
Abstract
Background:
Estrogens may enhance thyroid cancer cell growth. We have recently reported that a novel isoflavone-derived anti-estrogenic compound developed in our laboratory, the N-t-boc-hexylenediamine derivative of 7-(O)-carboxymethyl daidzein (cD-tboc), can induce apoptosis and retard growth in human thyroid carcinoma cell lines through inhibitory interaction on estrogen receptor β. Here we tested the hypothesis that cD-tboc can likewise retard cell growth in cultured human thyroid papillary carcinoma cells, normal thyroid cells, and goiter cells removed during thyroidectomy.
Methods:
In vitro experiments in cultured human thyroid normal, goiter, and papillary thyroid carcinoma (PTC) cells were performed. Estrogen receptors α and β (ERα and ERβ), DNA synthesis and creatine kinase (a marker of estrogenic genomic response), and the effects of cD-tboc on DNA synthesis in cultured human PTC cells were assessed.
Results:
First, all cell types thus harvested and grown in culture expressed both ERα and ERβ, with a variably higher abundance of ERβ over ERα seen in the goiter and PTC cells, but not in the normal thyroid cells. Second, DNA synthesis and creatine kinase were increased in response to estradiol-17β (E2), the ERα agonist propyl-pyrazole-trisphenol as well as the ERβ agonist diarylpropionitrile. Third, cD-tboc dose-dependently inhibited DNA synthesis in cultured human PTC cells (−65%) and to a lesser extent in goiter cells (∼−30%).
Conclusion:
This study provides the first evidence that cD-tboc can act to inhibit growth in primary cultures of human PTC cells and goiter cells removed during thyroidectomy. Whether this can be utilized for the treatment of human thyroid cancer and/or goiter remains to be explored.
Introduction
Daidzein is a naturally occurring phytoestrogen of the isoflavone family that displays some estrogenic properties in a large number of cell types (12 –14). We have previously generated a synthetic derivative of daidzein [7-(O)-carboxymethyl daidzein conjugated to N-t-Boc-hexylenediamine, hence cD-tboc] and reported that it can induce apoptosis in several cancer cell lines expressing estrogen receptor β (15–16). In a study reported this year we observed that cD-tboc induces apoptotic cell death in several cell lines of human thyroid carcinoma, such as WRO (17). Here we tested the effect of cD-tboc on cell growth in thyroid carcinoma cells as well as in unaffected adjacent normal thyroid cells, harvested during total thyroidectomy for thyroid cancer. Human thyroid goiter cells from tissue obtained during thyroidectomy from patients with benign colloid goiter were also examined.
Methods
Preparation of cells
The use of human thyroid tissue for this study was approved by the institutional Helsinki committee. Thyroid tissue removed during total thyroidectomy for thyroid cancer in three patients was inspected by a pathologist who, following proper dissection and measurements, provided some of the cancerous tissue and some of the unaffected normal thyroid tissue for further processing for this study. Histological examination of the cancerous tissue confirmed the diagnosis of papillary thyroid carcinoma (PTC) in these specimens. Cells were prepared as described previously (18–19) with slight modifications. Tissue fragments were immediately placed in cold phosphate-buffered saline (PBS) containing antibiotics (2% penicillin/streptomycin/amphotericin) in a sterile tissue culture dish. They were further cut into small pieces which were washed three times with PBS by centrifugation at 200 g for 12 min to get rid of blood. The precipitated tissue fragments and cells were then suspended in an RPMI 1640 medium containing 10% fetal bovine serum (FBS), 2 mM L-glutamine, 1% Na pyruvate, and 1% MEM-Eagle nonessential AA containing antibiotics (0.1 mg/mL streptomycin, 100 U/mL penicillin, and 0.025 mg/mL amphotericin B) and placed in tissue culture flasks. The larger fragments were placed on tissue culture dishes, dried of the PBS solution, and then allowed to adhere to the dishes, after which the tissue culture medium was added. Fragments were kept for about a week until cell migration was evident. Upon confluence, cells were trypsinyzed and re-seeded for the different experiments. Normal cells were used only at first and second passages, while cancer cells were used up to four passages. The method of preparation of benign human thyroid cells from tissue obtained at thyroidectomy from patients with benign colloid goiter has been previously reported in detail (18,20). In the present study, cells of three patients with benign colloid goiter were released in and maintained under conditions developed by us for the culture of human vascular smooth muscle cell growth (21).
mRNA expression
Thyroid transcription factor 1 (TTF-1) and PAX-8 expression in the thyroid tissue was assessed by semi-quantitative polymerase chain reaction (PCR) as recently described by us (17) and normalized for cyclophylin expression. PCR products were sent for sequencing at Hy-labs (Rehovot, Israel). The expression of estrogen receptors α and β (ERα and ERβ) was assessed as previously reported from this laboratory by real time PCR (17) with an ABI 7700 Real Time PCR System, using specific primer probe sets obtained from Applied Biosystems (Foster City, CA).
