
Abstract
Background:
Studies in men are not consistent regarding the effects of thyroid hormone on the production of gonadotropins. In hypothyroidism consequent to diverse causes, an increase or no change in serum luteinizing hormone (LH) have been reported. The attempt to explain the mechanisms involved in this pathology using rats as an experimental model also seems to repeat this divergence, since hypothyroidism has been shown to induce hypogonadotropic hypogonadism, a hypergonadotropic state, or not to affect the basal levels of LH. Notably, the promoter region of the gene encoding the Lh beta subunit and GnRH (gonadotropin-releasing factor) does not contain a thyroid responsive element. Therefore, we investigated the hypothesis that, in male rats, posttranscriptional mechanisms of LH synthesis are altered in hypothyroidism. We also attempted to determine if hypothyroidism directly affects testicular function in male rats.
Methods:
Male Wistar rats, 60 days old, were thyroidectomized or sham-operated. After 20 days, they were decapitated, and the pituitaries were collected and analyzed for Lh mRNA, LH content, poly(A) tail length, and polysome profile. The testes were collected and analyzed for Lh receptor mRNA, LH receptor content, and histology using morphometric analyses. The testis, epididymis, seminal vesicle, and ventral prostate were weighed, and serum concentrations of LH, testosterone, thyrotropin (TSH), and triiodothyronine (T3) were measured.
Results:
Hypothyroidism was associated, in the pituitary, with an increase in Lh mRNA expression, a reduction in Lh mRNA poly(A) tail length, a reduction in the number of LH transcripts associated with polysomes. Pituitary LH was decreased but serum LH was increased from 102 to 543 pg/mL. Despite this, serum testosterone concentrations were decreased from 1.8 to 0.25 ng/mL. A decreased germinative epithelium height of the testes and a reduced weight of androgen-responsive tissues were observed (ventral prostrate: 74 vs. 23 mg/100 g body weight [BW]; seminal vesicle undrained: 280 vs. 70 mg/100 g BW; and seminal vesicle drained: 190 vs. 60 mg/100 g BW).
Conclusions:
Hypothyroidism in adult male rats has dual effects on the pituitary testicular axis. It alters posttranscriptional mechanisms of LH synthesis and probably has a direct effect on testicular function. However, these data suggest the possibility that reduced LH bioactivity may account in part for impaired testicular function.
Introduction
The attempt to explain the mechanisms involved in this pathology using rats as an experimental model also repeat the divergent observations in men, with the exception of the sperm parameters, which, as far as we know, have not been evaluated yet. Thus, hypothyroidism has been shown to induce hypogonadotropic hypogonadism or a hypergonadotropic state, or to have no effect on the basal levels of luteinizing hormone (LH) in rats (2). But, exogenous administration of GnRH in hypothyroid men (3) or male rats (4) does not result in a release of pituitary gonadotropins, indicating the inability of the gonadotropes to respond to this stimulus.
In order to explore the molecular mechanisms underlying these alterations, we subjected adult male Wistar rats to a hypothyroid state, and evaluated posttranscriptional events associated with LH biosynthesis. Specifically, we evaluated serum concentrations of LH, pituitary LH mRNA, and protein expression, as well as LH mRNA translational efficiency. We also evaluated the expression of the LH receptor (LHR) in the testes, the serum testosterone concentrations, the weights of the androgen-dependent reproductive organs, and the morphology of the seminiferous epithelium.
Materials and Methods
Chemicals
Methylmercaptoimidazole (MMI) was purchased from Sigma-Aldrich (St. Louis, MO); Platinum SYBR green qPCR SuperMix UDG, oligo-dT, deoxyribonucleotide triphosphates (dNTP), bovine serum albumin (BSA), protein molecular weight markers, 100 bp DNA ladder, and mercaptoethanol were purchased from Life Technologies Corp. (Camarillo, CA); M-MLV Reverse Transcriptase and GoTaq Flexi DNA Polymerase were purchased from Promega Corp. (Madison, WI); the anti-LH antibody was obtained from the National Hormone and Pituitary Program (Torrance, CA); anti-GAPDH, anti-LHR (H-50), and secondary peroxidase-conjugated antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA); the enhanced chemiluminescence kit was purchased from Amersham Biosciences (Little Chalfont, United Kingdom); Coat-A-Count Total Testosterone and total triiodothyronine (T3) kits were purchased from Siemens (Deerfield, IL); hybridization nitrocellulose membrane and Luminex xMAP technology for rat LH were purchased from Millipore Corp. (Milliplex MAP rat pituitary panel, Billerica, MA).
Animals and treatments
Male Wistar rats, 60 days old (250–300 g), were obtained from our breeding colony, and maintained on rat chow ad libitum, under a 12:12 hour dark/light cycle at controlled room temperature (23±1°C). They were randomly assorted into euthyroid (sham-operated animals receiving water ad libitum) and hypothyroid groups [surgically thyroidectomized, receiving methylmercaptoimidazole (MMI) 0.03% and CaCl2 0.05% in their drinking water for 20 days, as previously described (5 –7)]. All procedures were in accordance with the Brazilian College of Animal Experimentation guidelines and approved by the Institute of Biomedical Sciences, University of Sao Paulo—Ethical Committee for Animal Research (protocol 029/55/02).
Real-time polymerase chain reaction for Lh and Lhr
Total RNA was extracted from the pituitaries and testes according to the guanidine–phenol–chloroform method (8) and submitted to real-time polymerase chain reaction (PCR) as previously described (9). Briefly, 1.0 μg of total RNA was reverse-transcribed at 65°C for 10 min using 0.5 μg oligo-dT and diethyl pyrocarbonate–treated (DEPC) water in a total volume of 5.0 μL. Subsequently, 25 U RNAse Out, 4.0 μL buffer (250 mM Tris-HCl pH 8.3; 375 mM KCl, 15 mM MgCl2, 50 mM dithiothreitol), 1.0 μL dNTP (10 mM), 200 U M-MLV Reverse Transcriptase and DEPC water were added up to 20 μL, following incubation for 60 min at 42°C. Quantification of Lh beta and Lhr mRNA was performed by real-time PCR using the Rotor-Gene termocycler (Corbett Life Sciences, Sydney, Australia). The reaction was performed with 1.0 μL of cDNA, 0.4 μM of specific primers, 5.0 μL of Platinum SYBR Green qPCR SuperMix UDG and DEPC water for a total volume of 12.5 μL. Quantification was performed by the ΔΔCt method, as previously described (10,11). The Rpl19 constitutive gene was used as an internal control. The primers are shown in Table 1.
F, forward; R, reverse.
