
Abstract
Background:
Thyroglobulin (Tg) is a macromolecular precursor in thyroid hormone synthesis to which iodine is stably bound. Tg, which is stored in the follicular space, is also a potent negative feedback regulator of follicular function, and this is achieved by suppressing mRNA levels of thyroid-specific genes such as the sodium/iodide symporter (Slc5a5), Tg, and thyroid peroxidase. Dual oxidase 1 (DUOX1) and DUOX2, originally identified in the thyroid, are nicotinamide adenine dinucleotide phosphate (NADPH) oxidases that are necessary to produce the H2O2 required for thyroid hormone biosynthesis. Since follicular Tg regulates the expression of genes that are essential for thyroid hormone synthesis, we hypothesized that Tg might also regulate DUOX expression and H2O2 production.
Methods:
Rat thyroid FRTL-5 cells were treated with Tg, and the mRNA expression of Duox1 and Duox2 and their corresponding maturation factors Duoxa1 and Duoxa2 were evaluated by DNA microarray and real-time PCR. Duox2 promoter activity was examined by luciferase reporter gene assay. Protein levels of DUOX2 were also examined by Western blot analysis. Intracellular H2O2 generation was quantified by a fluorescent dye, 5-(and-6)-chloromethyl-2′,7′-dichlorodihydrofluorescein diacetate, and acetyl ester (CM-H2DCFDA).
Results:
mRNA levels of Duox2 and its activation factor Duoxa2 (but not Duox1 or Duoxa1) were significantly suppressed by Tg in a dose-dependent manner and a time-dependent fashion in rat thyroid FRTL-5 cells. DUOX2 promoter activity was significantly suppressed by Tg in a dose-dependent manner. Protein levels of DUOX2 and H2O2 generation in cells were also reduced by Tg treatment.
Conclusions:
We show that physiological concentrations of Tg suppressed the expression and function of DUOX2 in thyroid cells. These results suggest that Tg is a strong suppressor of the expression and the activity of DUOX2/DUOXA2, thereby regulating iodide organification and hormone synthesis in the thyroid. The evidence supports a reported model in which accumulated Tg in thyroid follicles plays important roles in autoregulating the function of individual follicles, which produces the basis of follicular heterogeneity.
Introduction
The thyroid gland is a unique organ that utilizes dietary iodine to synthesize thyroid hormones, a process requiring H2O2, which is the final electron acceptor for the biosynthesis of the thyroid hormone catalyzed by TPO at the apical surface of thyrocytes. H2O2 is produced by two isoform enzymes, DUOX1 and DUOX2, which were initially identified in the thyroid as nicotinamide adenine dinucleotide phosphate (NADPH) oxidases (NOX) that produce H2O2, either by direct generation or through the production of O2 − as a precursor (1 –4). The activities of NOX/DUOX are regulated at the level of enzyme assembly. In resting cells, they predominantly present either as an unassembled form on the plasma membrane or in the cytosol (5). Nox family members are transmembrane proteins that transport electrons across biological membranes to reduce oxygen to superoxide. Seven molecular species have been detected in the NOX family: NOX1–NOX5, DUOX1, and DUOX2. In the case of NOX2 stimulation, transmembrane protein p22phox and cytosolic components p47phox, p67phox, p40phox, and rac2 translocate to the plasma membrane and associate there with membrane-bound flavocytochrome b to constitute an intact and functioning enzyme (6). NOX1 requires NOXA1 and NOXO1, while NOX3 requires p22phox. NOX4 does not require a cytosolic subunit, and NOX5 activation is mediated by an increase in the cytoplasmic Ca2+ concentration (7). NOX isoforms (NOX1, NOX2, NOX3, and NOX5) are not expressed in the thyroid tissue, whereas DUOX1 and DUOX2 are primarily expressed in the thyroid tissue and in respiratory and gastrointestinal epithelial cells. Recently, NOX4 was reported to be present in normal and cancer thyroid tissue, but the function of the molecule is not clear (8).
DUOXA1 and DUOXA2 are required as maturation factors to localize DUOX1 and DUOX2, respectively, to the cell membrane (2). Both DUOX and TPO colocalize on the apical membrane of the thyroid follicular epithelium (9). The H2O2 produced by DUOXs is used by TPO to oxidize iodide, to iodinate tyrosyl residues of Tg to form monoiodotyrosine and diiodotyrosine; then, they subsequently couple to the iodotyrosines to form thyroid hormones (T3 and T4) (10). Several other molecules are also involved in this process. Membrane proteins such as EF-hand binding protein 1 (EFP1), a member of the thioredoxin-related protein family, and p22phox were shown to form a multiprotein complex with DUOX and TPO (11). Membrane raft protein caveolin-1 seems to play an important role in the sorting of DUOX and TPO (12), although the role of these components in H2O2 generation is not fully understood. Generated thyroid hormones are transported by thyroid hormone transporters, such as monocarboxylate transporter 8 (MCT8)/Slc16a2 and MCT10/Slc16a19, which catalyze proton-linked transport across the plasma membrane (13).
The major role played by DUOXs in thyroid hormone synthesis has been proved in patients suffering from congenital hypothyroidism (CH) that is caused by mutations of either DUOX2 or DUOXA2 (14 –19); while the roles of DUOX1 and DUOXA1 in the thyroid are still elusive. The DUOX proteins have two Ca2+ binding EF-hand motifs that are considered as accounting for their Ca2+-dependent activity (20,21). While the induction of Duox expression by TSH seems to be less crucial than other thyroid genes such as Slc5a5, Tg, and Tpo (21 –23), the molecular mechanisms that control Duox expression are not known.
Tg is a major product of the thyroid gland that provides sites for iodide organification. The stored Tg in the thyroid follicle is a potent negative feedback regulator of follicular function (24 –31). We have shown that physiological concentrations of follicular Tg suppressed not only thyroid-specific genes such as Slc5a5, Tpo, and Tg but also iodide uptake in the thyroid (28,30,32). In addition to suppressing gene expression, low concentrations of Tg also induce expression of the Pendred syndrome gene (Slc26a4) that mediates iodide transportation to the follicular lumen, which enhances hormone synthesis until enough Tg is accumulated, and the process is terminated by Tg-mediated suppression (25,33). Although Tg has strong effects, the underlying biochemical mechanisms that explain the action of Tg are still largely unknown. Thus, the possible presence of bioactive sites within the Tg molecule (660 kDa), cellular recognition systems for Tg (or its degradation fragments), and the signaling cascades that exert such unique and potent effects remain to be defined. However, the effects were shown to be specific to the Tg molecule and are not mimicked by other factors (30,34 –36). Although Tg prepared from iodide-deficient goiter and congenital goiter suppressed thyroid-specific gene expression more effectively than did Tg prepared from normal tissue (27,30), the effects of immature Tg under physiological conditions are not clear, especially because Tg moieties bind to several proteins in order to undergo vectorial transport to the follicular lumen (37 –40). The range of Tg concentrations required for these effects was 1–10 mg/mL, which is within the normal follicular Tg concentration range of 0.1–250 mg/mL (41 –43), reflecting follicular heterogeneity. Although it was previously shown that follicular Tg plays an important role in the regulated expression of several thyroid-specific genes, the effect of Tg on DUOXs is not known. In this study, we investigated the possible role of Tg on the expression and function of DUOXs using rat FRTL-5 thyroid cells.