Assessment of DNA synthesis, cell proliferation and apoptosis
To assess DNA synthesis, cells were treated with various hormones or agents for 24 hours as indicated. At the end of incubation, [3H]-thymidine was added for 2 hours. Cells were then treated with 10% ice-cold trichloroacetic (TCA) for 5 minutes and washed twice with 5% TCA and then with cold ethanol. The cellular layer was dissolved in 0.3 mL of 0.3 M NaOH and aliquots were taken for counting radioactivity and [3H]-thymidine incorporation into DNA was calculated. Cell proliferation was assayed using the cell proliferation kit based on the XTT colorimetric assay of Biological Industries (Kibbutz Beit Haemek, Israel) as previously described by us (17). Apoptosis was assessed with a photometric enzyme-immunoassay for the quantitative in vitro determination of cytoplasmic histone-associated-DNA-fragments (mono-and oligo-nucleosomes; Cell Death Detection ELISA plus kit, Roche kit Molecular Biochemicals). This is a sandwich assay constructed to identify DNA fragments through the use of two antibodies, one against histones and the second directed against DNA.
Assessment of creatine kinase-specific activity
Supernatant extracts from homogenized cells collected from the culture dishes with an extraction buffer were obtained by centrifugation of homogenates at 14,000 g for 5 minutes at 4°C in a Hermeles micro centrifuge. Creatine kinase (CK)–specific activity was assayed by a coupled spectrophotometric assay (15,16). Protein was determined by Coomasie blue dye binding using bovine serum albumin as the standard, and the activity was defined as specific activity: nmol/min/mg protein.
Statistical analysis
The significance of differences between the mean values obtained from experimental and control cultures was evaluated by the unpaired two-tailed Student's t-test, and by analysis of variance. A p-value of <0.05 was considered significant.
Results
Estrogen receptors are expressed in normal and human thyroid carcinoma cells
TTF-1 and PAX8 were expressed in the carcinoma cells grown from the three specimens, validating the thyroid nature of the cells (data not shown). Cancerous as well as normal thyroid cells, grown from either the normal tissue of the thyroid gland removed during total thyroidectomy for thyroid cancer or from thyroid goiter tissue, expressed ERα and ERβ (data not shown). However, there was some excess of ERβ expression relative to ERα both in thyroid cancer tissue and in goiter cells grown from patients free of thyroid cancer (ERβ/ERα=1.36; 1.33, respectively), relative to the ERβ/ERα ratio in normal thyroid tissue removed from patients with thyroid cancer (ERβ/ERα=0.27; p<0.01).
Effect of E2 and specific ERα and ERβ agonists on human thyroid carcinoma and goiter cell proliferation
As shown in Figure 1, estradiol-17β (E2; 30 nM) elicited a significant increase in [3H]-thymidine incorporation in normal thyroid cells (Fig. 1a), human thyroid carcinoma cells (Fig. 1b), and goiter cells (Fig. 1c). Such increments were also seen when these cells were exposed to the ERβ agonist diarylpropionitrile (DPN; 420 nM). In contrast, a proliferative response to the ERα agonist propyl-pyrazole-trisphenol (PPT; 390 nM) was seen in carcinoma and goiter cells, but not in normal thyroid cells. E2 also increased CK activity, a marker of the genomic effect of estrogen [formerly known as estrogen-induced-protein (21)] in all three cell types and the response was seen with both E2 per se and the ERα- and ERβ-selective agonists PPT and DPN, respectively. Due to limited tissue availability, single concentrations of each agent were applied. In each case, the concentration chosen was previously shown by us to elicit the maximal responses in thyroid carcinoma cell lines (17).
