
Abstract
Background:
Thyroid hormone regulates a wide range of cellular activities, including the balance between cell proliferation and differentiation. The thyroid-hormone-inactivating type 3 deiodinase (DIO3, D3) has been shown to be reactivated in human neoplasias. Here, we evaluated DIO3 expression in human papillary thyroid carcinoma (PTC).
Methods:
Tumor and surrounding normal thyroid tissue were collected from 26 unselected patients with PTC. Clinical data were retrospectively reviewed in medical records. DIO3 mRNA levels were measured by real-time polymerase chain reaction and D3 activity by paper-descendent chromatography. Studies of DIO3 gene regulation were performed in a human PTC-derived cell line (K1 cells). BRAF V600E mutation was identified in DNA from paraffin-embedded tissues by direct sequencing. Immunohistochemistry analyses were performed using a specific human D3 antibody.
Results:
Increased D3 activity was detected in all 26 PTC samples analyzed as compared with adjacent thyroid tissue. The augmentations in D3 activity were paralleled by increased DIO3 mRNA levels (approximately fivefold). In PTC-derived cells, DIO3 transcripts were further upregulated by the transforming growth factor β1 (TGFβ1). Interestingly, preincubation with mitogen-activated protein kinase (MAPK) cascade inhibitors U0126 (ERK pathway) and SB203580 (p38 pathway) decreased DIO3 mRNA levels and blocked the TGFβ1-induced increase in DIO3 transcripts, suggesting that D3 induction might be mediated through the MAPK signaling pathway. Accordingly, DIO3 mRNA and activity levels were significantly higher in BRAF V600E-mutated samples (p=0.001). Increased D3 activity was correlated with tumor size (r=0.68, p=0.003), and associated with lymph node (p=0.03) or distant metastasis (p=0.006) at diagnosis. Conversely, decreased levels of the thyroid-hormone-activating type 2 deiodinase (DIO2) gene were observed in PTC, which might contribute to further decreases in intracellular thyroid hormone levels. Increased D3 expression was also observed in follicular thyroid carcinoma but not in medullary or anaplastic thyroid carcinoma samples.
Conclusions:
These results indicate that the malignant transformation of thyroid follicular cell toward PTC promotes opposite changes in DIO3 and DIO2 expression by pretranscriptional mechanisms. The association between increased levels of D3 activity and advanced disease further supports a role for intracellular triiodothyronine concentration on the thyroid tumor cell proliferation or/and dedifferentiation.
Introduction
Thyroid hormone influences a wide variety of biological events, including the balance between proliferation and differentiation. The activation of the prohormone thyroxine (T4) into the biologically active hormone triiodothyronine (T3) is catalyzed by type 1 (D1) and type 2 (D2) iodothyronine deiodinases via outer-ring deiodination (11). In contrast, type 3 iodothyronine deiodinase (D3) catalyzes the inactivation of both T4 and T3 via inner-ring deiodination. D2 plays a critical role in providing local T3 to regulate intracellular T3 concentration and also seems to be an important source for circulating T3 (11). The main physiological role of D3 is protecting tissues, particularly those in the developing fetus, from inappropriate high levels of T3.
Although the role of deiodinases in neoplasias has not been fully understood, studies focusing on DIO1 and DIO2 genes have reported changes on the expression of these enzymes in benign and malignant tumors (12,13). DIO1 and DIO2 mRNA levels are diminished or unaltered in the majority of thyroid neoplasias, with the exception of augmented D2 activity in follicular thyroid carcinoma (FTC) and medullary thyroid carcinoma (MTC) (14,15). D3 has been shown to be reactivated in human benign neoplasias, such as hepatic haemangiomas, as well as in malignant brain tumors (16). More recently, the increased D3 expression has been described in human malignant basal cell carcinoma (BCC). Interestingly, augmented D3 activity resulted in increased levels of cyclin D1 and cell proliferation whereas Dio3 knockdown caused a fivefold reduction in the growth of tumor xenografts (17). These findings suggest a role for D3 in cell proliferation and might indicate that the local hypothyroidism, due to the augmented D3 activity, could favor tumor cell growth.
Here we have investigated the expression of D3 in human PTC. Increased levels of D3 activity were observed in all PTC samples analyzed. The induced DIO3 expression occurred at transcriptional level and PTC tumors carrying the BRAF V600E mutation had the highest levels of D3 activity. We observed a positive correlation between tumor size and D3 activity. Moreover, increased D3 activity in tumor samples was associated with advanced disease at diagnosis.
Materials and Methods
Patients and tissues
Tumor and surrounding normal thyroid tissue were collected from 26 unselected patients diagnosed with PTC attending the Endocrine or Head and Neck Surgery Divisions at Hospital de Clínicas de Porto Alegre, Brazil. Sample tissues were immediately frozen in liquid nitrogen and stored at −70° until analysis. Surgery was independently indicated by attending physicians. Tumors were histologically classified according to WHO recommendations (18). The clinical stage was determined by the tumor/node/metastasis system (19). Clinical data were retrospectively reviewed in medical records. The study was approved by the Ethical Committee of the Hospital de Clínicas de Porto Alegre, Porto Alegre, Brazil.