Materials and Methods
Cell culture and treatment
The effect of Tg on DUOX expression and function was analyzed using rat FRTL-5 thyroid cells. The cells were grown either in Coon's modified Ham's F-12 medium supplemented with 5% bovine serum (Invitrogen) and a six-hormone mixture (1 mU TSH/mL, 10 μg insulin/mL, 10 ng somatostatin/mL, 0.36 ng hydrocortisone/mL, 5 μg transferrin/mL, and 2 ng glycyl-L-histidyl-L-lysine acetate/mL) or with 0.2% bovine serum without TSH and insulin mixture as previously described (44,45). Bovine Tg (Sigma) was used at a final concentration of 1–30 mg/mL, which was previously confirmed as having essentially the same effect as highly purified 19S Tg (30,34,35).
DNA microarray analysis
FRTL-5 cells grown in the presence or absence of TSH; insulin or serum were treated with the medium containing 10 mg/mL of Tg for 6 or 24 hours. Total RNA was purified using the RNeasy Mini Kit (Qiagen). For the preparation of labeled double-strand (ds) DNA, 10 μg of total RNA was reverse transcribed using SuperScript II (Invitrogen). The generated cDNA was incubated with 4 μg of RNase A (Promega) at 37°C for 10 minutes, phenol-chloroform extracted, and precipitated with ethanol. Cy3 labeling reactions were performed with NimbleGen One-color Labeling Kit (Roche Diagnostics). About 1 μg ds-cDNA was incubated for 10 minutes at 98°C with one OD unit of Cy3-9mer primer. The addition of 8 mmol of dNTPs and 100 U of Klenow fragment was followed by incubation at 37°C for 2 hours. The reaction was stopped by adding 0.1 volume of 0.5 M EDTA, and labeled cDNA was precipitated with isopropanol. Cy3-labeled samples were resuspended in NimbleGen Hybridization Buffer (Roche Diagnostics), denatured at 95°C for 5 minutes, and hybridized to arrays in a NimbleGen Hybridization System (Roche Diagnostics) for 18 hours at 42°C. The arrays were washed using the NimbleGen Wash Buffer Kit, dried by centrifugation, and scanned at 5 μm resolution using the GenePix 4000B scanner (Molecular Devices).
Total RNA isolation and real-time PCR
Total RNA was purified using RNeasy Plus Mini Kit (Qiagen), and cDNA was synthesized using the High-Capacity cDNA Reverse Transcription Kits (Applied Biosystems) as described (44,45). Quantitative real-time PCR was performed using the ABI Prism 7700 Sequence Detection System (Applied Biosystems). A total of 20 ng of cDNA mixed with 20 μL of FastStart Universal SYBR Green Master (Roche Diagnostics) was amplified by incubation for 10 minutes at 95°C, followed by 40 cycles consisting of 15 seconds at 95°C and 1 minute at 60°C. The mRNA levels were normalized against β-actin using the ΔΔCt method as described (44,45). The sequences of PCR primers were as follows: Duox1 forward, 5′-CCGTTTCTTGGGAGGTCCAGCGAT-3′; Duox1 reverse, 5′-CGTGGTAGCCGAAGAAGACACCCAG-3′; Duoxa1 forward, 5′-GCTGTTCTGGCTGCTACGGGTG-3′; Duoxa1 reverse, 5′-ACTGGGGTCCCTGTGAGGGTGA-3′; Duox2 forward, 5′-CAGCGCTACGACGGCTGGTTTA-3′; Duox2 reverse, 5′-CCCAAGCACTGTGCGGTTGT-3′; Duoxa2 forward, 5′-TCAGCGTACCGCTGCTCATCGT-3′; and Duoxa2 reverse, 5′-ACCAACCAGAACCAGCGCGAGT-3′.
Plasmid construction, transient transfection, and reporter gene assay
A luciferase reporter gene construct driven by 5′-932 to +135 (relative to the transcription initiation site) of the human DUOX2 (NM_014080.4) promoter (pSGG_Prom human DUOX2) was purchased from Promega. The promoter region spanning 5′-626 to +135 was PCR-amplified using the following forward and reverse primers containing MluI and BglII restriction sites, respectively, and subcloned into empty pSGG_Prom plasmids: forward, 5′-AGGACGCGTCCCCCACTCTACCTGCACTA-3′, reverse, 5′-CGAAGATCTGCTTCTGTGCTCTACTTCTTG CC-3′. Inserts were sequenced to ensure there was no misincorporation.
FRTL-5 cells (2×104) plated on poly-D-lysinecoated 24-well plates (Greiner Bio One) were transfected with 0.2 μg of the reporter plasmids using FugeneHD (Roche Diagnostics). Cells were then stimulated with 1–30 mg/mL of Tg. The DUOX2 promoter assay was performed with the Bright-Glo Luciferase assay systems (Promega) 48 hours after Tg stimulation. Luciferase activity was measured by FLUOstar galaxy (BMG Labtechnologies), and the data were normalized to corresponding protein concentrations, which were determined using a Bio-Rad Protein Assay Kit (Bio-Rad) (44,45).
Western blotting analysis
To extract protein from FRTL-5 cells, cultures were lysed in a buffer containing 50 mM HEPES, 150 mM NaCl, 5 mM EDTA, 0.1% NP40, and 20% glycerol, followed by the addition of protease inhibitor cocktail tablets (Complete Mini; Roche Diagnostics). Cellular protein extracts were mixed with a 4× lithium dodecyl sulfate sample buffer and a 10× reducing agent (Invitrogen) and incubated for 10 minutes at 70°C before electrophoresis. The proteins were separated on NuPage 4%–12% Bis Tris by gel electrophoresis and transferred to i-Blot Gel transfer stacks of nitrocellulose (Invitrogen) (44,45). The membranes were washed with PBS with 0.1% Tween 20, and then incubated with either goat polyclonal anti-β-actin (Santa Cruz Biotechnology; dilution 1:1000) or goat polyclonal anti-DUOX2 antibody (sc-49939) (Santa Cruz; dilution 1:4000) for 12 hours at 4°C. Membranes were incubated with donkey anti-goat IgG-biotin conjugates (Millipore; dilution 1:10,000) for one hour at room temperature, followed by streptavidin horseradish peroxidase (HRP) (GE Healthcare; dilution 1:20,000) for one hour at room temperature. HRP was visualized using the Amersham ECL Plus Western Blotting Detection System (GE Healthcare) and exposed to X-ray film (44,45).
