
Abstract
Background:
Glucose is transported into cells by specific glucose transporter proteins (GLUTs) that are widely expressed in a tissue-specific manner. The mechanisms that regulate glucose uptake and metabolism in thyroid cells are poorly defined. Recently, our group showed that AMP-activated protein kinase (AMPK) plays a pivotal role in the rat thyroid gland, downregulating iodide uptake by thyroid cells even in the presence of its main stimulator thyrotropin (TSH). Since AMPK increases glucose uptake in different tissues, and taken into consideration that in pathophysiological conditions such as thyroid cancer a negative correlation between iodide and glucose uptake occurs, we hypothesized that AMPK might modulate glucose uptake in thyroid cells.
Methods:
Rat follicular thyroid PCCL3 cells cultivated in Ham's F-12 supplemented with 5% calf serum and hormones were exposed to the AMPK pharmacological activator 5-aminoimidazole-4 carboxamide ribonucleoside (AICAR) or AMPK antagonist compound C for 24 hours either in the presence or absence of TSH. Glucose uptake was assessed in vitro using 2-deoxy-D-[3H]glucose.
Results:
AMPK activation by AICAR induced a significant increase in glucose uptake by PCCL3 cells, an effect that was completely reversed by the AMPK inhibitor compound C. Also, the AICAR mediated increase in glucose uptake was detected either in the presence or absence of TSH. The mechanism by which AICAR increases glucose uptake is related to higher levels of GLUT 1 protein content and hexokinase (HK) activity in thyroid cells.
Conclusion:
Our results show that AMPK activation significantly upregulates GLUT 1 content and glucose uptake, and it also stimulates hexokinase activity, the first step of glycolysis.
Introduction
In thyroid cells, only a few studies have evaluated the regulation of glucose uptake and metabolism (2 –6). In rat thyroid cells, three GLUT isoforms (GLUT 1, 3 and 4) have been described, although the most physiological relevant isoform seems to be GLUT 1 (2,3,6). Hosaka et al. reported that FRTL-5 follicular thyroid cells express the mRNA and protein of both GLUT 1 and 4 isoforms, which are positively regulated by thyrotropin (TSH) through the cAMP-PKA signaling pathway (5,6). Also, several studies published in the 1960s and 1970s demonstrated that TSH signaling increased energy and glucose metabolism in thyroid slices (7 –9). However, important species-specific differences regarding intracellular TSH signaling pathways have been reported (10), and TSH was also shown to exert its effects in glucose uptake by activating the phospholipase C-PIP2 signaling cascades in human and rat thyrocytes, and via the PI3K-Akt pathway in rat thyrocytes (3,10).
Recently, we described that the energy sensor AMP-activated protein kinase (AMPK) is expressed in the thyroid gland and plays an important physiological role in thyroid iodide uptake in vitro and in vivo (11). AMPK is a metabolic stress sensing kinase, widely expressed in several tissues, including skeletal muscle, liver, white adipose tissue, and brain (12). Stressors that deplete energy and increase the intracellular AMP:ATP ratio induce allosteric activation of AMPK, promoting conformational changes that lead to the phosphorylation of threonine-172 residue of the enzyme by LKB1, which further activates AMPK. In its activated state, AMPK shuts down processes that consume energy and upregulates energy-producing pathways, a process that tends to restore intracellular ATP levels (12,13).
Apart from the significance of glucose for normal function of thyroid cells, it is well known that malignant thyroid cells depend on the energy obtained from glycolysis (1,14,15). Indeed, a decrease in thyroid cell iodide uptake ability detected in cancer, as demonstrated by a negative 131I whole body scan, is usually accompanied by an increase in glucose uptake, as detected by positron emission tomography scan, using 18 F-FDG (the so-called “flip-flop” phenomenon) (16). This phenomenon is correlated with higher aggressiveness and mortality in thyroid cancer (16,17), although the molecular mechanisms underlying these findings have not been defined. Hence, our aim was to better understand the intracellular pathways involved in the regulation of glucose transport and the first step of its metabolism in thyrocytes.
Since AMPK activation using different pharmacological treatments like metformin and 5-aminoimidazole-4 carboxamide ribonucleoside (AICAR) increases glucose uptake in different tissues (12,13), the hypothesis of this article is that AMPK is involved in glucose uptake regulation by thyroid cells. Here, we provide novel evidence that AICAR induced AMPK-activation increases glucose uptake in thyroid cells by increasing both GLUT 1 expression and hexokinase (HK) activity.