Immunohistochemistry studies
Eight PTC (randomly selected from the initial 26 patients), 4 FTC, 10 MTC, and 1 anaplastic thyroid carcinoma (ATC) samples were used for immunohistochemistry studies. Immunohistochemistry analysis was performed on 6-μm sections of previously formalin-fixed and paraffin-embedded tissues. The routine immunohistochemical technique comprised deparaffinization and rehydration, antigenic recovery, inactivation of endogenous peroxidase, and blockage of unspecific reactions. D3 antibody 676 was kindly provided by Dr. Domenico Salvatore (University of Naples). Primary antibodies were incubated overnight at a temperature of 4°C, at dilutions of 1:400 for D3 (the same antibody concentration was also used for positive control) followed by subsequent incubation with biotinylated secondary antibody, streptavidin horseradish peroxidase conjugate (LSAB; Dako Cytomation, Inc.), and diaminobenzidine tetrahydrochloride (Kit DAB; Dako Cytomation, Inc.). Placenta tissue was used as a positive control. The negative control was obtained by omission of the primary antibody. The slides were examined using an Olympus BX51 microscope with an Olympus QColor 5 camera. The QCapturePro software was used to capture the images.
Cell culture
Studies to evaluate DIO3 expression were performed in a human PTC-derived cell line that harbors BRAF V600E mutation (K1 cells, purchased from HPA Culture Collections). Cells were grown in Dulbecco's modified Eagle's medium (DMEM):Ham's F12:MCDB105 (2:1:1) plus 2 mM glutamine and 10% fetal bovine serum. Cells were maintained at 37°C in a humidified atmosphere of 5% CO2 and 95% air, and the culture medium was changed three times a week.
To evaluate the DIO3 regulation, K1 cells were incubated for 18–24 hours in the appropriate medium above plus 0.1% bovine serum albumin (BSA) and 10–200 nM T3, 100 nM selenium, 1 mM dibutyril cAMP (Bu) 2cAMP (DBT), 20 ng/mL transforming growth factor β1 (TGFβ1), or 20 μM iopanoic acid as indicated. Additional studies evaluating DIO3 gene expressions were performed in the presence of 5 μM MAPK pathway inhibitors (U0126, ERK1/2 kinase inhibitor; SB203580, p38 inhibitor). Controls were incubated with medium+vehicle (2% DMSO). At the appropriate times, cells were harvested and processed for total RNA extraction or measurement of D3 activity. All reagents were obtained from Sigma-Aldrich or Invitrogen Life Technologies. All experiments were performed in triplicate in at least two independent experiments.
Assay of D3 activity
D3 activity in tissue samples and cells was determined using paper chromatography as previously described (20,21). Tissue or cells were individually homogenized and sonicated with 10 mM Tris-HCl, 0.25 sucrose buffer (pH 7.5), and 10 mM dithiothreitol (DTT). Protein concentration was quantified by Bradford assay using BSA as a standard. The homogenates were incubated for an hour with 200,000 cpm 125I-labeled T3, 2 nM T3, 20 mM DTT, and 1 mM propylthiouracil (PTU) in order to inhibit any D1 activity. The addition of 200 nM of T3 completely abolished D3 activity in all samples. The reaction was stopped by adding 200 μL ethanol 95%, 50 μL NaOH (0.04 N), and 5 mg PTU. Deiodination was determined based on the amount of 125I-3, 3′-T2 produced after separation of reaction products by paper chromatography. Results were expressed as the fraction of T2 counts minus the nonspecific deiodination (always <1.5%), obtained with the saturating concentration of T3 (200 nM). D3 activity was expressed as femtomoles T3 per minute per milligram protein. The quantity of protein assayed was adjusted to ensure that <30% of the substrate was consumed. All experiments were performed in triplicate in at least two independent experiments.
Real-time polymerase chain reaction
Total RNA was extracted from normal, tumor tissue or cells using Trizol Reagent and the cDNAs generated were used in a real-time polymerase chain reaction (PCR) with an SYBR Green PCR Master Mix (Applied Biosystems) in ABI Prism 7500 Sequence Detection System (Applied Biosystems). Standard curves representing 5-point serial dilution of cDNA of the experimental and control groups were analyzed and used as calibrators of the relative quantification of product generated in the exponential phase of the amplification curve. The r 2 was >0.99, and the amplification efficiency varied between 80% and 100%. Samples were measured by relative quantification (change in expression in the experimental group versus control; untreated versus treated cells). The data generated by ABI Prism 7500 system SDS software (Applied Biosystems) were then transferred to an Excel spreadsheet (Microsoft), and the experimental values were corrected to that of the cyclophilin A standard. The oligonucleotides used were the following: human DIO2 gene, 5′-ACTTCCTGCTGGTCTACATTGATG-3′ and 5′-CTTCCTGGTTCTGGTGCTTCTTC-3′; human DIO3 gene, 5′-TCCAGAGCCAGCACATCCT-3′ and 5′-ACGTCGCGCTGGTACTTAGTG-3′; cyclophilin A (internal control), 5′-GTCAACCCCACCGTGTTCTTC-3′ and 5′-ACTTGCCACCAGTGCCATTATG-3′.