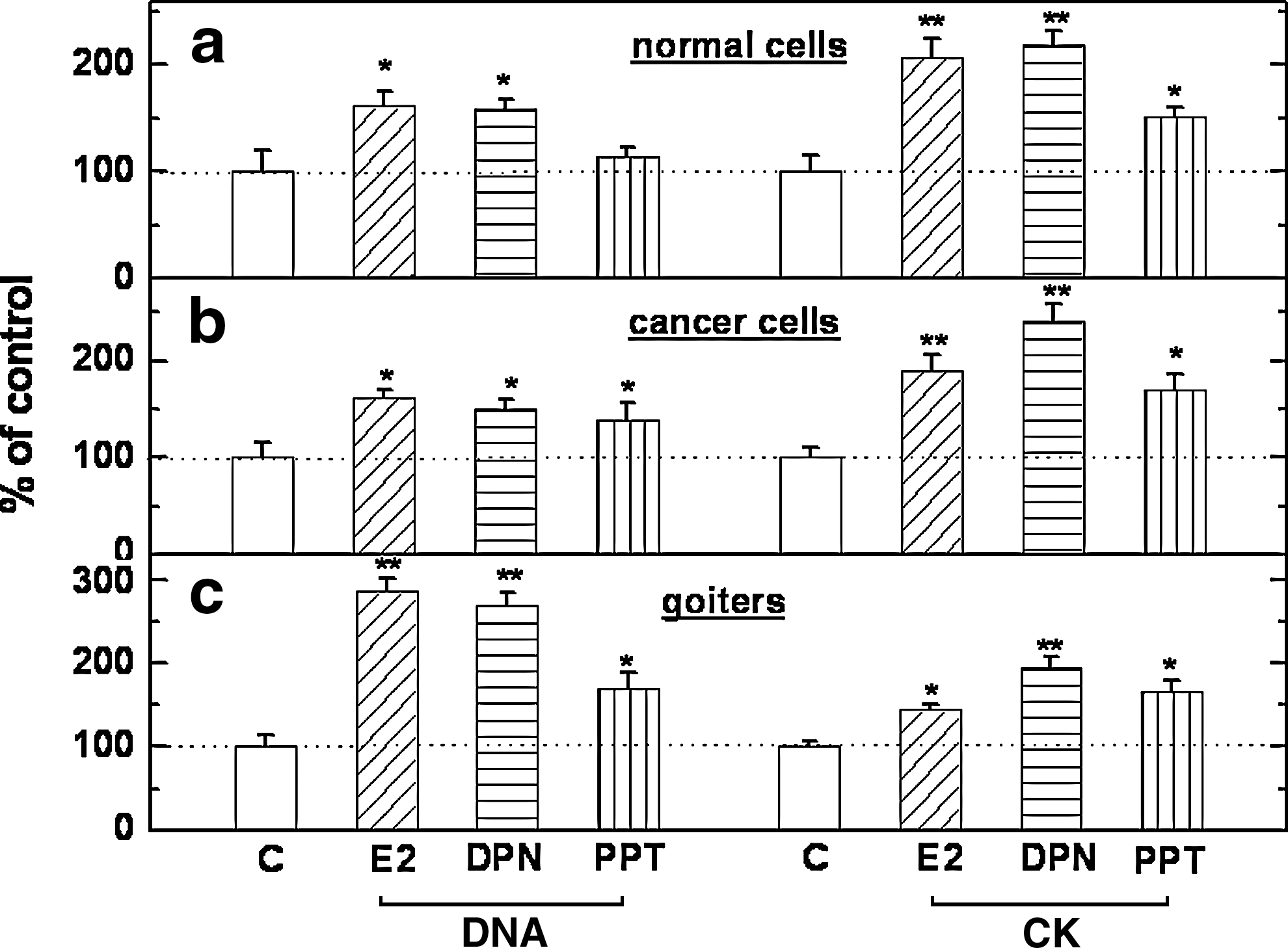
Effect of E2 (30 nM), DPN (420 nM), and PPT (390 nM) on DNA synthesis (the left four bars in each panel) and CK-specific activity (the right four bars in each panel). Cultured human thyroid normal cells and thyroid carcinoma cells from three patients
Effect cD-tboc on human thyroid cancer cell growth and survival in vitro: interaction with ERα and ERβ antagonists
Figure 2 illustrates the effect imparted by different concentrations of cD-tboc on DNA synthesis in cultured thyroid cancer cells, normal thyroid cells from the same three patients, and goiter cells from three patients with benign colloid goiter. As shown, cD-tboc at submicromolar to micromolar concentrations inhibited [3H]-thymidine incorporation by ∼70% in cancer cells and ∼30%–35% noncancerous human thyroid cells. Further, as shown in Table 1, the inhibitory effect of cD-tboc was significantly attenuated by the the ERβ antagonist phenol trifluoromethyl pyrazolo-pyrimidine phenol (PTHPP; 150 nM), but not by the ERα antagonist 1,3bis(4-hydroxyphenyl)-4-methyl-5[4-(2)-piperidinyletoxy)phenol]-1H-pyrazole dihydrochloride (MMP; 150 nM), thus suggesting that cD-tboc inhibits 3[H]-thymidine incorporation in normal thyroid and PTC cells via interaction with ERβ. Because of the limited supply of thyroid cells from the three patients undergoing total thyroidectomy for thyroid carcinoma and because of the similar ERα- and ERβ-mRNA expression and response profile to E2 and cD-tboc of thyroid carcinoma and goiter cells, effects of cD-tboc were further explored in goiter cells, which were abundantly retrieved from patients with large, nonmalignant goiters. We observed an almost 50% reduction in cell number and a twofold increase in apoptotic cell death as detected by a Roche assay which quantifies histone-DNA particles (Table 2).