Materials and Methods
Reagents
AICAR was purchased from Toronto Research Chemicals, Inc.; “compound C” (6-[4-(2-piperidin-1-yl-etoxy)-phenyl)]-3-pyridin-4-yl-pyrrazolo[1,5-a]pyrimidine) was kindly provided by Merck. 2-deoxy-D-[3H]glucose from Amersham. A specific antibody against glyceraldehyde-3-phosphate dehydrogenase (GAPDH), enhanced chemiluminescence, and polyvinylidene difluoride (PVDF) membranes were purchased from Millipore. GLUT 1 antibody was obtained from Santa Cruz Biotechnology. Specific antibodies against phospho-AMPK and phospho-ACC were purchased from Cell Signaling. Ham's F-12, and calf serum was from HiMedia. Cytochalasin B was from Sigma. Bovine TSH, insulin, transferrin, hydrocortisone, somatostatin, and Gly-His-Lys were from Sigma. All other chemicals were of the highest grade available.
Cell culture
Rat PCCL3 thyroid cells were a generous gift from Dr. R. Di Lauro (Stazione Zoologica Anton Dohrn). Cells were cultivated in Ham's F-12 medium supplemented with 5% calf serum and containing six hormones (6H): 1 mU/mL bovine TSH, 10 μg/mL insulin, 5 μg/mL transferrin, 10 nM hydrocortisone, 6 nM somatostatin, and 2.5 μM Gly-His-Lys. Cells were maintained at 37°C and 5% CO2 with a change of medium every other day.
PCCL3 cells treatment with AICAR or compound C
PCCL3 cells were grown in TSH-rich medium using 24- and 6-well plates for 48 hours. AICAR was dissolved in Ham's F-12 6H medium (1.5 mM or other concentrations as indicated). Compound C (20 μM) was dissolved in the same medium, added one hour prior and for the entire period of AICAR treatment. To investigate the effects of TSH deprivation in glucose uptake, cells were incubated for 24 hours in Ham's F-12 medium with the same composition but devoid of TSH (5H medium).
Glucose uptake
PCCL3 cells were cultivated in 24-well plates and with AICAR (1.5 mM or as indicated), compound C (20 μM), or both, in the presence (6H) or absence (5H) of TSH for 24 hours. The cells were subsequently washed twice, and glucose transport was assayed in HEPES buffer saline solution (140 mM NaCl, 20 mM HEPES–Na, 2.5 mM MgSO4, 1 mM CaCl2, and 5 mM KCl, pH 7.4) containing 10 μM 2-deoxyglucose (18,500 Bq/mL 2-deoxy-D-[3H]glucose) for 3 minutes as described previously (18). The incubation medium was aspirated, and then cells were washed with ice-cold HEPES buffer and lysed with 1 mL of NaOH (0.05 M). Cell lysates were transferred to scintillation vials for radioactivity counting. An aliquot was used to determine the protein concentration by the Bradford method. Nonspecific uptake was determined in the presence of cytochalasin B (10 μM) and was subtracted from all values.
Western blot analysis of phospho-AMPK, phospho-ACC, and GLUT 1 content
PCCL3 cells were homogenized in lysis buffer containing 135 mM NaCl, 1 mM MgCl2, 2.7 mM KCl, 20 mM Tris, pH 8.0, 1% Triton, 10% glycerol, and protease and phosphatase inhibitors (0.5 mM Na3VO4, 10 mM NaF, 1 M leupeptin, 1 M pepstatin, 1 M okadaic acid, and 0.2 mM phenylmethyl sulfonyl fluoride) and then passed through a syringe five times. Subsequently, the vials were centrifuged and the supernatant was collected. An aliquot was used to determine the protein concentration by the Bradford method. Fifty micrograms of protein were then subjected to sodium dodecyl sulphate–polyacrylamide gel electrophoresis, transferred to PVDF membranes, and probed with the indicated antibodies. The detection of the proteins was performed using an enhanced chemiluminescence reagent (Amersham); GAPDH was used as loading control.