BRAF V600E mutation analysis
Of the initial group of 26 patients, 17 paraffin-embedded formalin-fixed tissue blocks were available for analysis. The DNA was extracted from 10-μm slides of paraffin-embedded tissue blocks, using the Magnesil Genomic Fixed Tissue System (Promega Corporation) according to the manufacturer's instructions. The BRAF exon 15 was amplified by PCR using specific oligonucleotides: 5′-ACCTAAACTCTTCATAATGCTTGCT-3′ and 3′- CTGATTTTTGTGAATACTGGGAACT-5′. To PCR amplification we used 100–300 ng/mL of DNA in a reaction mix (25 μL) containing 20 mM Tris-HCl (pH 8.0), 50 mM KCl, 2 mM MgCl2, 0.2 mM dNTPs, 0.2 mM of each primer, and 1.25 U Platinum Taq DNA Polymerase (Invitrogen Life Technology). BRAF V600E mutation genotyping was performed using direct sequencing. For sequencing, PCR products were purified using the GFX PCR DNA purification kit (GE Healthcare) and submitted to direct sequencing using the Big Dye™ Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems).
Statistical analysis
Data were expressed as median and interquartile interval (P25-P75) for clinical features unless otherwise indicated. D3 activity was expressed as media±standard deviation. Statistical analysis was performed using Mann–Whitney test or Pearson's correlation coefficient. Statistical significance was assumed for p-value<0.05.
Results
DIO3 is induced in follicular-cell-derived tumors