Western blotting analysis for Lh
Pituitaries were individually homogenized using a Politron in 300 μL of homogenization buffer (0.25 M sucrose, 2 mM MgCl2, 20 mM Tris-HCl), as previously described (9), and the supernatant was used to determine total protein concentration (12). Briefly, 30 μg of protein were dried in a Speed Vack®, resuspended in 10 μL of Laemmli buffer (62.5 mM Tris-HCl pH 6.8; 20% glycerol; 10% 2-mercaptoethanol; 4% SDS; 0.08% bromophenol blue) plus 20 μL of homogenization buffer, submitted to SDS-PAGE 15% (electrophoresis buffer: Tris-base 25 mM, 193 mM glycine, 0.1% SDS) and transferred to hybridization nitrocellulose membrane through a semi-dry blotting system (Bio-Rad Laboratories Inc.) at 15 mV for 60 min (transfer buffer: Tris-base 25 mM, 192 mM glycine, methanol 20%). The membrane was preincubated with blocking buffer (5% nonfat dry milk, phosphate-buffered saline, and 0.1% Tween 20) to reduce nonspecific binding overnight at 4°C and subsequently incubated for 3 h at room temperature (RT) with anti-Lh antibody (1:1500) or anti-GAPDH antibody (1:1000) diluted in blocking buffer. Immunocomplex Band detection was performed using an appropriate peroxidase-conjugated secondary antibody (1:5000) diluted in the same primary antibody buffer for 1 h at RT and submitted to the Enhanced Chemiluminescence (ECL) kit (Amersham Biosciences). Blots were analyzed with Image J 1.42q software (National Institutes of Health [NIH]). GAPDH or alpha actinin was used as an internal control.
Western blotting analysis for Lhr
The testes were pulverized in liquid nitrogen and dounce-homogenized in RIPA buffer containing protease inhibitors, as previously described (13). One hundred micrograms of total protein was diluted in 20 μL of Laemmli buffer and submitted to SDS-PAGE 8% in the same conditions as previously described for LH. The membrane was preincubated with blocking buffer (3% BSA, 5 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.5% Tween 20 and 0.5% NP-40) for 2 h at RT and subsequently incubated overnight at 4°C with anti-LHR antibody (1:200) diluted in 1% BSA, 5 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.5% Tween 20 and 0.5% NP-40. Two bands corresponding to the immature and mature forms of the receptor were quantified separately. After immunocomplex detection, the membrane was stripped (14) and incubated with an anti-alpha actinin antibody (1:1000) in the same conditions as for the anti-LHR (internal control).
Polysome profile by sucrose gradient
A previously described protocol (15) was adapted (6). In brief, a pool of 10 pituitaries was homogenized in 0.6 mL lysis buffer containing 15 mM Tris–HCl (pH 7.4), 25 mM MgCl2, 300 mM NaCl, 1% Triton X-100, 0.1 mg/mL cycloheximide, 1 mM dithiothreitol, and 10 mg/mL heparin. After removal of cell debris by centrifugation at 10,000 g for 10 min at 4°C, the nucleic acid concentration of the supernatant was quantified, and 14 AU (absorbance unit) were loaded onto a 10 mL sucrose gradient (7–47% w/v) and centrifuged at 114,817 g (39,000 rpm) for 150 min (SW41 Ti rotor, Beckmann). The UV absorbance of the resulting gradients was monitored at 254 nm (Monitor UVis-920; GE HealthcareBio-Sciences AB, Uppsala, Sweden). The gradients were collected into 11 fractions of approximately 1 mL each for RNA isolation, immediately frozen on dry ice, and then stored at −70°C. For statistical analysis, fractions 6 through 11 were classified as the polysome region and polysomal RNA was isolated from these fractions.
Isolation of polysomal RNA
The isolation of polysomal RNA was performed, as described previously (6). Briefly, 1 mL from each polysomal fraction was mixed with 2 mL guanidine thiocyanate 4 M and 3 mL absolute ethyl alcohol, kept overnight at −70°C and centrifuged at 10,000 g for 20 min at 4°C for RNA precipitation. RNA pellets were washed with 70% ethyl alcohol and dissolved in DEPC water, at 65°C, for 15 min. Then, 2 mM sodium acetate, ethyl alcohol (1:0.5 v/v), and 45 U of heparin were added to the samples, which were centrifuged at 15,700 g (13,000 rpm) for 20 min at 4°C (5415R—Eppendorf ). The quantification of mRNA of LH was the same described for total mRNA by real-time PCR.
Poly(A) tail Lh mRNA analysis
The assay was performed by rapid amplification of cDNA ends poly(A) test (RACE-PAT), as previously described (7). Two micrograms of RNA were preincubated with 200 ng of an oligo-dT anchor (5′-GCG AGC TCC GCG GCC GCG-T12) for 5 min at 70°C, followed by 1 h at 42°C in a mix containing 10 mM of each dNTP, 5× first-strand buffer, 0.01 M of dithiothreitol, and 1 μL of 200 U/μL of reverse transcriptase MMLV. Three micrograms of reverse transcriptase reaction product were added to a reaction containing 25 mM MgCl2, 10 mM of each dNTP, 25 pmol of each primer (LH: 5′-CCA ATG ACC TGT GAC CTT CC and anchor: 5′-GCG AGC TCC GCG GCC GCG-T12), 1.25 U of GoTaq Flexi DNA Polymerase, and 5× Green GoTaq Flexi Buffer Migration Pattern. PCR was performed using the following conditions: 5 min at 95°C, followed by 40 cycles of 30 sec at 95°C, 1 min at 68°C, 1 min at 72°C, and ending with a 7 min final extension at 72°C. The PCR products were electrophoretically analyzed using an ethidium bromide-stained 2.5% agarose-TAE gel. The top of the smear indicated the longest amplified fragment, which represents the poly(A) tail size plus 128 bases upstream, according to the primer used. The changes in poly(A) tail length were detected by analyzing the length of the cDNA smears obtained in both groups. The sample analysis was estimated by densitometry and compared with a 100 bp DNA ladder using the ImageMaster 1D-Pharmacia Biotech SW software (Pharmacia Biotech, Uppsala, Sweden).
Morphometry of seminiferous epithelium
The testes were fixed in Bouin's solution for 8 h, treated with alcohol, embedded in paraffin, and cuts of 5 μm were prepared as stained laminas with hematoxylin and eosin. A magnification of 100× was used for analysis of linear morphometry from the seminiferous tubules by determining the tubular diameter, seminiferous epithelium, and luminal diameter, as previously described (16).
Reproductive organ weights
The testes, epididymis, ventral prostate, and seminal vesicle were weighed, and the values converted to relative weights as mg/100 g of body weight (BW). The seminal vesicle was weighed with fluid (undrained) and after fluid removal (drained).
