a common mechanism HGF induced. The pro-invasive role of COX-2 has been demonstrated in humans and mice (27 –30). In fact, tumor cells overexpressing COX-2 have increased matrix metalloproteinase activity; moreover, the COX-2 product PGE2 stimulates tumor invasiveness via PI3K signaling, and transactivates Met-receptor; this latter event causes an increased accumulation of β-catenin in the nucleus, and increased expression of urokinase-type plasminogen activator receptor on the cell membrane (31 –34). The relevance of COX-2, Met, and nuclear β-catenin in regulating invasiveness is confirmed by their higher expression at the tumor front of invasive colorectal carcinoma (32) and TPC (35).
The molecular mechanisms determining the downregulation of KAI-1/CD82 in tumor cells were previously investigated. It has been shown that poor expression of KAI-1/CD82 in prostate cancer cells is related to the inhibitory action of beta-catenin–reptin complexes on KAI-1/CD82 gene transcription, and is antagonized by Tip60 (36). Other authors have demonstrated that the SUMOylation status of the reptin–chromatin-remodeling complex modulates KAI-1/CD82 transcription and decreases the invasive capacity of tumor cells (37). Reduced expression of KAI-1/CD82 may also derive from increased degradation of the protein, as a consequence of binding to ubiquitin ligase gp78 (38). The existence of a possible linkage between KAI-1/CD82 and Met receptor was investigated in carcinoma cell lines derived from lung (18), prostate (15), and pancreas (20). In all these studies, it was found that re-expression of KAI-1/CD82 had an anti-invasive effect on tumor cells, and that this effect was associated with a lower degree of Met-phosphorylation upon HGF stimulation. Negative interaction of KAI-1/CD82 with Met-induced cell migration is also supported by the demonstration of inactivation of small GTP-binding proteins of the Rho family, including RAC-1; these latter are on the downstream signaling of HGF–Met interaction and are involved in the organization of the actin cytoskeleton and in epithelial cell spreading (18). Our results are consistent with these previous observations since we have observed that TPC cells re-expressing KAI-1/CD82 exhibit decreased migratory and invasive capacity in vitro, are poorly responsive to HGF stimulation, and show low levels of RAC-1 activation. Moreover, they may indicate that there is a common molecular pathway regulating the action of KAI-1/CD82 in Met-positive epithelial tumors independently of the organ of origin.
The effect of KAI-1/CD82 has been investigated in numerous xenograft studies using metastatic cell lines, and it has been found that expression of the molecule significantly reduced the rate of metastasis without affecting primary tumor growth (4,11,12,36,38). We have investigated the functional role of K1-transfected cells in vivo using nu/nu mice. We have found that mice inoculated intravenously with K1-transfected cells developed less and smaller lung metastasis than control mice. Thus, the in vivo experiments confirm that KAI-1/CD82 manipulation in K1 cells profoundly interferes with the tumorigenic capacity.
Indeed, there are several reports indicating that KAI-1/CD82 may interfere with motility and invasion of tumor cells through an action on beta1 or beta3 integrins (13 –16). Depending on the tumor system, cells re-expressing KAI-1/CD82 were more or less adhesive to fibronectin, collagen, or vitronectin; the different effects were caused by quantitative alterations in the levels of beta-1 or beta-3 integrins expressed at membrane level. We have observed that KAI-1/CD82–transfected K1 cells were significantly less adhesive to fibronectin than vector-transfected cells (data not shown). Thus, this feature or perhaps others may interfere with tumor cell seeding in the mouse tissues and hence may reduce or delay the chance of tumor development at the injection site.
In conclusion, we provide evidence that re-expression of KAI-1/CD82 strongly limits the invasive capacity of PTC cells, and that poor expression of this molecule is controlled by COX-2, and perhaps by other molecules involved in the HGF/Met-mediated pathway of invasiveness. Our findings may contribute to the understanding of the high diffusive potential of papillary carcinoma of the thyroid, a slowly growing tumor characterized by a high incidence of lymph node metastasis even in the early stages of the disease.
Footnotes
Acknowledgment
This work was supported by the Italian Association for Cancer Research (to D.D.B.).
Author Disclosure Statement
There are no financial or other relationships that may lead to a conflict of interest.