Measurement of H2O2 generation
The generation of H2O2was quantified in FRTL 5 cells by a fluorescent dye 5-(and-6)-chloromethyl-2′,7′-dichlorodihydrofluorescein diacetate, acetyl ester (CM-H2DCFDA; Invitrogen). CM-H2DCFDA is nonfluorescent until the acetate groups are removed by intracellular esterases, and oxidation occurs within the cells. In the presence of H2O2, CM-H2DCFDA is oxidized to highly fluorescent DCF. Since released H2O2is immediately absorbed into the cells, the measurement of intracellular H2O2 reflects the generation of H2O2 by DUOX2/DUOXA2. FRTL-5 cells were treated with Tg, washed once with Dulbecco's phosphate-buffered saline (DPBS), and incubated in DPBS containing CM-H2DCFDA (5 μg/mL) for 30 minutes. After washing cells with DPBS, DCF fluorescence was measured in a microplate reader (FLUOstar galaxy; BMG Labtechnologies) using excitation at 485 nm and emission at 520 nm. Extracellular H2O2 was measured using the Amplex Red Hydrogen Peroxide/Peroxidase Assay Kit (Invitrogen) according to the instructions. Briefly, FRTL-5 cells were washed thrice with Hank's balanced salt solution (HBSS) without Ca2+ and Mg2+ [HBSS(−)], resuspended with HBSS(−) or HBSS containing Ca2+ (1.26 mM) and Mg2+ (0.9 mM) [HBSS(+)]. The cells were transferred to 96-well plates and mixed with Amplex Red reaction mixture. The cells were then incubated at 37°C for 3 hours, and fluorescence intensity was measured using excitation at 544 nm and emission at 590 nm. For both measurements, reagents without cells and cells without reagents served as background fluorescence.
Statistical analysis
All experiments were repeated at least thrice. Student's t-test was used for statistical analyses.
Results
Effect of Tg on the regulation of Duox2 mRNA levels in FRTL-5 cells
TSH and insulin/IGF-1 induce the expression of thyroid-specific genes such as Slc5a5 (NIS), Tg, and Tpo, but the expression of these genes is significantly suppressed by physiological concentrations of the Tg protein (28,30). To further investigate the possible effects of Tg on the gene expression of Duox1 and Duox2, rat thyroid FRTL-5 cells grown in the presence or absence of TSH, insulin, and 5% serum were treated with Tg for 24 hours. Total RNA was isolated from cells and subjected to DNA microarray analysis. Tg up-regulated 946 genes more than twofold, and down-regulated 1390 genes less than half in 24 hours (Fig. 1). We have reported that Tg strongly regulates thyroid-specific genes such as Slc5a5, Tpo, Pax8, and thyroid transcription factor-2 (Foxe1). In addition to that, our present study revealed that Tg strongly suppresses Duox2 and Duoxa2 producing H2O2, which is essential for the organification of iodide. Tg decreased Duox2 and Duoxa2 mRNA levels, while it did not significantly affect Duox1 and Duoxa1 mRNA levels (Fig. 2). The original levels of Duox2 and Duoxa2 were higher in the cells maintained in the presence of TSH, insulin, and serum (Fig. 2C, D, closed bars) than those in their absence (Fig. 2C, D, open bars). However, even after 24 hours in the presence of TSH, insulin, and serum, Tg decreased mRNA levels of Duox2 and Duoxa2 as low as those of the control cells maintained without TSH, insulin, and serum (Fig. 2C, D).
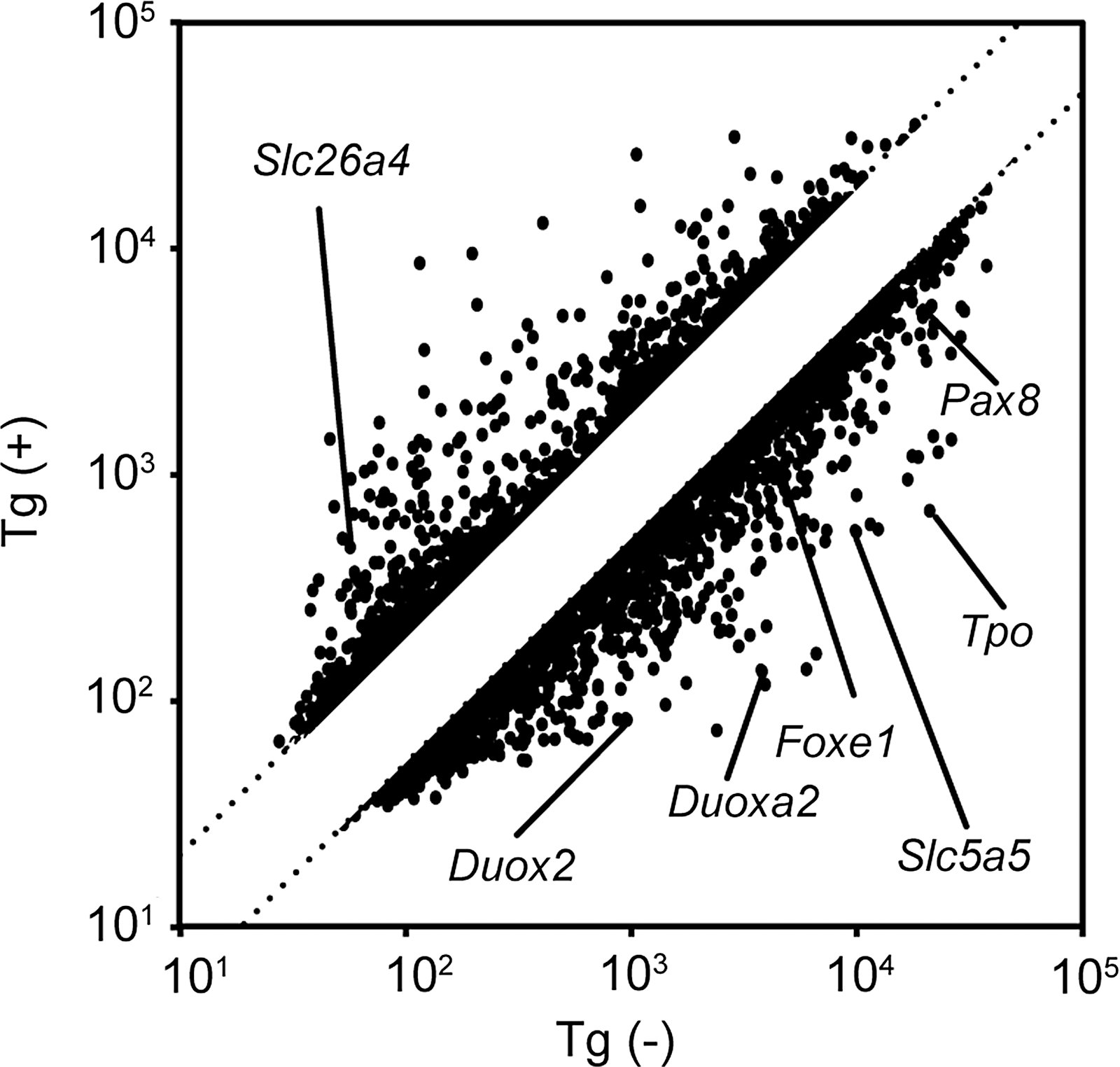
DNA microarray analysis of genes expressed in FRTL-5 thyroid cells after stimulation of Tg. Each • corresponds to the fluorescent intensity of each gene in the microarray of cells treated without Tg (x-axis) or with Tg for 24 hours (y-axis). Twofold and half-fold changes in expression are indicated as parallel lines. The spots of genes regulating cell-signaling, cell-cycle, and thyroid-specific genes are indicated. Tg, thyroglobulin.