HK activity in PCCL3 cells
PCCL3 cells were seeded in 24-well plates and maintained in 6H medium for 48 hours. After this period, cells were treated with AICAR (1.5 mM or as indicated), compound C (20 μM), or both and in a medium with (6H) or without (5H) TSH for 24 hours. The cells were trypsinized, suspended in phosphate-buffered saline, and frozen at −70°C. HK activity was assayed as previously described (19). Briefly, NADH oxidation or NAD+ reduction were followed by measuring the absorbance at 340 nm in a microplate reader (VICTOR 3; PerkinElmer). Reactions were initiated by the addition of an aliquot of cellular homogenate. An aliquot was used to determine the protein concentration by the Bradford method. Blanks with none of the coupled enzymes were performed to control for nonspecific oxidation/reduction. Each curve was performed in triplicate.
Total RNA extraction and quantitative reverse transcriptase–polymerase chain reaction
The level of rat GLUT 1 transcript (NC_005104.2) was analyzed by quantitative real-time reverse transcriptase–polymerase chain reaction (RT-PCR). Immediately after treatment of the PCCL3 cells, total RNA was extracted and DNase treated using the RNeasy® Plus Mini Kit (Qiagen) following the manufacturer's instructions. Total RNA was reverse-transcribed into cDNA using the High Capacity Reverse Transcriptase Kit (Applied Biosystems). For the quantification of rat GLUT 1 mRNA expression, we have used forward and reverse primers located in different exons, as follows: f-GCTCATCCTGAACCAAGTGA and r-ACGAGCATTACCACAACCTG. Real-Time amplifications were carried out in the ABI Prism 7500 (Applied Biosystems) machine. The amplification reactions were performed in 96-well plates in 12 μL final volume that contained 3 μL of 50× diluted cDNA, 6 μL of 2× Power SYBR Master Mix (Applied Biosystems), and 200 nM of each forward and reverse primer. The amplification program was 55°C 2 minutes, 95°C 10 minutes followed by 40 cycles of 95°C 30 seconds, and 58°C 1 minute. The