The clinical and oncological characteristics of the 26 patients with PTC included in this study are detailed in Table 1 and were as follows: the mean age was 38.73±16.18 years and 76.9% were women. The median size tumor was 2.5 cm (0.7–10.5); nine patients (34.6%) had lymph node metastasis while eight (30.7%) had distant metastasis at diagnosis. All patients had normal serum TSH levels (0.4–4 IU/L) at surgery.
TNM, tumor/node/metastasis system; +, positive; −, negative; NA, not available.
D3 activity was significantly increased in PTC samples as compared with the surrounding thyroid normal tissue (p<0.0001; Fig. 1A). The augmentations in D3 activities were paralleled by increases in DIO3 mRNA levels (approximately fivefold, p<0.0001; Fig. 1B), indicating that the DIO3 induction occurs at transcriptional level. Further analysis by immunohistochemistry assay with a specific D3 antibody demonstrated that D3 is almost undetectable in normal follicular thyroid tissue whereas strong positive staining was observed in PTC cells (Fig. 1C, D).
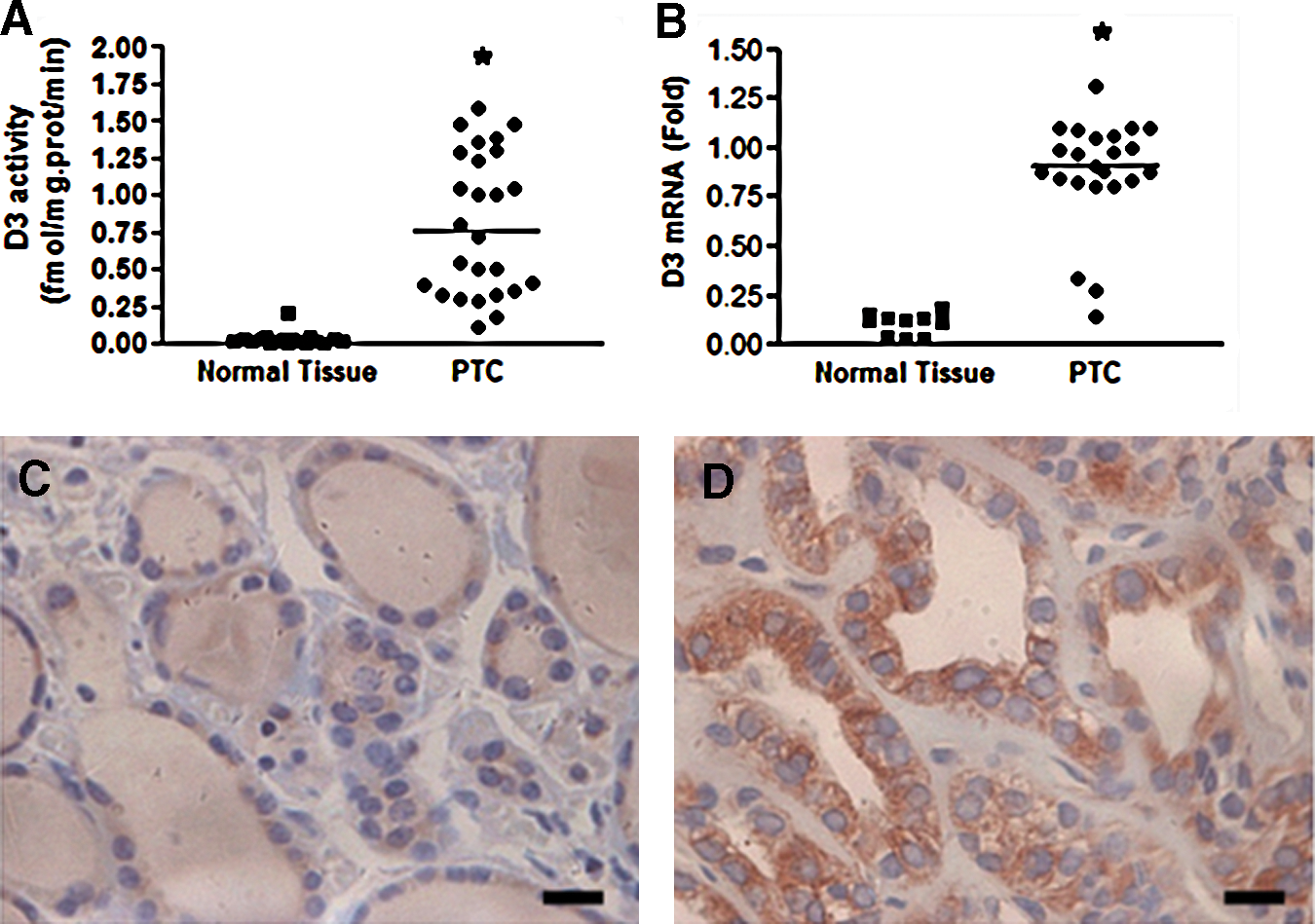
D3 activity
D3 expression in a human PTC-derived cell line
To further evaluate DIO3 expression in PTC, we undertook studies in K1 cells, a human PTC-derived cell line. DIO3 transcripts and activity were readily detected in K1 cells. Additional studies demonstrated that D3 activity was stimulated by selenium (100 nM), T3 (10 nM), or DBT (1 mM). Conversely, D3 activity was abolished by higher T3 doses (200 nM) or iopanoic acid (20 μM) (Fig. 2A).
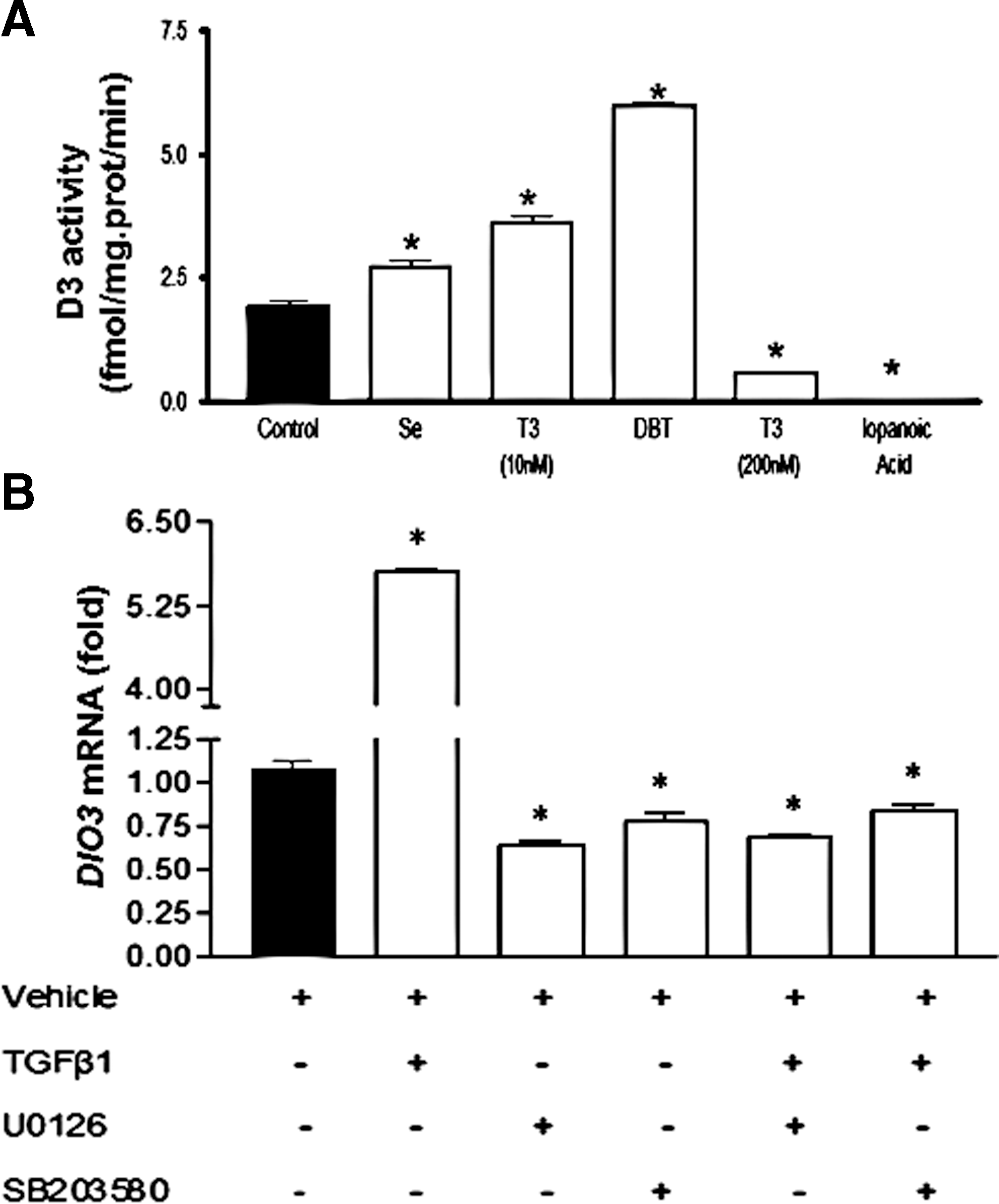
Regulation of DIO3 expression in human PTC-derived cell line.