Tg suppressed the mRNA expression of Duox2 and Duoxa2, but not Duox1 or Duoxa1. FRTL-5 cells grown in the presence (■) or absence (□) of TSH; insulin and 5% serum were treated with Tg for 6 and 24 hours. Total RNA was isolated at the indicated time after Tg stimulation, and DNA microarray analysis was performed as described in the Materials and Methods section. The normalized signal intensities reflecting the mRNA levels of Duox1
To further evaluate the effects of Tg on Duox mRNA levels, we performed real-time PCR analysis using RNA purified from cells grown in the presence of TSH, insulin, and serum and with different Tg concentrations for 24 hours. Duox2 and Duoxa2 mRNA levels were significantly decreased even when cells were treated with 1 mg/mL of Tg, and the effect of Tg was stronger at higher concentrations (Fig. 3). In fact, 10 mg/mL Tg significantly suppressed Duox2 and Duoxa2 mRNA levels to 4% and 1% of the original levels, respectively. The suppression of mRNA levels of Duox2 and Duoxa2 was evident one hour after stimulation with Tg and maximized after 6 hours. The low levels were sustained even after 72 hours (Fig. 4A, B). The levels of Duox1 and Duoxa1 mRNAs were transiently decreased after 6 to 12 hours and returned to the original levels after 24 hours. The effect was specific to Tg and was not duplicated by the same concentration of bovine serum albumin (BSA), which is in agreement with previous reports (28,30) (Fig. 5A). When Tg was removed from the culture medium, the mRNA expressions of Duox2 and Duoxa2 were restored to the original levels in 24 hours, which is consistent with the previous report (25) (Fig. 5B). These results indicated that Tg suppressed the expression of Duox2 and Duoxa2 mRNAs in both concentration- and time-dependent manners by counteracting the effects of TSH, insulin, and serum to induce these genes (25,28,30).
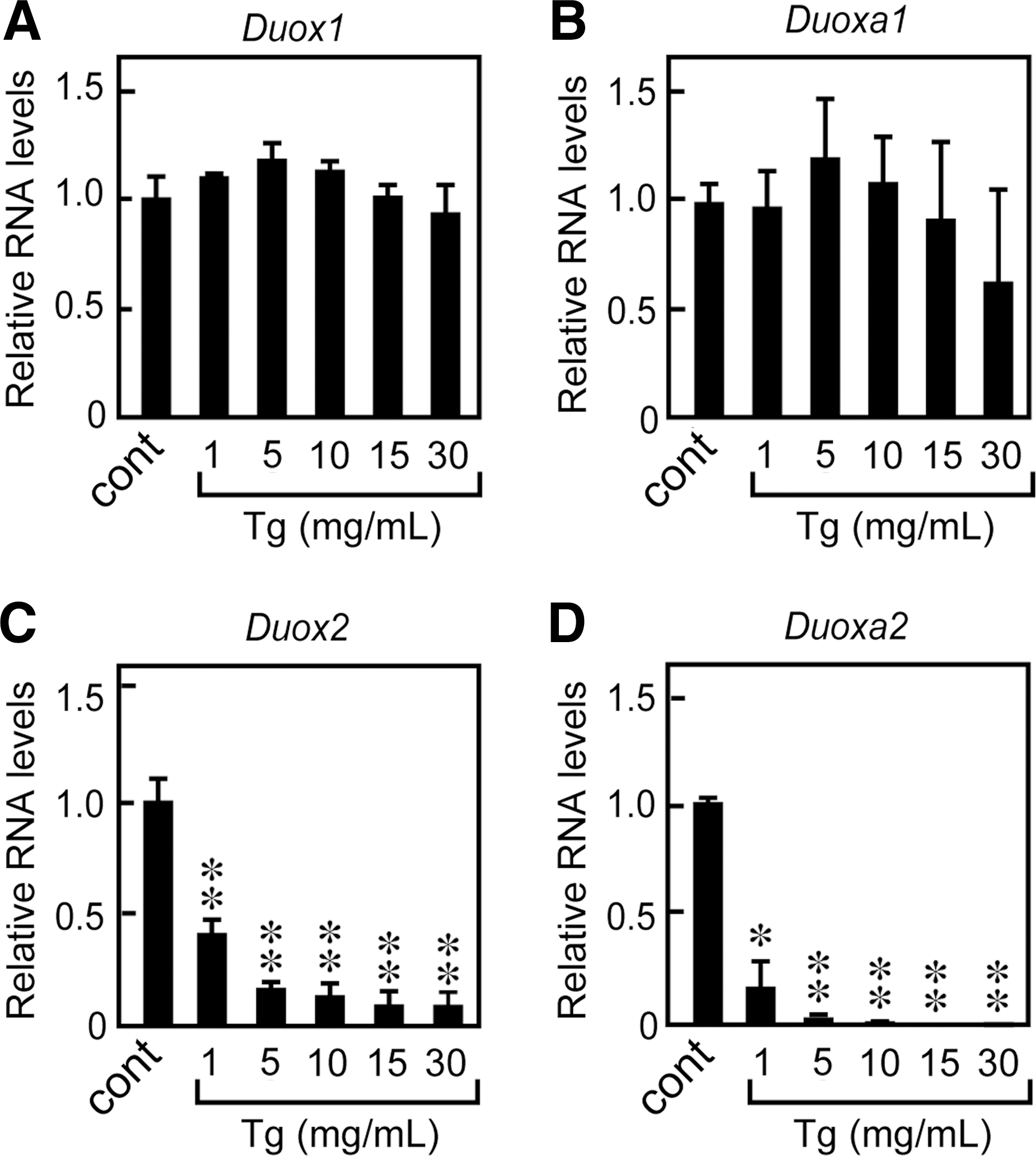
Tg modulated Duox2 and Duoxa2 mRNA levels in a concentration-dependent manner. FRTL-5 cells were treated with 1–30 mg/mL of Tg for 24 hours, and a quantitative real-time PCR analysis of Duox1
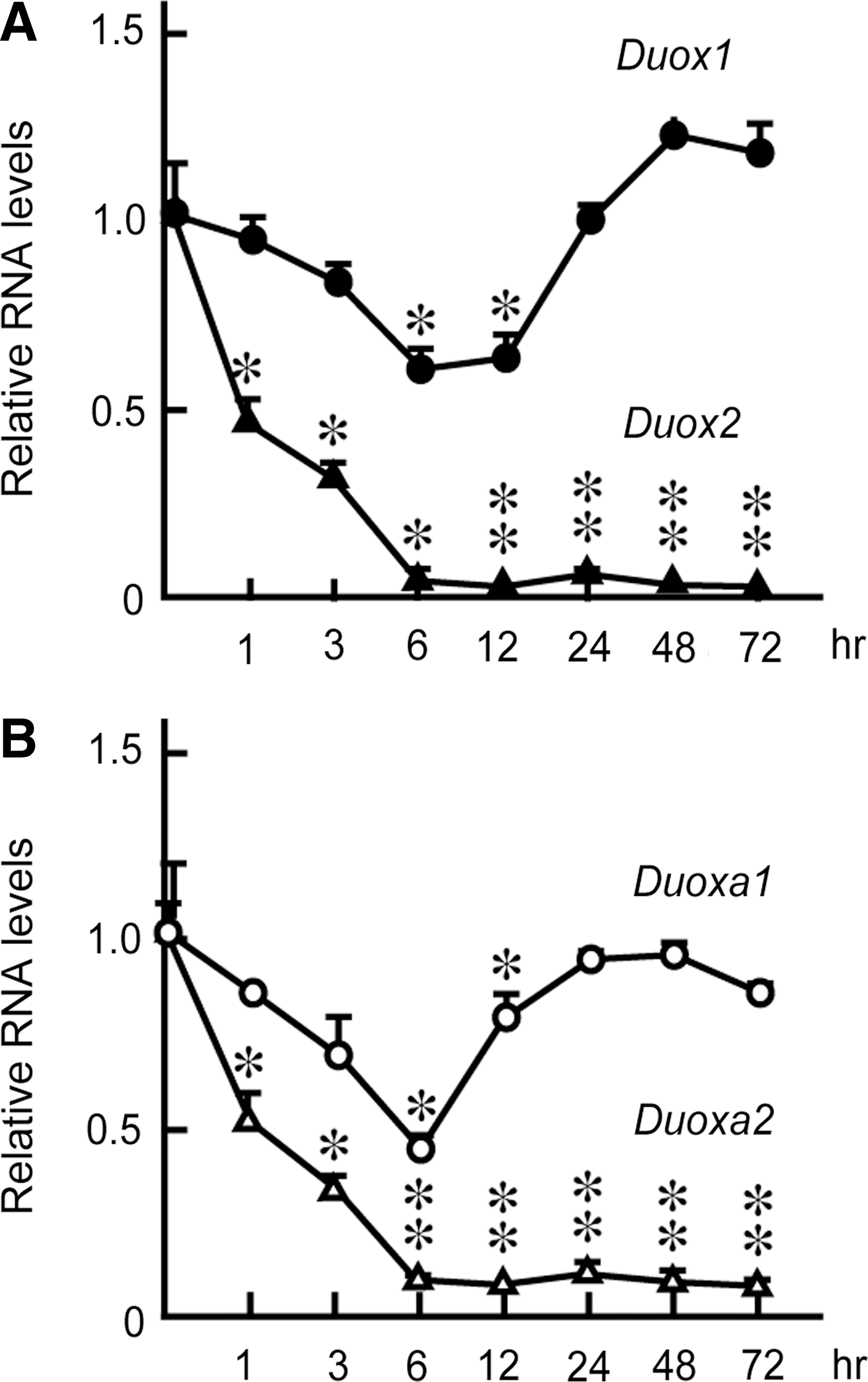
Tg suppressed Duox2
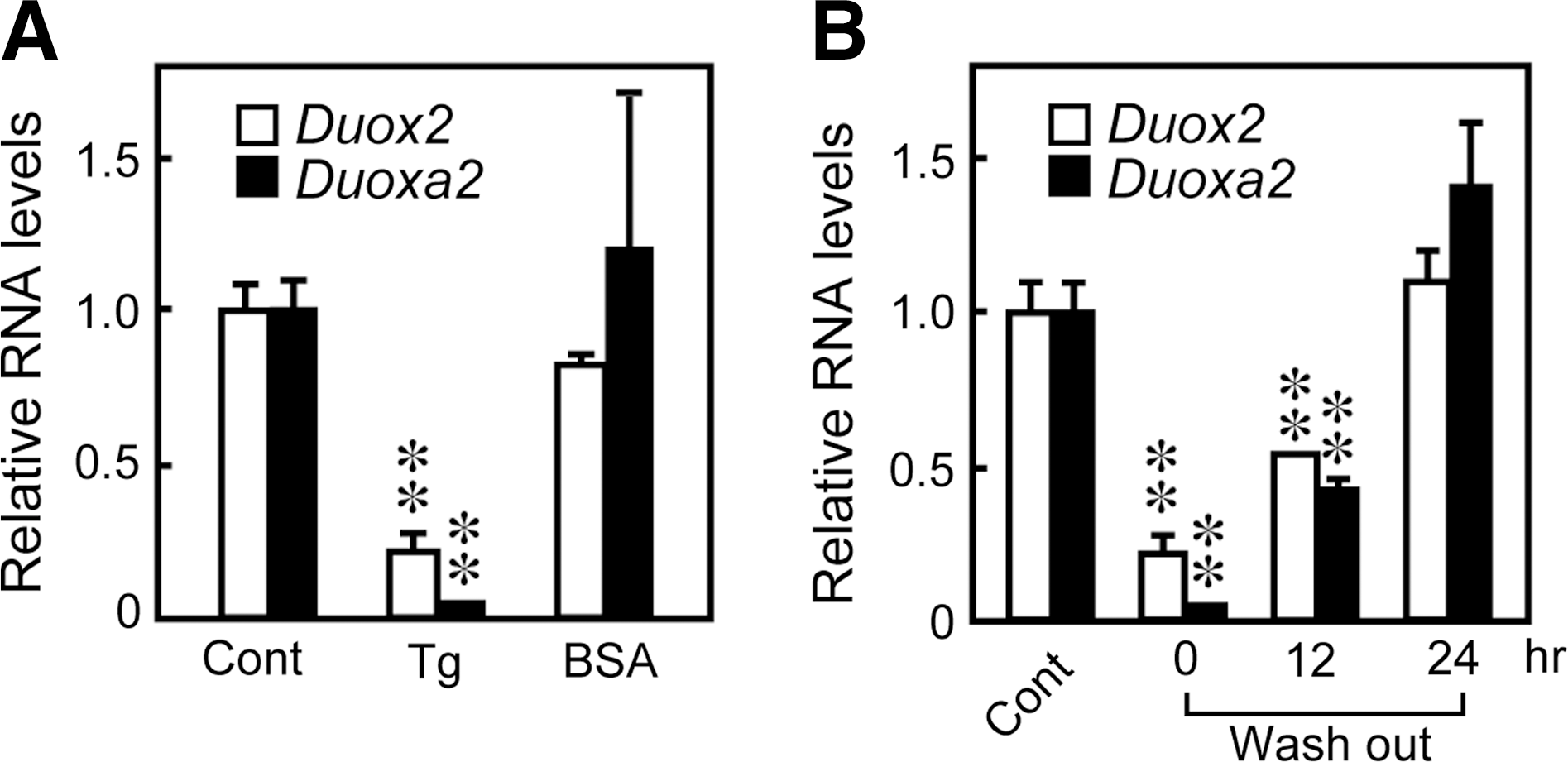
The effect is specific to Tg and was reversible when Tg was removed.