RT-PCR efficiency was evaluated using serial dilutions of the template cDNA, and melting curve data were collected to determine the RT-PCR specificity. Each cDNA was amplified in triplicate, and a corresponding sample without RT (no-RT sample) was included as negative control. The content of the GLUT 1 transcript was normalized to that of GAPDH. The forward and reverse primers for rat GAPDH were TGATTCTACCCACGGCAAGT and AGCATCACCCCATTTGATGT, respectively. The relative quantities of GLUT 1 expression were determined by the ΔΔCt method expressed by the formula 2(ΔΔCt) (20), where Ct refers to the “threshold cycle” and is determined for each plate by the 7500 Real-Time PCR System Sequence Detection Software (Applied Biosystems).
Statistical analysis
Statistical analyses were performed by one-way analysis of variance followed by the Bonferroni multiple comparison post hoc test; level of significance was set at p<0.05.
Results
Effects of AICAR and compound C in glucose uptake in PCCL3 cells
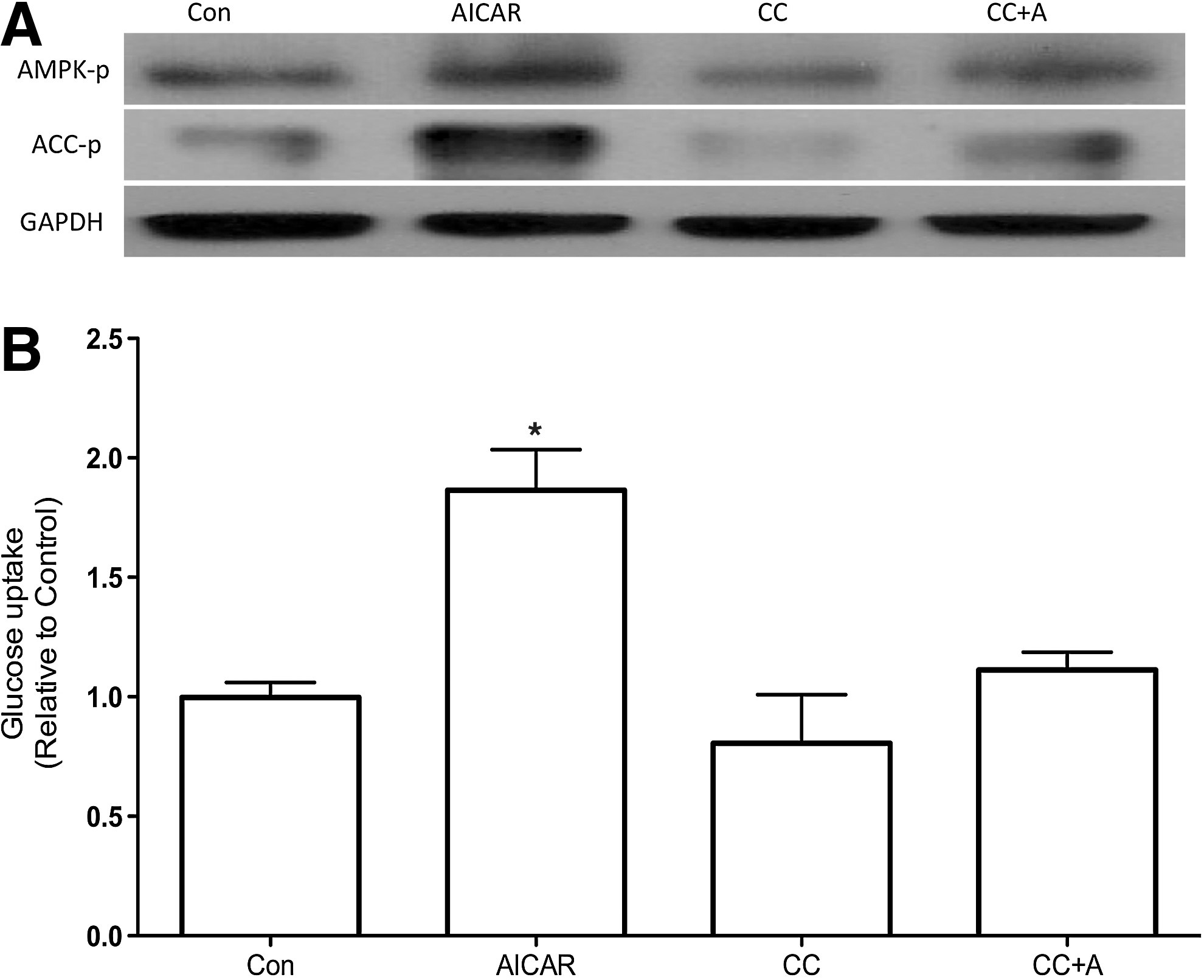
To first analyze the involvement of AMPK in regulation of glucose uptake in thyroid cells, we treated PCCL3 cells with different concentrations of AICAR, which is a pharmacological activator of AMPK. Treatment of PCCL3 cells for 24 hours with AICAR elicited a concentration-dependent increase in glucose uptake reaching values 1.56- and 2-fold higher than control in the presence of 1 and 2 mM of AICAR, respectively (Fig. 1). To investigate whether the effects of AICAR in glucose uptake were indeed mediated by AMPK, we measured phospho-AMPK and its downstream substrate phospho-ACC in PCCL3 cells cultivated in Ham's F-12 medium supplemented with 5% calf serum and containing six hormones (6H). AICAR treatment caused a significant increase in phospho-AMPK and phospho-ACC (Fig. 2A), which accompanied the increase in glucose uptake (Fig. 2B), while compound C fully prevented both, AMPK and ACC phosphorylation and the stimulatory effect of AICAR in glucose uptake.