Hormone measurements
The serum hormone concentrations were measured by radioimmunoassay using commercial kits for total testosterone and T3 (Coat-A-Count, DPC, Los Angeles, CA). The minimum sensitivity was 0.008 ng/mL for testosterone and 0.41 ng/mL for T3. The intrassay coefficient was <4.97% for testosterone and <4.7% for T3. The serum LH measurement was determined by chemiluminescence immunoassay using the Luminex xMAP kit technology (Milliplex MAP rat pituitary panel). The minimum sensitivity was 2.91 pg/mL for LH and the intraassay coefficient was <2.5%. The thyrotropin (TSH) assay was performed by a specific radioimmunoassay using rat TSH for iodination and preparation of the standard curve. The minimum sensitivity was 0.18 ng/mL and the intrassay coefficient was <4.3%.
Effectiveness of the treatment
To evaluate the hypothyroid status of animals, the TSH and T3 serum concentrations, as well as the heart weight, were determined and compared to euthyroid animals, as described previously (5 –7).
Statistical analysis
The variables under study were first submitted to tests of normality from Kolmogorov–Smirnov, homoscedasticity by Bartlett's test, and then analyzed with a Student's t-test using the Statistic 7.0 software (Statsoft Inc, Tulsa, OK). The weights of undrained and drained seminal vesicles were compared using a paired Student's t-test. Significant differences were considered significant when p<0.05. Values were expressed as the mean and standard error of the mean (±SEM). A minimum of nine animals per group were used for each procedure, and experiments were repeated at least three times.
Results
Thyroid function tests
At the end of experimental period, hearts were weighed and blood was analyzed for serum TSH and T3 concentrations. Compared to the euthyroid group, heart weights and serum T3 concentrations were decreased in the hypothyroid group and the serum TSH concentrations were increased (t-test, p<0.05 and 0.001, Table 2).
Values are mean±SEM; n=20 animals per group. T3 and TSH serum concentrations were measured by radioactive iodine immunoassay. Hypothyroidism was confirmed through the decreased heart weight and T3 serum concentrations and increase in the TSH serum concentrations.
a/b differ p<0.05; c/d differ p<0.001 (Student's t-test).
HW, heart weight; BW, body weight; DHW, dehydrated heart weight; T3, triiodothyronine; TSH, thyrotropin.
Lh mRNA expression in the pituitary of the adult male rats
There was an increase in the expression of Lh mRNA in the hypothyroid group compared to the euthyroid group (1.8-fold, t-test, p<0.05, Fig. 1).
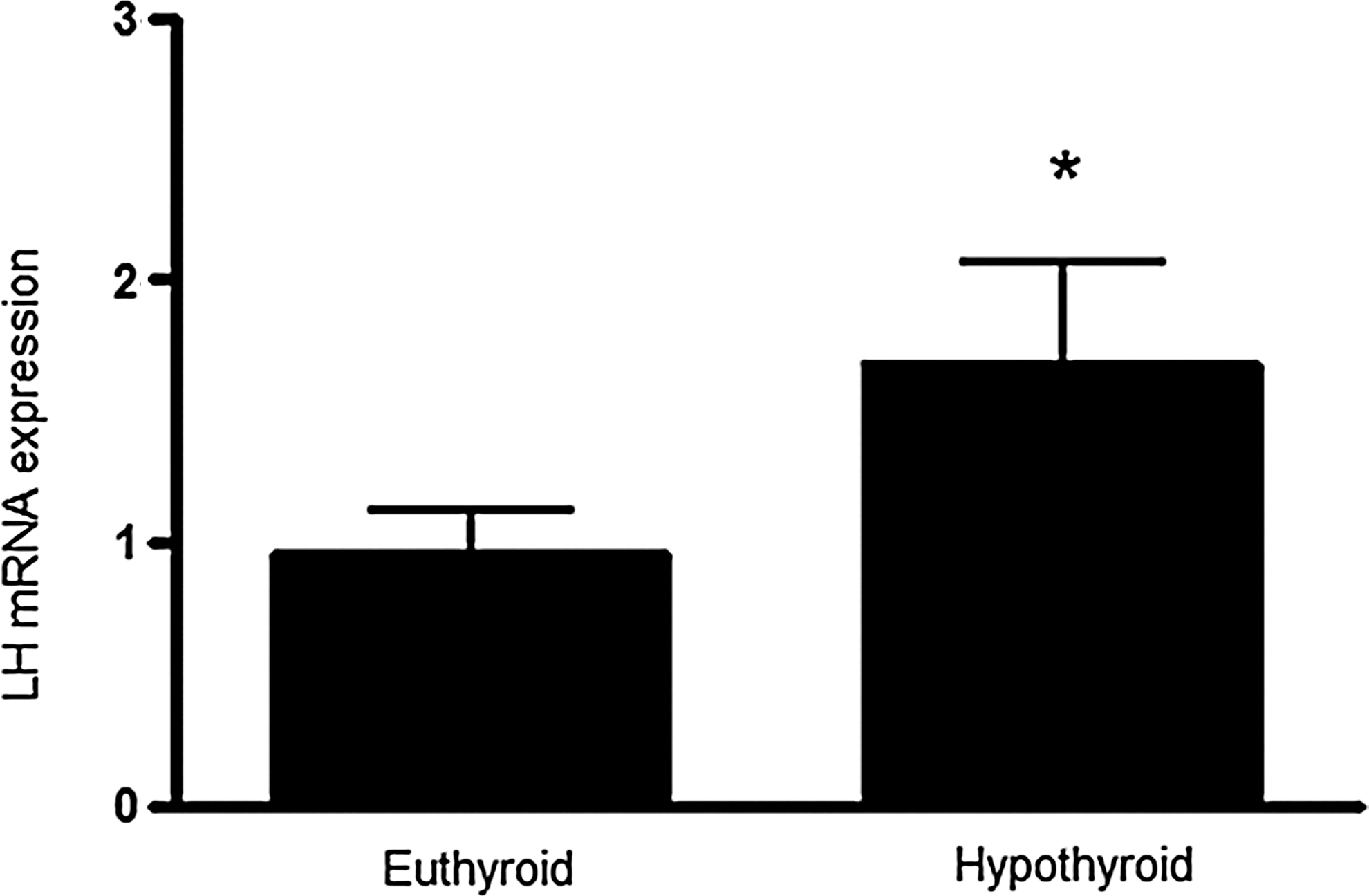
Pituitary Lh mRNA expression relative to Rpl19, a housekeeping gene, evaluated by real-time polymerase chain reaction in euthyroid and hypothyroid male Wistar rats. The Lh mRNA expression increased in hypothyroidism. Values are means±SEM (n=10 animals per group). p-Value indicates the difference between groups using a Student's t-test (*p<0.05). LH, luteinizing hormone; Rpl19, ribosomal protein L19; SEM, standard error of the mean.
Poly(A) tail length of Lh mRNA in the pituitary of the adult male rats
Lh mRNA poly(A) tail length in the pituitaries of the hypothyroid group was significantly shorter than in pituitaries of the control group (t-test, p<0.05, Fig. 2).