Next, we investigated the role of TGFβ on DIO3 expression in PTC since it has been previously implicated in DIO3 regulation in other tissues (22). Addition of TGFβ1 (20 ng/mL) to K1 cell culture medium promoted an approximately fivefold induction in DIO3 mRNA levels (Fig. 2B). Interestingly, preincubation with MAPK cascade inhibitors U0126 (ERK1/2 kinase inhibitor) and SB203580 (p38 pathway) decreased DIO3 mRNA levels and blocked the TGFβ1-induced increase in DIO3 transcripts, suggesting that D3 induction might be mediated through the MAPK signaling pathway (Fig. 2B).
DIO3 activity is induced in BRAF V600E-mutated samples
Next we investigated the role of BRAF V600E mutation on DIO3 expression. Of the initial group of 26 PTC patients, 17 PTC samples were available for analysis. Seven (41.2%) out of the 17 PTC samples analyzed were positive for the BRAF V600E mutation. A significant increase in D3 activity was observed in BRAF-mutated as compared with negative samples (p=0.009; Fig. 3A). As expected, the increase in D3 activity was paralleled by an augment in DIO3 mRNA levels (Fig. 3B; p=0.001). No differences in gender, age, tumor size, and local or distant metastasis were observed between patients with or without the BRAF mutation (all p>0.5).
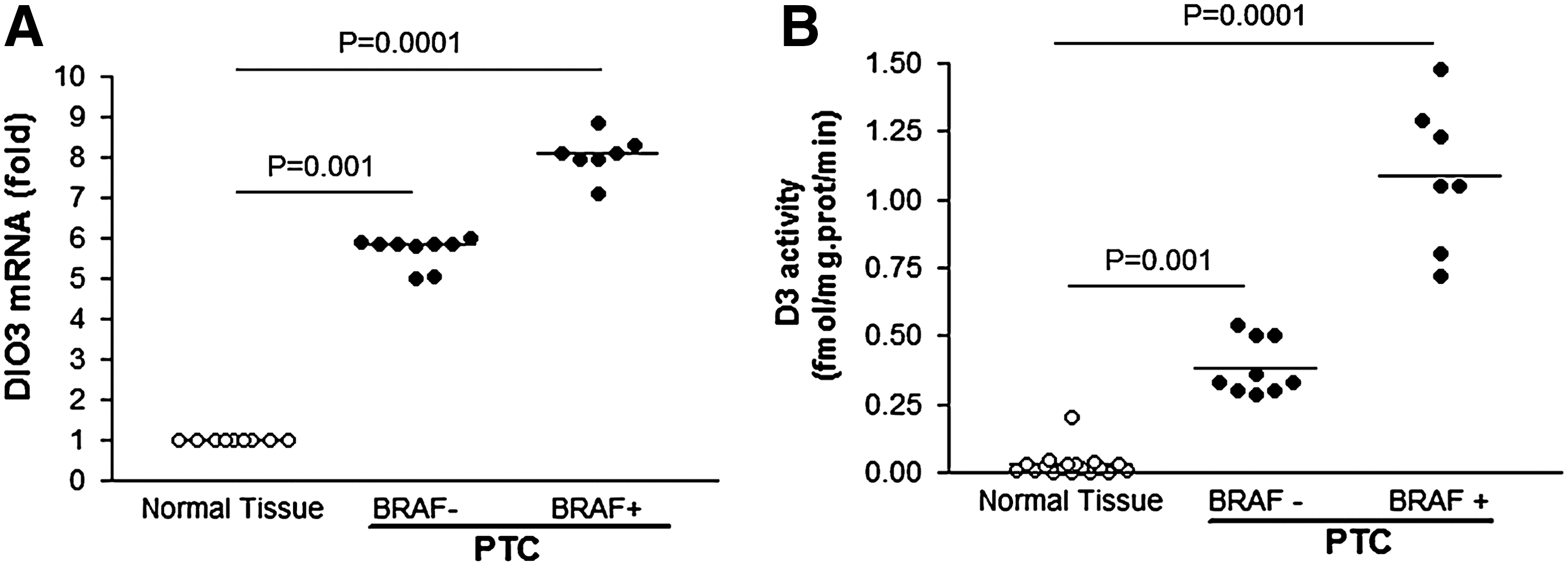
Human PTC samples harboring BRAF
V600E mutation display higher levels of DIO3 mRNA
D3 activity is associated with advanced disease at diagnosis
Previous studies indicated that D3 induction is associated with tumor cell proliferation; thus, we sought to evaluate whether the increase in DIO3 expression in PTC would be associated with disease presentation at diagnosis. Indeed, we observed that increased D3 activity was positively correlated with tumor size (r=0.68; p=0.003; Fig. 4A). Moreover, increased levels of D3 activity in tumor samples were associated with lymph node (p=0.03) or distant metastasis at diagnosis (p=0.006; Fig. 4B), indicating that higher D3 activity might be associated with metastatic disease.