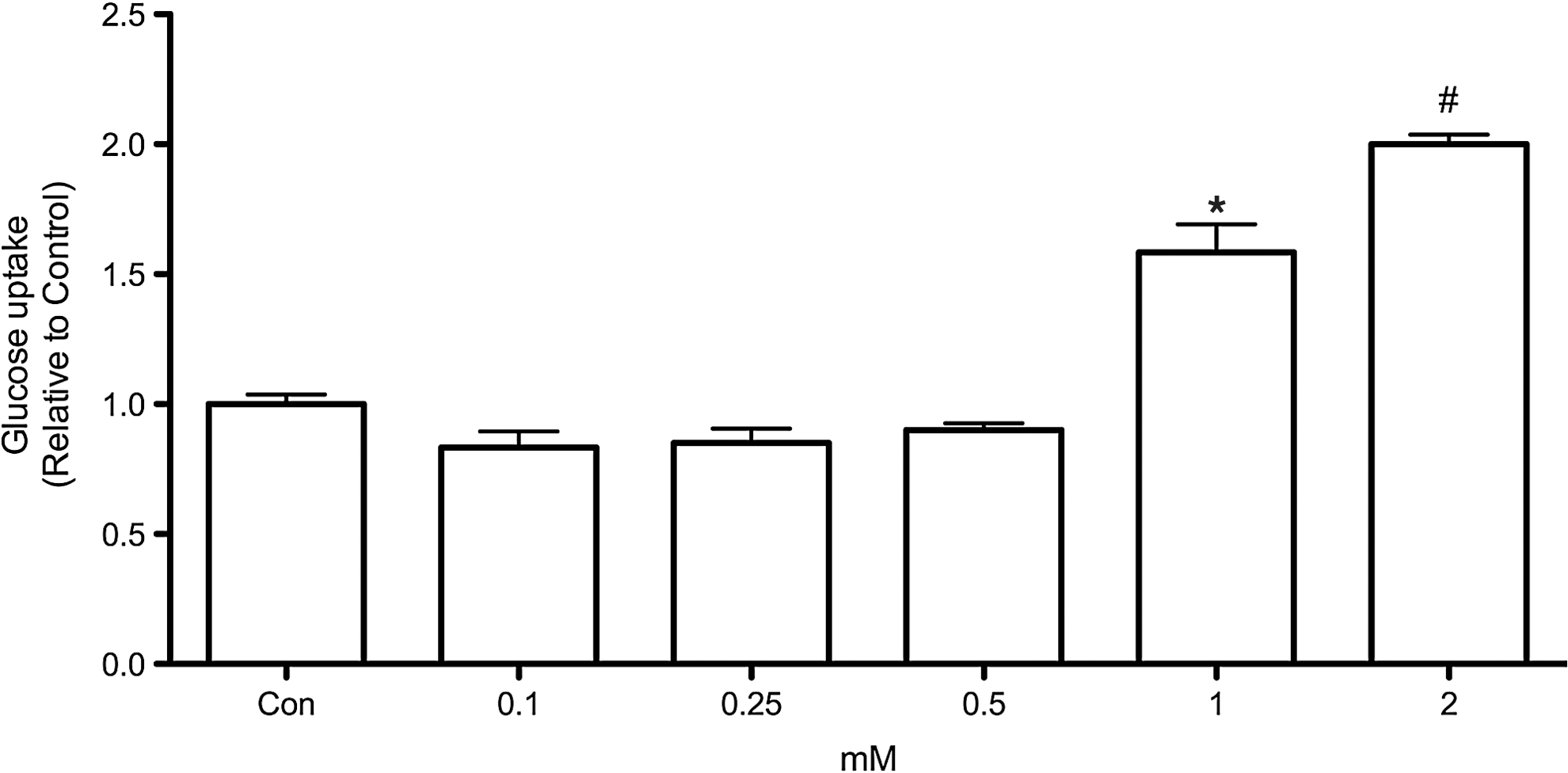
Concentration-response effect of 5-aminoimidazole-4 carboxamide ribonucleoside (AICAR) in glucose uptake by rat PCCL3 cells. Cells were incubated for 24 hours in F-12 medium containing thyrotropin (TSH), insulin, transferrin, somatostatin, hydrocortisone, and Gly-His-Lys acetate in the absence or presence of various AICAR concentrations. Data are expressed as mean±standard error (SE); n=6–7. *p<0.05 versus 0, 0.125, 0.25, and 0.5 mM AICAR. # p<0.01 versus all other conditions (analysis of variance [ANOVA] followed by Bonferroni post-test).
Effects of AICAR on HK activity in PCCL3 cells
Once inside the cell, glucose is immediately phosphorylated by HK, which is the first step of intracellular glucose metabolism. Little is known about HK regulation in thyroid cells; therefore, we evaluated whether AMPK activation could also modulate HK activity in PCCL3 cells. We noted that the incubation with AICAR for 24 hours significantly increased HK activity in thyroid cells (Fig. 3). As expected, compound C also reversed the AICAR-induced increase in HK activity. No changes were observed in HK activity when the cells were cultivated in the absence of TSH (5H medium) for 24 hours.