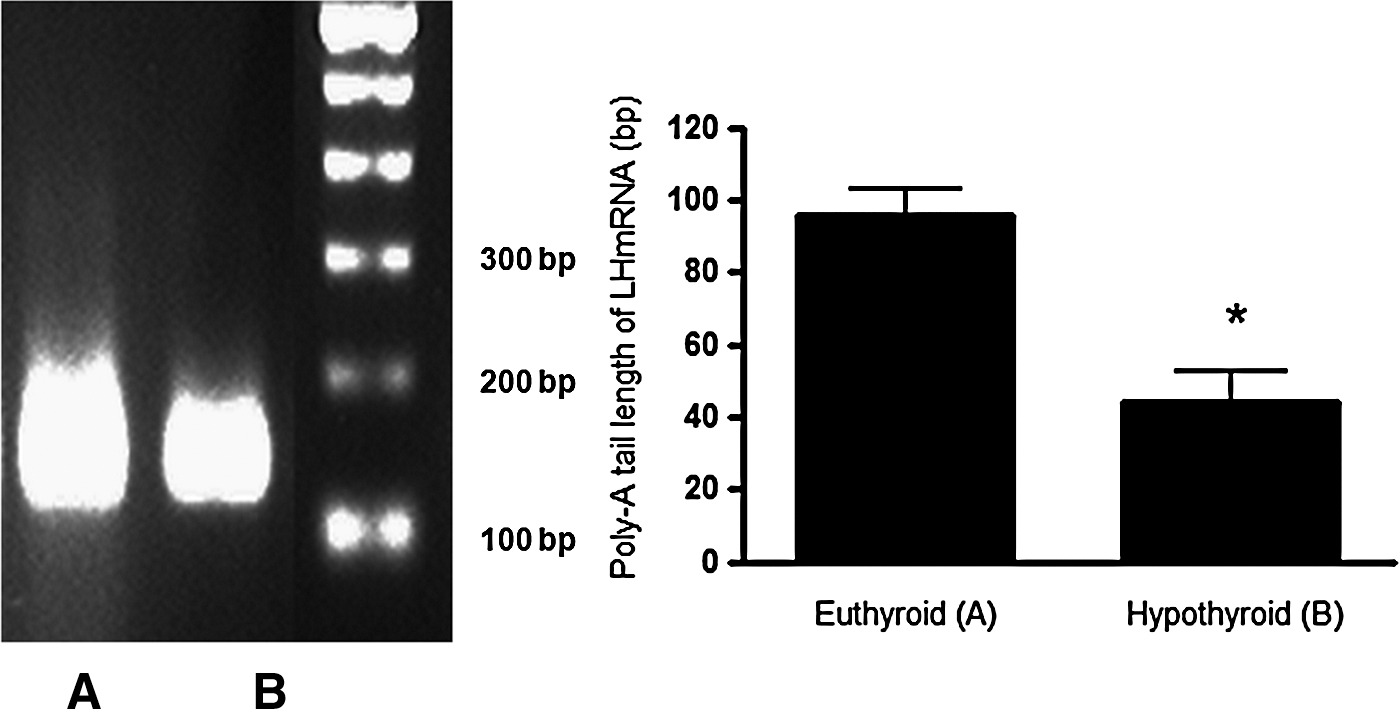
Analysis of poly(A) tail length of Lh mRNA evaluated by rapid amplification of cDNA ends for poly(A) test (RACE-PAT) method in pituitaries of euthyroid and hypothyroid male Wistar rats. Hypothyroidism caused a significant reduction in the Lh mRNA poly(A) tail length. The smearing pattern of PCR products in ethidium bromide-stained 2.5% agarose gel obtained by RACE-PAT is shown at the left. The top of the smear indicated the longest amplified fragment, which represents the poly(A) tail size plus 128 bases upstream, according to the primer used. Values are expressed as mean±SEM (n=10 animals per group). p-Value indicates the difference between groups using a Student's t-test (*p<0.05). Lh, luteinizing hormone.
Hypothyroidism decreases the binding of Lh mRNA to the translational machinery
To determine if the increases in the mRNA expression and the reduction in the poly(A) tail length in the hypothyroid group were associated with changes in parameters of translation, we evaluated the amount of Lh mRNA bound to polysomes in polysomal profiles. Although the polysomal profile was not changed in the hypothyroid group (Fig. 3A), the amount of Lh transcripts anchored to polysomes were decreased, indicating a reduction in recruitment of Lh mRNAs to the translational machinery in the hypothyroid group (t-test, p<0.01, Fig. 3B).
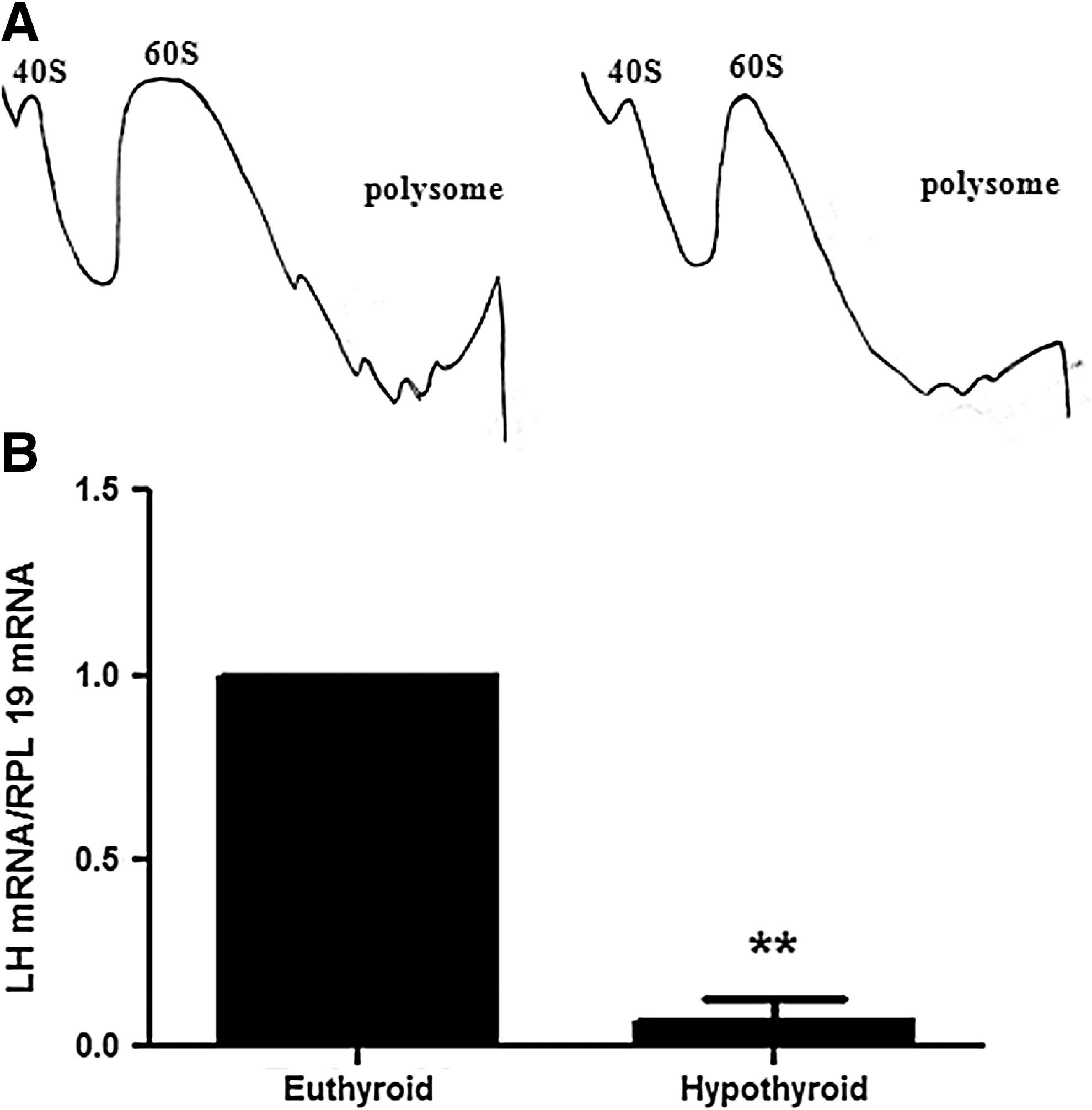
Quantification of Lh mRNA bound to the polysome in pituitaries of euthyroid and hypothyroid male Wistar rats. Hypothyroidism reduced the content of LH mRNA bound to the polysome. The ribosomal subunits are indicated on the polysomal profile in the 40S and 60S peaks, followed by peak of monosomal 80S and polysome