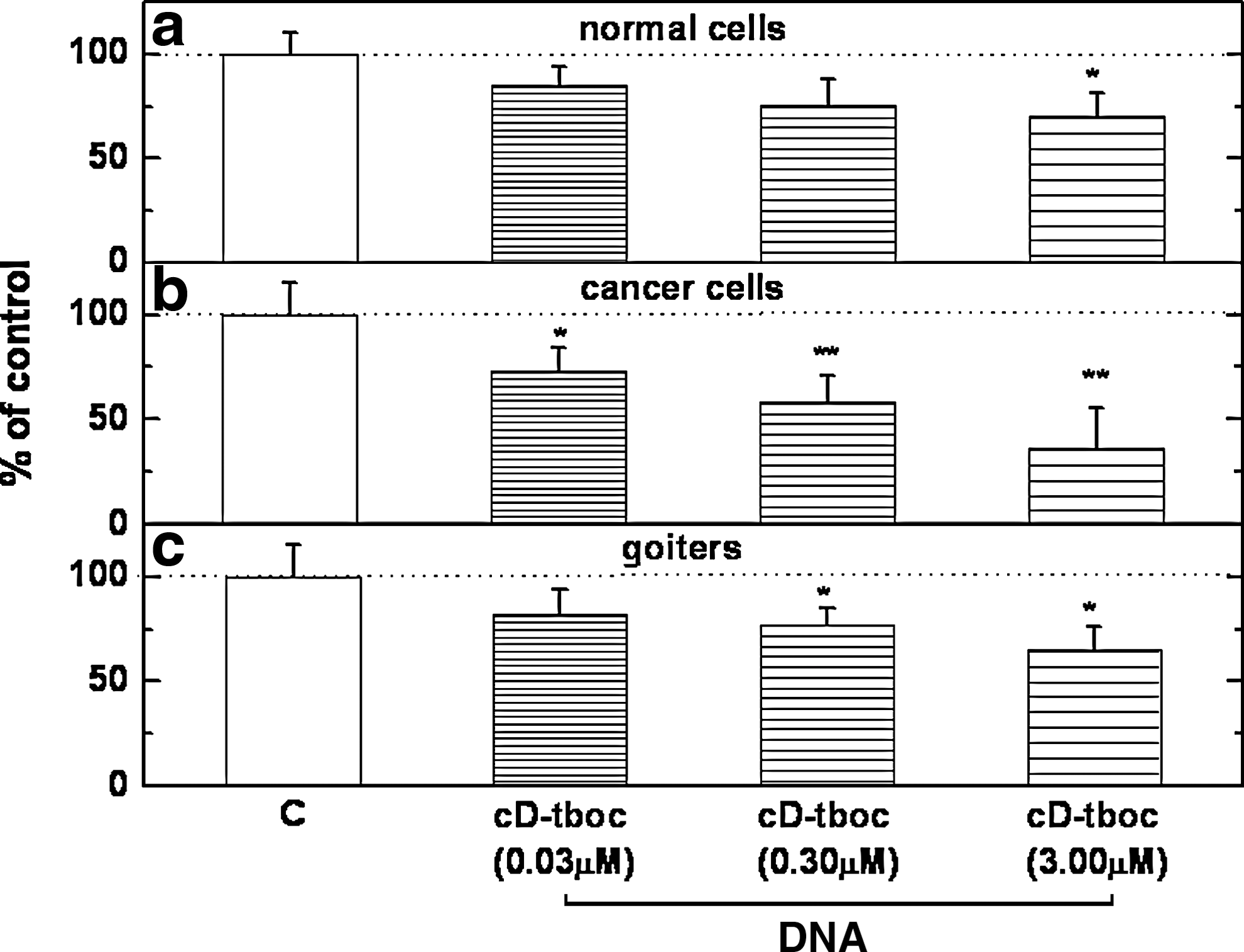
Effect of cD-tboc on DNA synthesis (the right three bars in each panel) in cultured human thyroid normal cells and thyroid carcinoma cells from three patients
cD-tboc was used at a concentration of 3 μM; the ERα antagonist MMP and the ERβ antagonist PTHPP were used at a concentration of 150 nM each. Basal 3[H]-thymidine incorporation was 776±19 dpm/well in the normal thyroid cells and 870±37 dpm/well in the PTC cells (n=3 for each treatment; * p<0.05).