Tg regulated DUOX2 promoter activity
To confirm whether the effect of Tg to decrease mRNA levels of Duox2 and Duoxa2 was mediated by changes in gene transcription, we examined the effect of Tg on DUOX2 promoter activity. FRTL-5 cells were transfected with a full-length (5′-932) or truncated (5′-626) DUOX2 promoter luciferase construct or empty plasmid, then treated with Tg for 48 hours. The full-length DUOX2 promoter (5′-932) was constitutively active in FRTL-5 cells, whereas the activity of the 5′-626 deletion construct was at the same level as those treated with the empty plasmid (Fig. 6). Tg significantly suppressed 5′-932 DUOX2 promoter activity in a dose-dependent manner (Fig. 6). These results indicated that the observed decrease in Duox2 mRNA levels by Tg treatment was mediated by the suppression of DUOX2 promoter activity. The data also suggest that an essential cis-element which is responsible for the expression in thyroid cells and suppression in response to Tg is localized between 5′-932 and 5′-626 bp of the 5′-flanking region of the DUOX2 promoter.
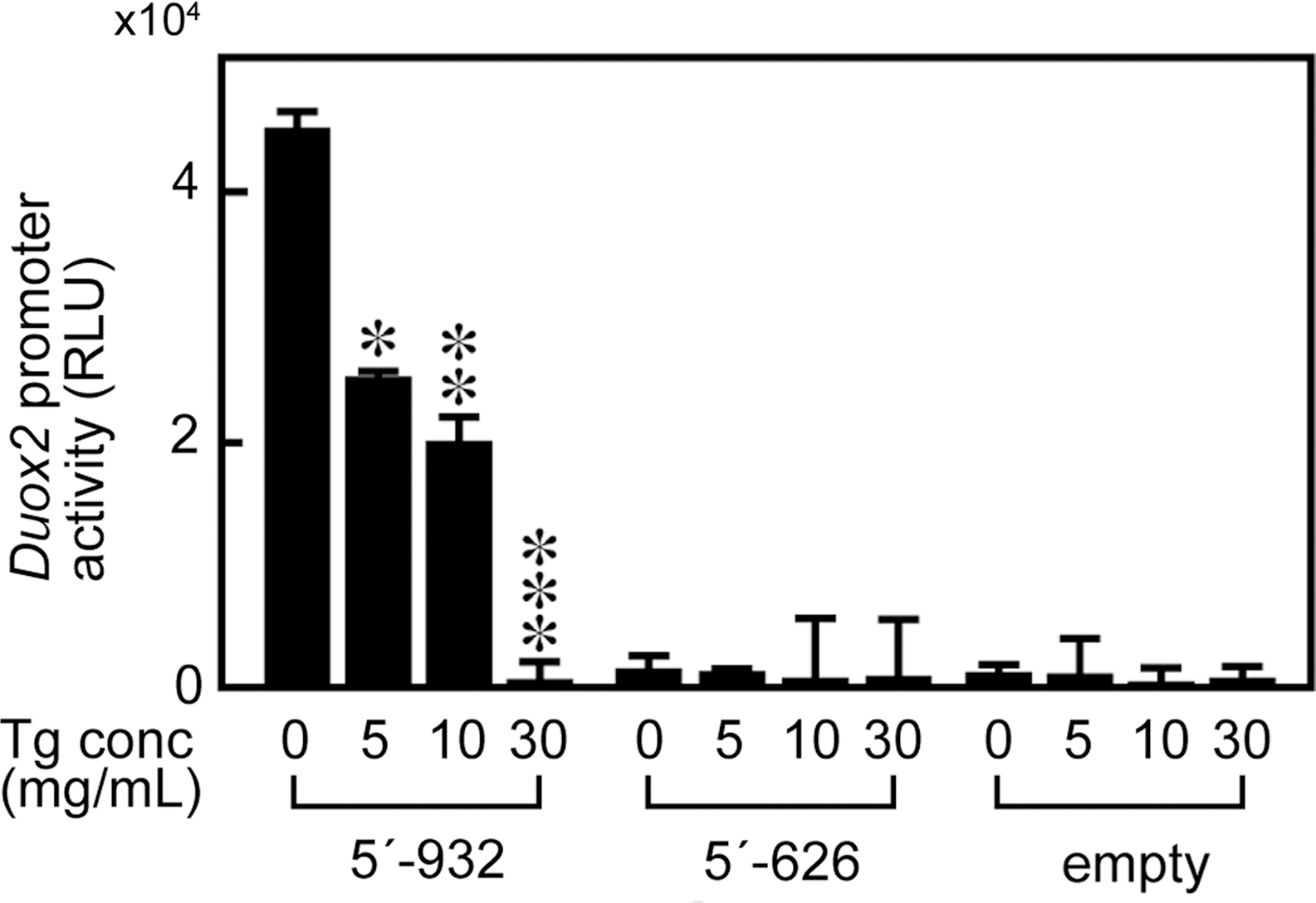
Tg suppressed DUOX2 promoter activity. Luciferase reporter gene constructs containing the full-length (5′-932) or the truncated (5′-626) DUOX2 promoter fragment were transiently transfected in FRTL-5 cells. The cells were then stimulated with 5–30 mg/mL of Tg as indicated, and luciferase activities were measured 48h after Tg stimulation. Data were normalized by corresponding protein concentrations. Luciferase activity is expressed as means±SD of RLU from four samples. *p<0.05, **p<0.01, ***p<0.001 vs. no treatment with Tg. RLU, relative light units.
Tg decreased protein levels of DUOX2
It was demonstrated that Tg suppressed transcription and mRNA expression of Duox2 in FRTL-5 cells. To investigate whether Tg also modulated the protein levels of DUOX2, we performed Western blot analysis using the goat anti-DUOX2 antibody. FRTL-5 cells cultured in the presence of TSH, insulin, and 5% serum were treated with 10 mg/mL Tg for 3–48 hours, and then, total cellular protein was extracted and subjected for Western blotting. The 175 kDa DUOX2 protein was clearly detected in cells before Tg treatment; however, the expression decreased after 6 hours and kept decreasing until 48 hours (Fig. 7A). A densitometric image analysis of the blot showed a gradual decrease in the DUOX2 protein levels after Tg treatment (Fig. 7B), which corresponded with the decrease in mRNA levels shown in Figure 3.