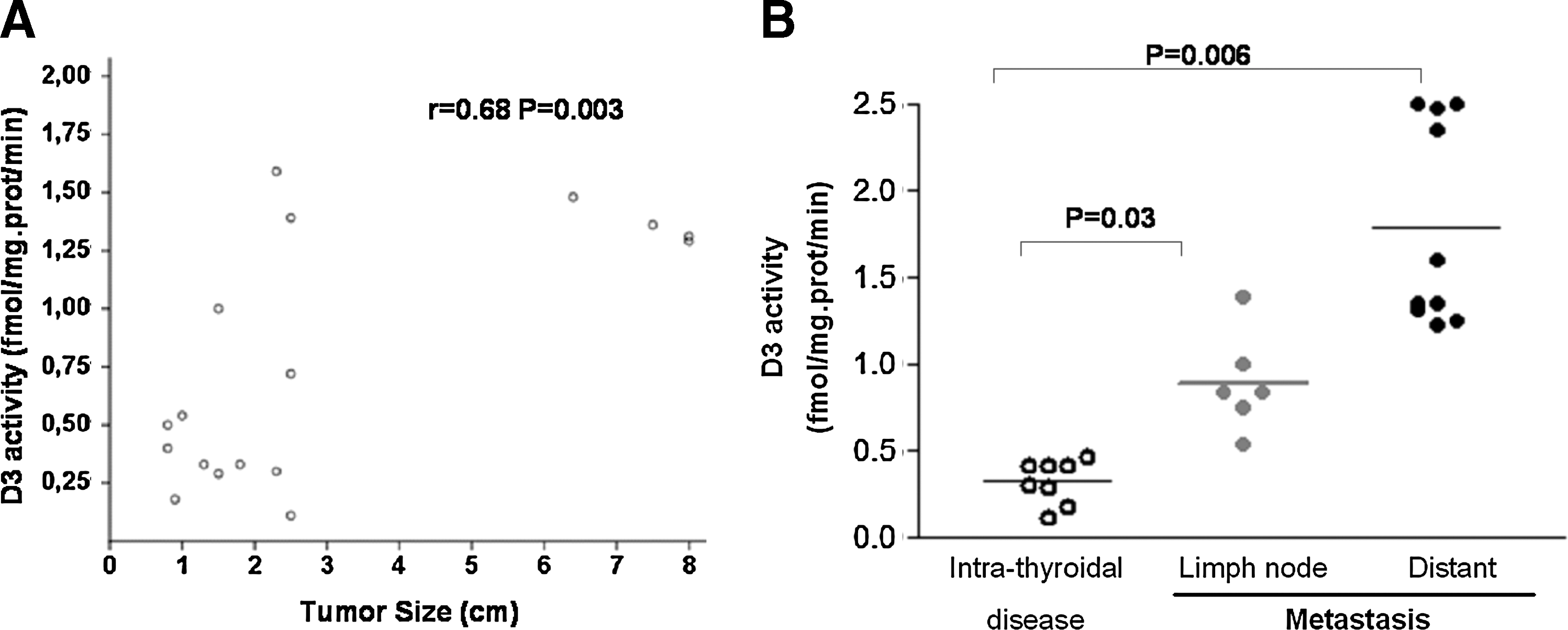
D3 activity levels correlate with tumor size (r=0.68, p=0.003)
DIO2 expression is downregulated in papillary thyroid tumors
Previous studies in BCC have shown that the DIO3 and DIO2 genes are regulated in an opposite fashion. Consistently, we observed a significant decrease in DIO2 mRNA levels in PTC sample as compared with the surrounding tissue (0.18±0.02 vs. 0.85±0.003 AU, respectively; p=0.001).
DIO3 expression in other malignant thyroid neoplasias
We next evaluated the D3 expression in other histological types of thyroid carcinomas. Figure 5 shows the immunohistochemical analysis of a panel comprising representative samples of normal thyroid cells, PTC, FTC, hereditary or sporadic MTC, and ATC. As already shown, D3 staining is barely visible in the normal thyroid tissue (Fig. 5A) while we observe a strong staining in PTC tumor (Fig. 5B). D3 staining is also readily detected in FTC samples, although weaker than in PTC (Fig. 5C). In contrast, D3 protein was barely detectable in an MTC sample of thyroid tissue of a patient with hereditary or sporadic MTC (Fig. 5D, E). No D3 staining was observed in an ATC sample from a 40-year-old man who underwent total thyroidectomy due to an 8 cm×6.5 cm×4 cm fast-growing neck mass (Fig. 5F).