Serum LH concentrations and pituitary LH content in hypothyroid rats
As noted, the amount of mRNA associated with the polysomes was decreased, although the total amount of the transcripts were increased. We determined if the changes in Lh mRNA in the hypothyroid group were associated with changes in serum and pituitary LH content using the methods described. Serum LH concentrations were increased in the hypothyroid group compared to the euthyroid control group (5-fold, t-test, p<0.05, Fig. 4B), whereas pituitary LH content was reduced (0.6-fold, t-test, p<0.01, Fig. 4A).
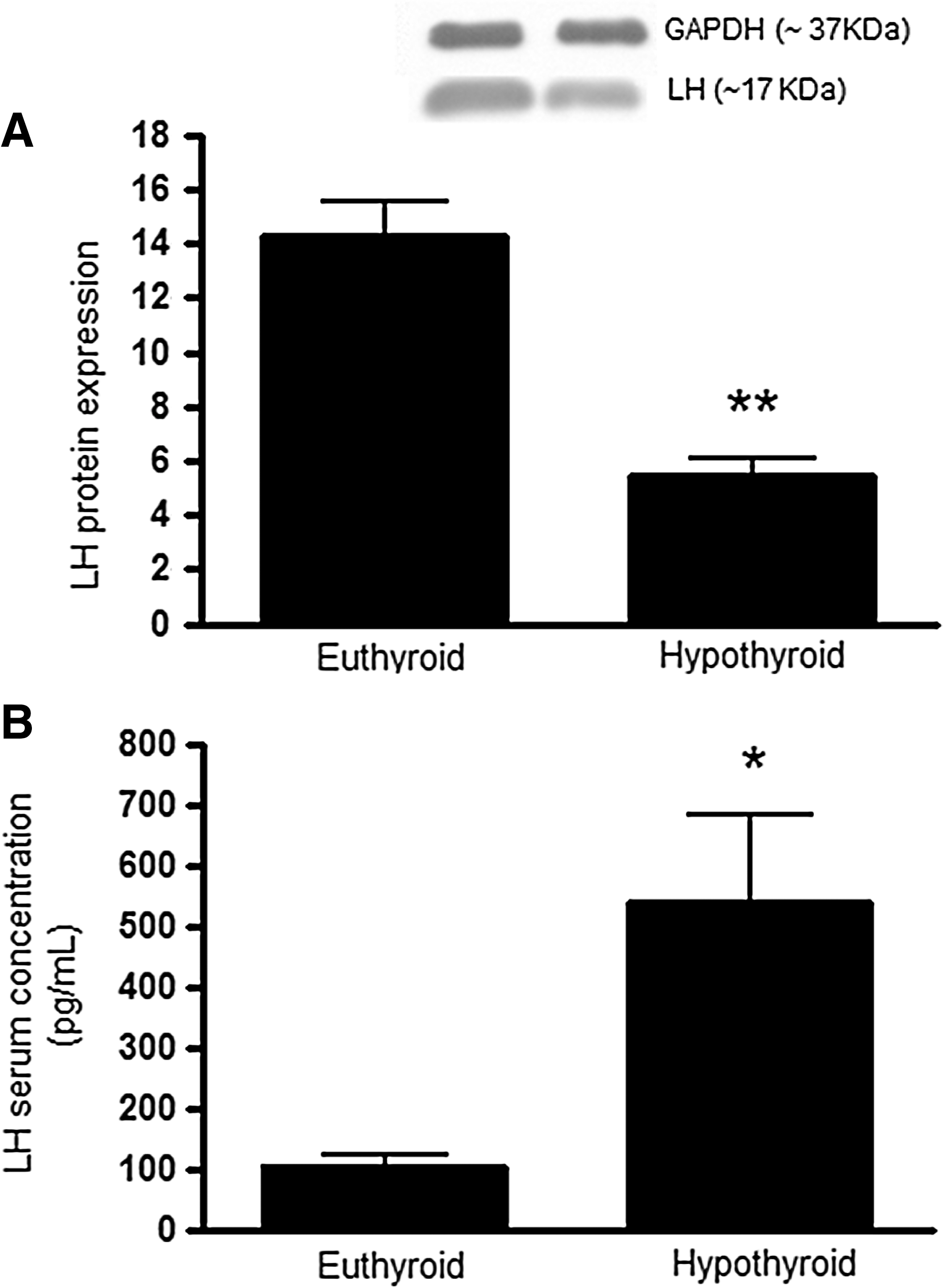
Pituitary LH protein content relative to Gapdh, a housekeeping gene, evaluated by Western blotting
Serum testosterone concentrations and the weight of androgen-responsive organs in hypothyroid rats
Serum testosterone concentrations were substantially reduced in hypothyroid rats compared to control euthyroid rats (10-fold, t-test, p<0.01, Fig. 5). Testes weight was similar in hypothyroid and euthyroid rats (Fig. 6A). The mean weight of the epidydimis in the hypothroid group was reduced compared to the euthyroid group, but this difference was not significant (t-test, p=0.1, Fig. 6B). However, ventral prostate weight was significantly decreased in the hypothyroid group (70%; t-test, p<0.001, Fig. 6C), and the seminal vesicles were also much smaller in size and contained less seminal fluid (paired t-test, p<0.05, Fig. 6D) in the hypothyroid rats.