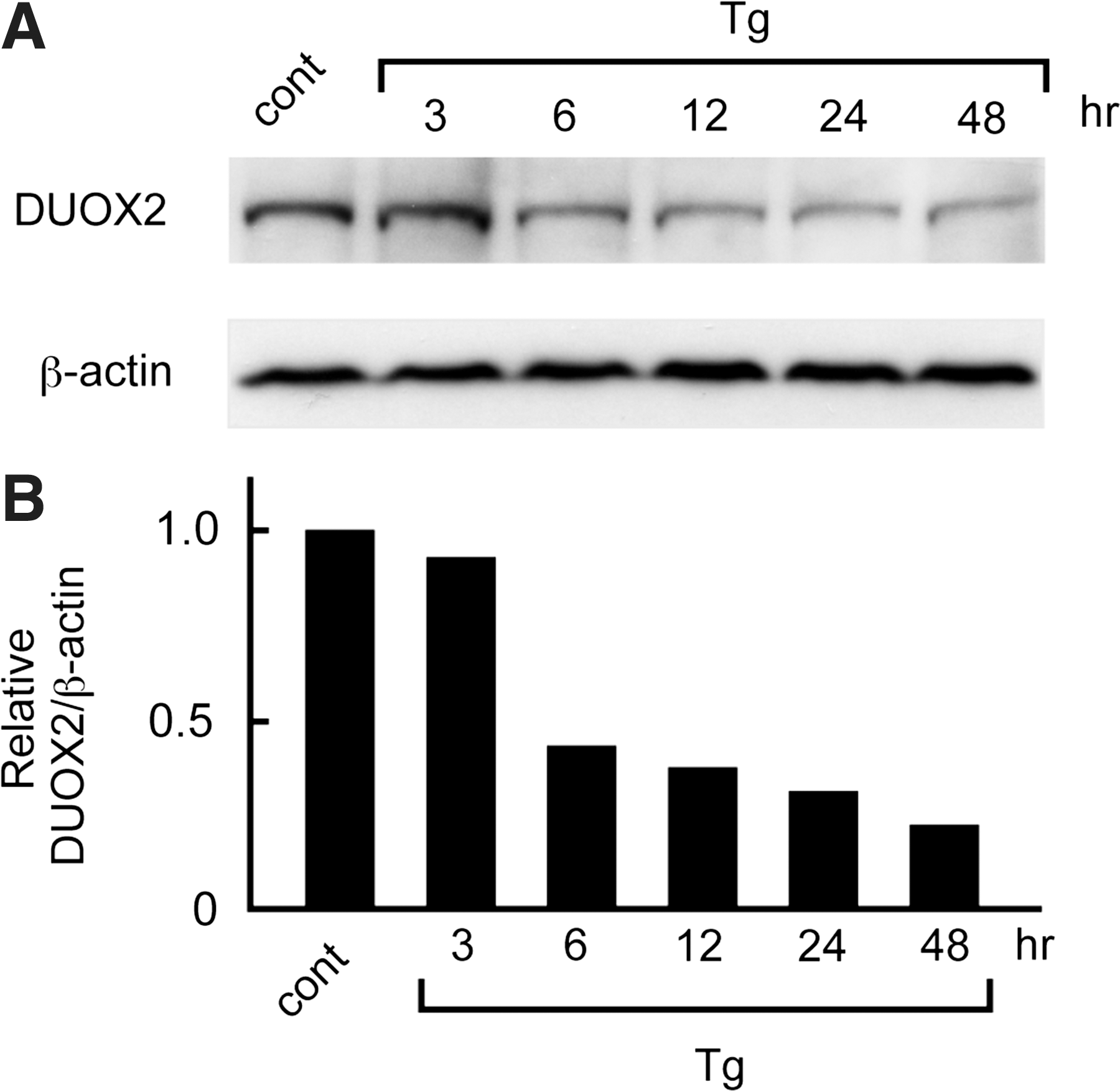
Tg suppressed DUOX2 protein levels in FRTL-5 cells. FRTL-5 cells cultured in the presence of TSH, insulin, and 5% serum were treated with 10 mg/mL of Tg. The effect of Tg on the reduction of DUOX2 protein levels was evaluated by Western blot analysis
Tg suppressed H2O2 generation in FRTL-5 thyroid cells
DUOX2 plays a major role in H2O2 production that is necessary for iodide organification and thyroid hormone synthesis (2 –4). We have shown in the present study that Tg down-regulated the expression of Duox2 and Duoxa2 mRNAs and proteins as just described. To evaluate whether the actual activities of DUOX2 and DUOXA2 were controlled by Tg, we monitored H2O2 generation in the cells using a fluorescent probe, CM-H2DCFDA, which is oxidized in the presence of H2O2 to become fluorescent DCF. As a result, DCF fluorescence significantly decreased 12 hours after the addition of Tg, and the decrease continued with further incubation (Fig. 8, ●). The same concentrations of BSA had no effect on H2O2 generation (Fig. 8, ○), suggesting that the effect is specific to Tg and nonspecific to protein concentrations. Since this method measures intracellular H2O2 levels, we also measured extracellular H2O2 levels using the Amplex Red Hydrogen Peroxide/Peroxidase Assay Kit (Invitrogen). Extracellular H2O2 levels in control FRTL-5 cells were much higher when measured in the presence of Ca2+ (Fig. 8B), indicating that the H2O2 was produced by DUOX2 and not derived from mitochondria (20,46). Tg treatment for 24 hours significantly decreased extracellular H2O2 levels (Fig. 8B).