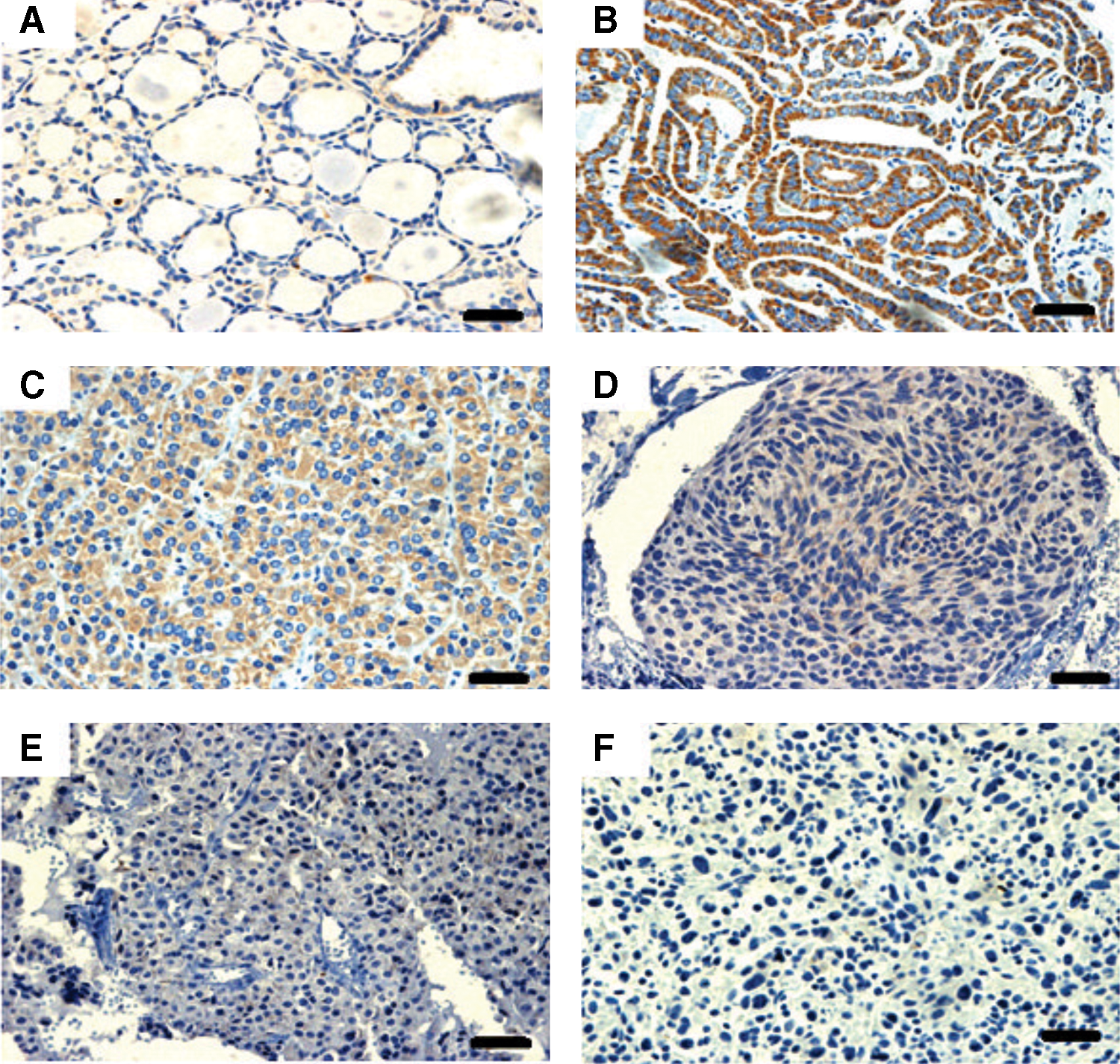
Panel demonstrating D3 immunostaining of representative human thyroid malignant neoplasias. A nearly undetectable D3 expression was observed in thyroid tissue
Discussion
In the present study we evaluated the expression of DIO3 in human PTC. DIO3 transcripts and D3 activity were increased in all PTC samples analyzed. D3 induction is likely to be mediated through the MAPK pathway, which is constitutively activated in this tumor. Accordingly, the highest levels of D3 activity were observed in PTC samples harboring the BRAF V600E mutation. Increased D3 activity was positively correlated with tumor size and associated with disseminated disease at diagnosis. In contrast, DIO2 transcript levels were significantly decreased in PTC as compared with surrounding thyroid tissue. The reduction of D2 activity might contribute to further decrease in intracellular thyroid hormone levels. Taken together, these findings add support for a role of intracellular hypothyroidism in the tumor cell proliferation and/or differentiation.
The ontogenic profile of DIO3 gene shows that D3 activity is present at high levels in developing organs and almost undetectable in most mature tissues. In adults, D3 is predominantly expressed in brain and skin (23,24). Recently, a previously unrecognized role of D3 has been documented in a number of pathological states, unraveling an important role of D3 in both health and disease (16,21,25). Moreover, a potential role of D3 in tumorigenesis has been postulated since induction of DIO3 gene has been observed in several benign and malignant tumors and cell lines (17,25,26).