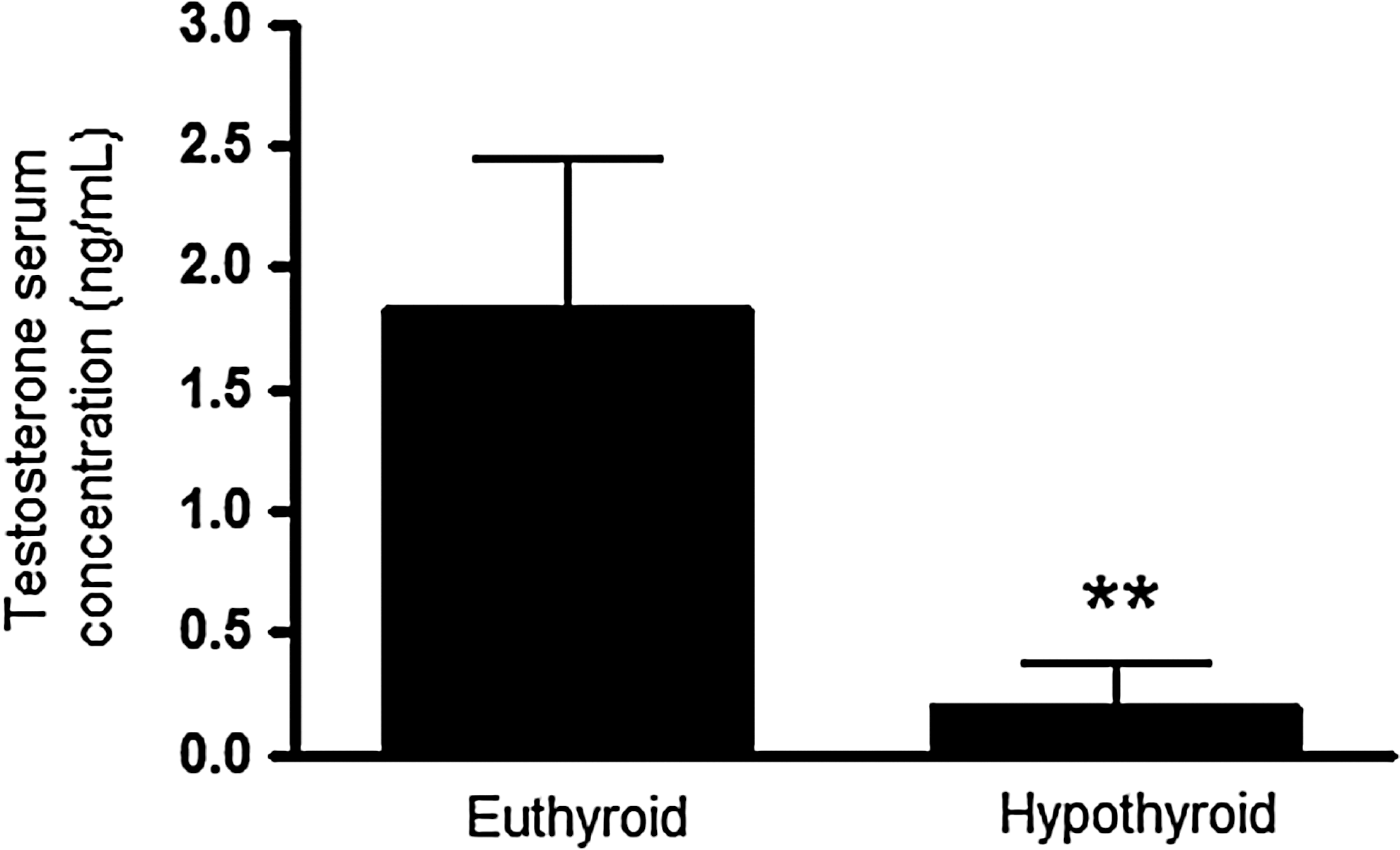
Testosterone serum concentrations evaluated by radioimmunoassay in euthyroid and hypothyroid male Wistar rats. Hypothyroidism caused an intense reduction in the serum testosterone concentrations. Values are expressed as mean±SEM (n=10 animals per group). p-Value indicates the difference between groups using a Student's t-test (**p<0.01).
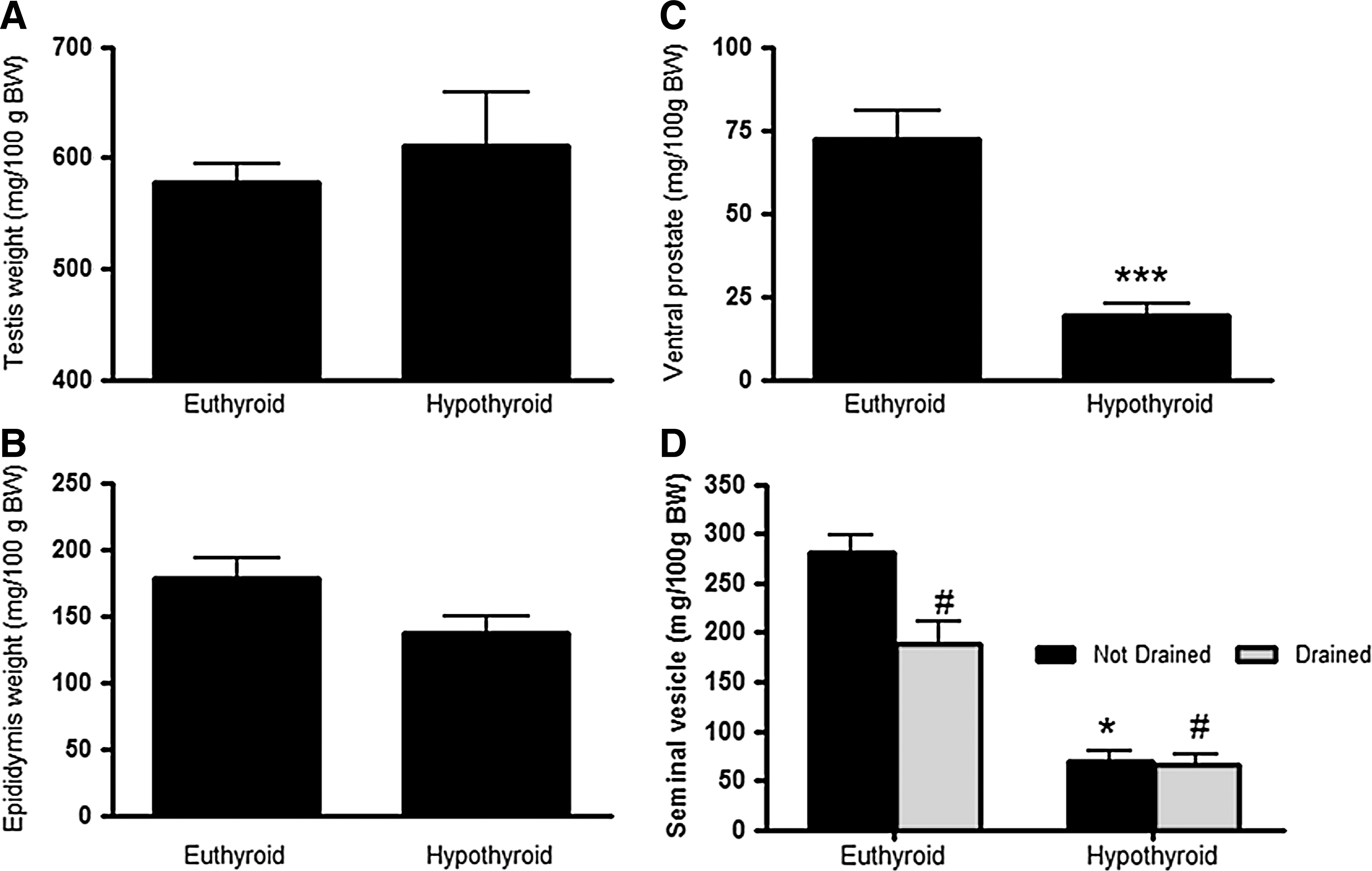
Reproductive organ weight in euthyroid and hypothyroid male Wistar rats. The weights of the testis
Testicular LH receptor in the testes
The mRNA expression of Lh receptor was increased in the hypothyroid group of rats (1.5-fold, t-test, p<0.05, Fig. 7A). However, both mature and imature forms of the LHR were reduced by approximately 25% in hypothyroid rats (t-test, p<0.05, Fig. 7B and C).