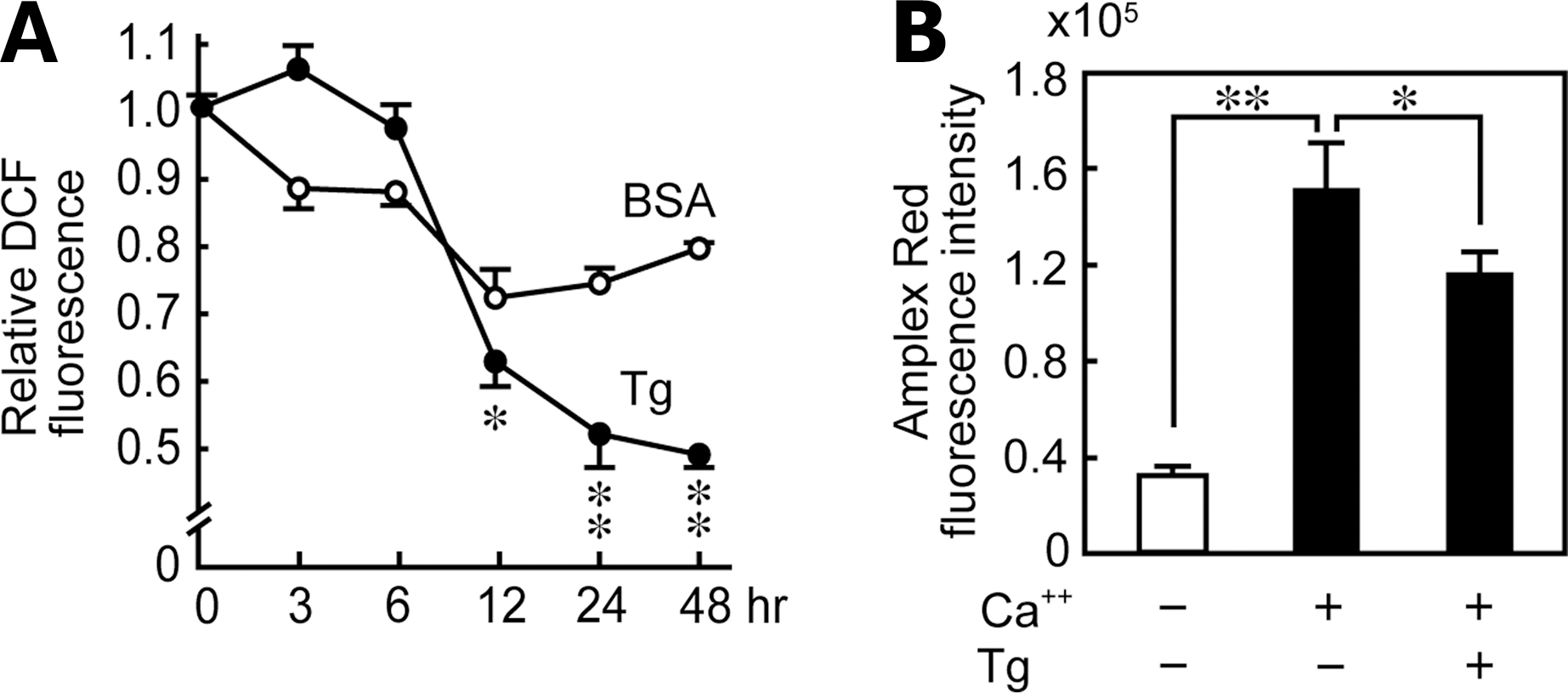
Tg suppressed H2O2 production in FRTL-5 cells.
Discussion
DUOXs are the major enzymes generating H2O2, which is necessary for iodide organification by TPO in thyroid follicles. In the present study, we have shown that levels of Duox2 and Duoxa2 mRNAs were significantly reduced by the physiological concentration of Tg in the presence of TSH, insulin, and serum. The suppression of Duox2 was controlled at the transcriptional level. DUOX2 protein levels and the resultant H2O2 generation were also reduced by Tg. Although the expression and function of most thyroid-specific genes that are necessary for hormone synthesis are tightly regulated by TSH and the Tg stored in each thyroid follicle (28,30,32,33), the regulation of Duoxs and Duoxas by Tg has not been investigated earlier.
Thyroid hormone biosynthesis requires several steps and involves coordinate expression and function of thyroid-specific genes, such as Slc5a5 (NIS), Slc26a4 (pendred syndrome gene), Tg, and Tpo. Although overall thyroid function is tightly controlled by TSH, follicular function is remarkably heterogeneous (27,32,47,48). The function of each follicle is regulated by the amount of Tg stored in the lumen, the action of which counteracts the action of TSH (25,28,30). Thus, the Tg stored in individual thyroid follicles is a potent negative feedback regulator of follicular function that mediates the transcriptional suppression of genes which are necessary for iodide transport and hormone biosynthesis (25,28,29,32). We, therefore, proposed that this is the basis of the follicular heterogeneity under constant TSH stimulation for every follicle (24 –29,31). The concentrations of Tg in normal follicles range from 0.1 to 250 mg/mL. The value obtained depends on the method of measurement (aspiration biopsy or micropuncture of a single follicle) (41 –43). The relatively large range of the estimates of the follicular Tg concentration again supports the significant functional heterogeneity among follicles.
Our present study of Tg's regulation of the expression and function of DUOX2 and DUOXA2 adds novel features to the model just mentioned. In a follicle where the stored Tg is absorbed and only a low concentration remains, all the thyroid gene expressions that are necessary for hormone synthesis are maximum. Oxidation and organification of iodide is achieved by TPO using H2O2 produced by DUOX2. When sufficient Tg accumulates within a follicle, the expression of Duox2 and Duoxa2 along with other essential genes is suppressed by the negative feedback action of Tg that shuts down the iodide organification process, in addition to stopping iodide uptake and transportation (24,25,28). Dietary iodide will be utilized by other follicles where hormone synthesis is actively taking place (24,25,28).
DUOXs were initially cloned from a thyroid cDNA library using gp91phox as a probe based on the functional similarities between H2O2 generation in leukocytes and the thyroid (49). It was shown that their expression had exclusive thyroid specificity, and they co-localized with TPO (9). The H2O2 production by DUOX2 requires a maturation factor (DUOXA2) that is co-expressed with DUOX2 and permits the translocation of DUOX2 from the endoplasmic reticulum to the Golgi apparatus, and its subsequent maturation and translocation to the apical membrane. Recent studies revealed that the expression of Duox2 mRNA is rapidly stimulated by TSH through the cAMP pathway in human, porcine, canine, and FRTL-5 cells (1,4,23,50).
The crucial role of DUOX2 has been demonstrated in patients suffering from a partial iodide organification defect and CH caused by mutations in the Duox2 gene, whereas monoallelic inactivation of the Duox2 or Duoxa2 gene is typically associated with transient CH (14 –19). Abnormalities of H2O2 generation and iodination of Tg may be related to the development of papillary carcinoma or Hashimoto's thyroiditis (51,52). Since the generation of H2O2 by DUOXs is indispensable for iodide organification, abnormalities of the Tg-mediated regulatory mechanism may also be involved in various pathologic consequences of thyroid diseases.
In this study, we have demonstrated that Tg regulates the expression of Duox2 and Duoxa2, but not of Duox1 or Duoxa1. Whether or not this difference in Tg action on the regulation of Duox1 and Duox2 is related to their functional redundancy is not clear. In addition, the underlying molecular mechanisms of Tg action and the cellular recognition system of Tg are still largely unknown. We have shown in this article that the activity of the DUOX2 promoter is suppressed by Tg, although further studies are required to determine the exact basal promoter and Tg-responsive element. Significantly, the rat DUOX promoter was recently characterized (53). Thus, a detailed study should be able to determine the responsible transcription factors regulating the DUOX2 promoter, data that will enhance our understanding of the mechanism of Tg action. In addition to this, we are currently trying to identify Tg binding proteins using electrospray ionization mass spectrometry analysis (45). A better understanding of Tg's role in the regulation of DUOX expression and in the process of thyroid hormone biosynthesis, especially using human primary cells, may enhance our understanding of thyroid physiology as well as the pathogenesis of thyroid diseases.
Footnotes
Acknowledgment
This work was supported in part by a Grant-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (#15390296 and #21591187 to K.S.).
Disclosure Statement
The authors have nothing to disclose.