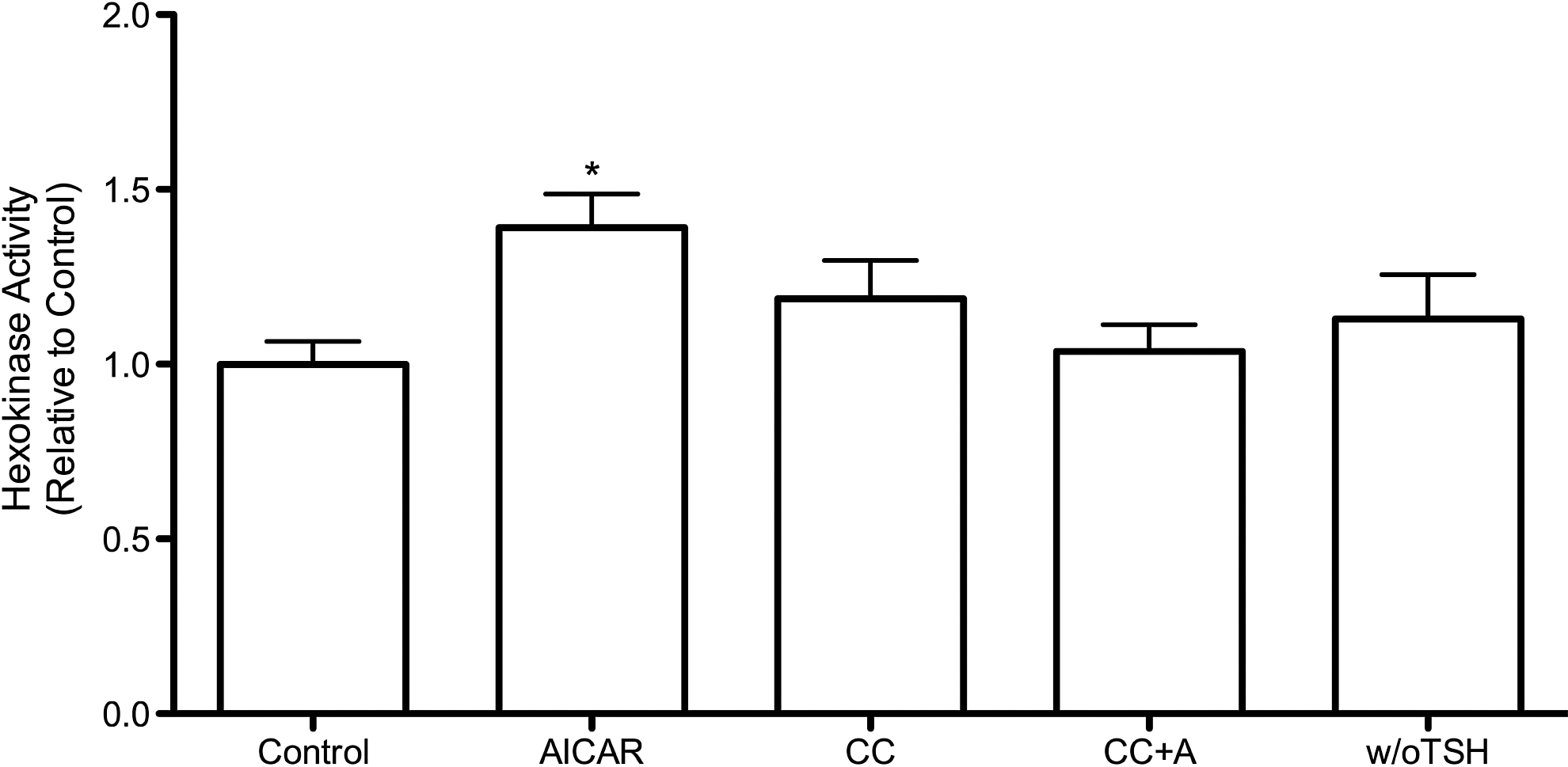
Effects of AICAR and deprivation of TSH on hexokinase (HK) activity in PCCL3 cells. Cells were incubated for 24 hours in F-12 medium containing insulin, TSH, transferrin, somatostatin, hydrocortisone, and Gly-His-Lys acetate. Incubations occurred in the absence (control) or presence of AICAR (1.5 mM), CC (20 μM), and CC plus AICAR (CC+A). Compound C was added one hour before AICAR and was present in the incubation medium during the entire 24-hour treatment period. Cells treated in the absence of TSH (w/o TSH) were incubated for 24 hours in F-12 medium without TSH. Data for HK activity are expressed as mean±SE; n=8–10; *p<0.05 versus all other conditions (ANOVA followed by Bonferroni post-test).