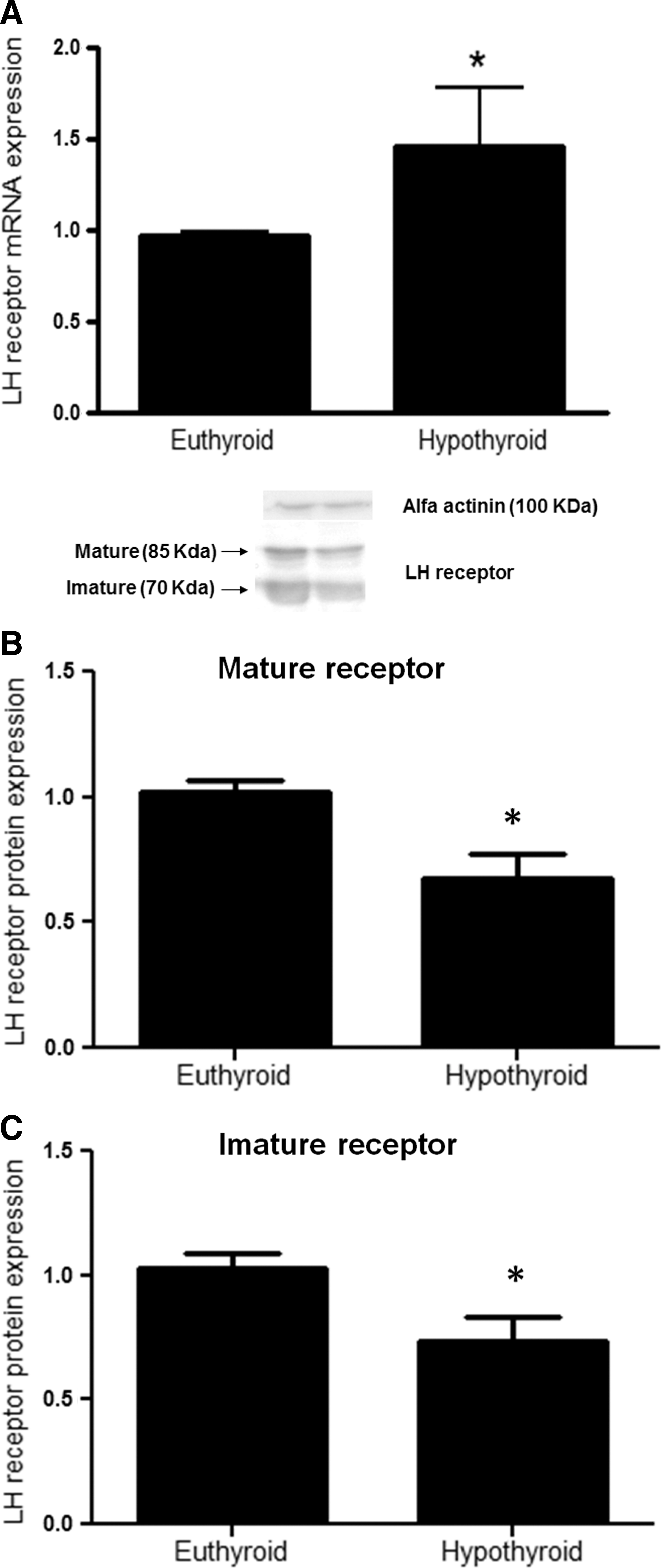
Testicular Lh receptor mRNA expression relative to Rpl19, a housekeeping gene, evaluated by real-time polymerase chain reaction
Morphometry of seminiferous epithelium
Epithelium height (t-test, p<0.05) was reduced, and the tubular lumen was enlarged (t-test, p<0.01) in the hypothyroid group compared to the euthyroid group. However, no alterations were observed in the total tubular diameter (Table 3 and Fig. 8).