PTC, papillary thyroid carcinoma; MMP, 1,3bis(4-hydroxyphenyl)-4-methyl-5[4-(2)-piperidinyletoxyphenol]-1H-pyrazole dihydrochloride; PTHPP, phenyl trifluoromethyl pyrazolo-pyrimidine phenol; cD-tboc, 7-(O)-carboxymethyl daidzein conjugated to N-t-Boc-hexylenediamine; ERα, estrogen receptor α; ERβ, estrogen receptor β.
Cultured human thyroid goiter cells were treated for 24 hours with different concentration of cD-tboc for the changes in cell replication as measured by the XTT assay (% of control of the mean absorbance at a wavelength of 490 nm after subtraction of nonspecific absorbance measured at 650 nm). Details are given in the Methods. * p<0.05 determined by ANOVA, for the different concentrations used.
Apoptotic cell death after treatment with cD-tboc was assayed in cultured human thyroid goiters that were treated for 24 hours with different concentrations of cD-tboc for the changes in apoptotic cell death as assessed by the detection histone–DNA fragments (% of change in treated goiter thyroid cells compared to control). Details are given in the Methods. † p<0.001 determined by ANOVA, for the different concentrations used.
ANOVA, analysis of variance.
Discussion
Since estrogens may enhance thyroid cancer growth in human subjects, which could explain the high risk for thyroid cancer in women (6,8 –11), it is only plausible that negative modulators of estrogen receptor function could have a potential value as anti-thyroid carcinoma agents. We have recently generated a novel synthetic derivative of the phytoestrogen daidzein, cD-tboc, which showed anti-cancer properties in ovarian, colon, and thyroid cancer cell lines. The anti-cancer effect cD-tboc in human thyroid carcinoma cell lines involved the induction of apoptosis and was ERβ-mediated as it was strongly attenuated in the presence of a specific ERβ antagonist, PHTPP, and in cells in which the expression of ERβ was nearly eliminated (by 98.6%) using ERβ siRNA (17). These earlier results are further confirmed in this study in cultured human normal thyroid and PTC cells (Table 2), as the inhibitory effect of cD-tboc was likewise blocked in the presence of an ERβ antagonist, but not an ERα antagonist.
Here we report, for the first time, that cD-tboc can exert a growth inhibitory effect in cultured thyroid carcinoma cells removed from three human subjects undergoing total thyroidectomy. * Such effect is achievable at the submicromolar- to the micromolar range with a clear log dose relationship between the concentration of cD-tboc and the growth attenuation. At the lowest effective concentration this would be rather comparable to the plasma concentrations of the mother compound of cD-tboc, daidzein, measured in human subjects consuming regular diet (22,23). Also, the in vivo administration of cD-tboc to nude mice that were subcutaneously preinoculated with two types of cancer cells was not associated with toxic effects in two separate studies.
The results presented here in thyroid carcinoma cells harvested from human subjects diagnosed with PTC are consistent with our recently reported study of the effect of cD-tboc in human thyroid cancer cell lines, such as WRO (17). Because current modes of treatment for human differentiated thyroid cancer leave some patients with incurable disease, the observation that cD-tboc can inhibit growth of human thyroid cancer cells in vitro should be followed by more extensive studies to explore its potential role in this subpopulation of thyroid cancer patients. Finally, the growth attenuating effect of cD-tboc in goiter cells seen here is of potential interest and merits confirmation in larger scale studies. Presently, the medical treatment of large goiters is rather limited. Hence, the possibility that cD-tboc (which was able to lessen cultured goiter cell population in the present report) can also reduce goiter size in humans should be explored.
Footnotes
Disclosure Statement
The authors declare that no competing financial interests exist.
*
Four additional patients with carcinoma were also studied, but in the interest of brevity, their data are not reported in this article. These patients showed essentially the same response.