Here we have demonstrated that DIO3 expression is induced in human PTC. D3 induction occurs at transcriptional level and it is likely to be driven by the constitutive activation of the MAPK pathway, which has been previously implicated in D3 upregulation in other pathological conditions (22,27 –29). Consistently with this hypothesis, we show that samples carrying the BRAF V600E mutation display the highest levels of DIO3 mRNA and activity (Fig. 3B). Based on these observations, we speculate that BRAF-induced increase in DIO3 expression might be involved in the development of aggressive PTC tumors and severe hypothyroidism in thyroid-specific knock-in Braf mice (LSL-BrafV600E/TPO-Cre) (30). Other potential inducer of DIO3 expression in PTC might be the TGFβ, which is overexpressed in thyroid tumors and implicated in cell dedifferentiation and proliferation (31 –33). TGFβ induces DIO3 gene via co-operative effects of Smads and MAPK-dependent pathways. Interestingly, recent studies show that mice with thyroid-specific expression of oncogenic Braf (Tg-Braf) developed PTCs that were locally invasive and had well-defined foci of poorly differentiated thyroid carcinoma. This effect seems to be mediated by oncogenic Braf induction of TGFβ secretion, which in turn might mediate effects as cell migration, invasiveness, and epithelial–mesenchymal transition (34).
A potential role of intracellular thyroid hormone levels on tumorigenesis has long been speculated. It has been demonstrated that thyroid hormone is able to block oncogenic Ras-mediated proliferation and transcriptional induction of cyclin D1 in neuroblastoma cells by interfering specifically with the activity of the MAPK pathway and cyclic AMP response element-mediated transcription (35). Further, studies have indicated a potential association of mutations in thyroid hormone receptor β gene with thyroid carcinogenesis (36,37). More recently, studies performed in BCC demonstrated that D3 induction reduces intracellular thyroid hormone levels and promotes the malignant keratinocyte proliferation (17). The role of D3 was demonstrated by silencing of DIO3 gene that resulted in reduction of both proliferation and cyclin D1 levels, whereas the reintroduction of DIO3 gene in D3-depleted cells reversed these effects. As suggested by the authors, the putative effect of D3 on diminishing microenvironment T3 could provide an advantage for tumor cell proliferation (17). Consistently with these observations, we observed a positive correlation between increased levels of D3 activity and tumor size in human PTC samples. Immunohistochemical staining of D3 protein was also detected in FTC, but, surprisingly, it was nearly undetectable in MTC and ATC samples. These findings were somehow unexpected since the hypoxia-inducible transcription factor (HIF)-1 has previously been shown to be a potent inducer of the DIO3 gene (38). Of note, the highest levels of HIF-1 have been found in ATC samples whereas, in differentiated PTCs and FTCs, its expression was variable (39).
Another interesting observation of this study was the association between increased levels of D3 activity in thyroid tumor samples and metastatic disease at diagnosis. PTC samples from patients with lymph node or distant metastasis displayed significantly higher levels of D3 activity than those samples obtained from patients with intrathyroidal disease (Fig. 4B). This association was independent of the presence of BRAF mutation, which has been associated with aggressive tumor behavior in some studies. One possible explanation for this observation could be an effect of the enhanced D3 activity in the regulation/reactivation, probably through hypothyroidism, of genes responsible for tumor invasiveness and metastasis development. Indeed, several previous studies have demonstrated an association between hypothyroidism and advanced disease. Xenograft tumors of hepatocellular carcinoma (HCC) or breast cancer cells implanted into hypothyroid nude mice were more aggressive and had a higher rate of distant metastasis than those tumors induced into euthyroid mice (40). Additionally, a case–control study observed that individuals with long-term hypothyroidism had a twofold higher risk for HCC development when compared with individuals with no history of thyroid disorders (41). Of interest, a recent study using a human and mouse HCC model has identified a cluster of microRNAs (miRNAs) involved in upregulation of DLK1-DIO3 genomic imprinted region. The authors show that overexpression of DLK1-DIO3 miRNA cluster was associated with higher metastatic rate and poor overall survival in HCC patients (42).
An opposite pattern between increased DIO3 and diminished DIO2 gene expression was observed in PTC samples. Changes on the deiodinase balance with dual convergent mechanisms, transcriptional upregulation of DIO3 and posttranscriptional downregulation of DIO2, were also demonstrated in BCC (17). The reduction of D2 activity might contribute with D3-induced decreases in thyroid hormone levels. Interestingly, DIO2 downregulation seems to be a characteristic feature of papillary carcinomas, since high levels of DIO2 expression have been reported in FTC and MTC (15,43).
In conclusion, we have demonstrated that DIO3 expression is upregulated in PTC. Remarkable, PTC samples harboring BRAF V600E mutation display the highest levels of D3 activity. Increased D3 activity was associated with tumor size and disease spread. These findings further support the concept that the intracellular decrease in thyroid hormone levels might be associated with the degree of tumor cell dedifferentiation and enhancement of tumor invasiveness.
Footnotes
Acknowledgments
This work was supported by Co-ordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Apoio a Pesquisa do Rio Grande do Sul (FAPERGS), and Fundo de Incentivo a Pesquisa do Hospital de Clínicas de Porto Alegre (FIPE), Brazil.
Disclosure Statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.