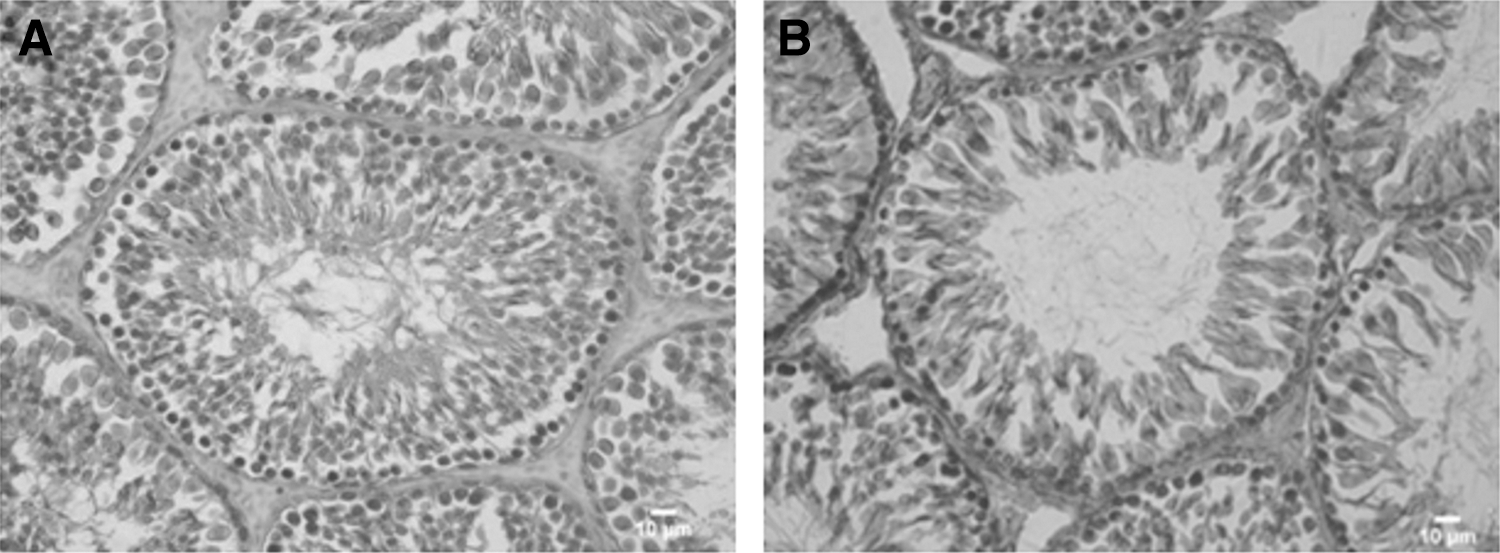
Histological sections of the testes in euthyroid
Values are mean±SEM; n=20 animals per group. The hypothyroid group showed a reduction on epithelial height, with no changes in tubular diameter, which could be related with a harmful effect on the spermatogenesis process.
a/b differ p<0.05; c/d differ p<0.01 (Student's t-test).
Discussion
Previous studies have reported that hypothyroidism induces hypogonadotropic hypogonadism, a hypergonadotropic state, or does not affect the basal levels of LH in rats (1,2). In the present study, we re-evaluated this issue attempting to identify the hypothyroidism-induced changes in post-transcriptional aspects of LH production in rats and its putative effects on testis function.
We show that Lh mRNA expression was increased in pituitaries of hypothyroid rats. Concomitant with this effect, a reduction in the length of Lh transcript poly(A) tails was observed, which could increase its vulnerability to nucleases and reduce its stability. This would impair the Lh mRNA translation, since this processes is directly related to the poly(A) tail length (17,18). Moreover, the polysome profile analysis revealed a lower association of Lh mRNA to the polysomes, which indicates that its translation rate is reduced. This post-transcriptional modification was also observed with the GH transcripts of hypothyroid rats (6), and the opposite was observed with the poly(A) tail length of TSH transcripts, which are negatively regulated by TH (7).
We have also detected a reduction of the pituitary content of LH, which could be due to the decreased translational rate, just reported. The former finding was also previously described (4,19). In contrast, we have detected a high amount of LH in serum, which could be due to an impaired LH renal clearance, since hypothyroidism can alter clearance (1) and renal drug excretion (20).
Despite high levels of circulating LH, which is supposed to bind its receptor in the testes leading to testosterone production, there was a marked decrease of total testosterone serum concentration in the hypothyroid rats, which may be mediated by a reduction in the amount of testicular LHRs, similar to that observed in primary hypogonadism.
In fact, we have observed a significant reduction in testicular LHR content, even though Lh mRNA receptor expression was shown to be increased. These findings indicate that in parallel to the post-transcriptional alterations in LH synthesis, hypothyroidism also interferes with the regulation of Lhr gene expression. Indeed, in vitro studies have shown that there are functional TREs in the Lhr gene (21). It is worth mentioning that the exposure of testicular cells to a high concentration of LH reduces both Lhr mRNA and LHR protein (22).
Nevertheless, we cannot ignore that the lower testosterone concentration reduces the negative feedback loop regulating LH and GnRH, leading to an increased production and secretion of LH. Thus, increased Lh gene transcription could compensate the lower association of Lh transcripts to polysomes.
The low serum testosterone concentrations seemed to affect the accessory sex organs, as shown by the low weight of the seminal vesicle and ventral prostate, which could be compromising reproductive function, since problems in sperm quality have been described in hypothyroid humans (1,23). The relative testis weight was not altered by lack of TH. Although it has been observed that hypothyroidism during the prepubertal period can lead to hypergonadism and cellular immaturity in testes of rats (24,25), in the present study the animals were already in adulthood and testicular development had already been completed.
The reduced epithelial height of the seminiferous tubules could also be ascribed to the hormonal imbalance. Decreases in testosterone, which is responsible for initial proliferation of spermatogonia in seminiferous epithelium by binding to the androgen receptor of Sertoli cells, and also the reduction in follicle-stimulating hormone serum concentrations (50% less than control, data not shown), could impair the spermatogenesis process, as described (26). Hence, a reduction in testosterone may cause changes in the architecture of the testes (16). Impairment in spermatogenesis due to hypothyroidism was also observed in human patients, in which a decrease in testicular germ cells and number of live sperm cells (27) and an increase in abnormal sperm morphology in semen were detected, effects that were restored by TH treatment (23).
Interestingly, some TH actions on Leydig and Sertoli cell growth, differentiation, and steroidogenesis are supposed to occur only during prepubertal phase (28). However, some dysfunctions of Leydig cells during hypothyroidism were reversible by TH supplementation in adulthood (29,30), which is highly suggestive that TH acts on adult testes. Despite this evidence, TH action on gonads is still controversial because a small amount of TRs are present in the testes of adult animals, which has classically been considered unresponsive to TH (31,32).
Conclusion
The results presented herein show that the hypothyroidism induced by thyroidectomy in rats leads to a decrease in serum testosterone concentrations, which reduces the negative feedback on GnRH and LH release and results in LH stimulation. The high serum LH is not able to stimulate the regular synthesis of testosterone because the content of LHRs is decreased, but we cannot discard the possibility that alterations in LH bioactivity could occur. As a result of low testosterone, the weight of the androgen-responsive tissues is reduced and the testicular morphology is altered. Additional experiments are necessary to evaluate the role of TH on the secretion and bioactivity of LH.
Footnotes
Acknowledgments
This work was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo—FAPESP, (2008/50977-2, 2009/17822-8) and Conselho Nacional de Desenvolvimento Científico e Tecnológico—CNPq (305659/2009-0), Brazil. The authors thank Leonice Lourenço Poyares (Department of Physiology and Biophysics, Institute of Biomedical Sciences, University of Sao Paulo) for his technical assistance.
Author Disclosure Statement
No competing financial interests exist.