Effects of AICAR in the absence of TSH on glucose uptake and GLUT 1 protein and mRNA total content in PCCL3 cells
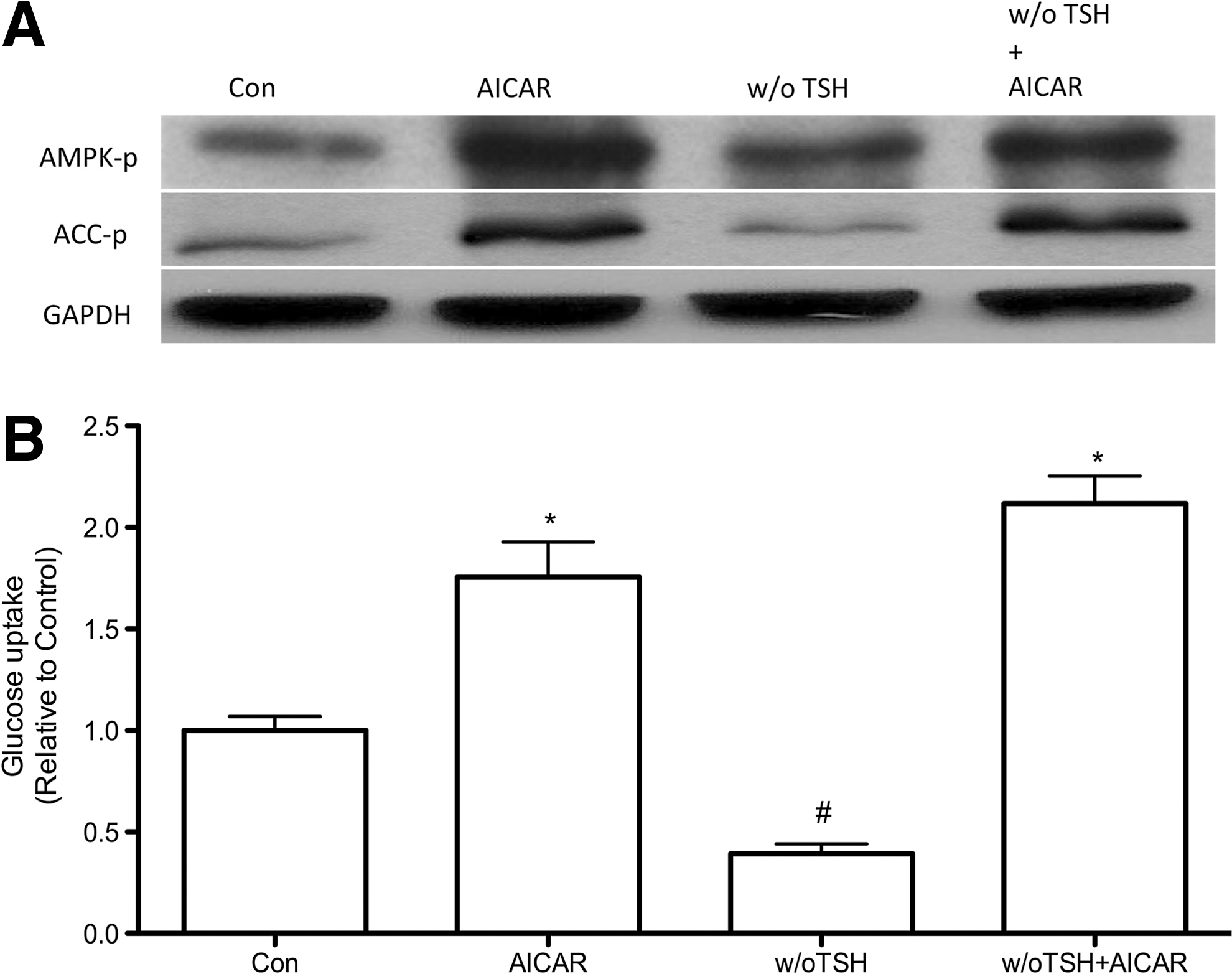
Since TSH is the main known stimulator of glucose uptake in thyroid cells, we analyzed whether the increase in glucose uptake promoted by AMPK activation was dependent on the presence of TSH. Again, in PCCL3 cells treated with AICAR in the presence or absence of TSH for 24 hours, the phosphorylation of AMPK and its substrate ACC were increased (Fig. 4A). Moreover, consistent with the increased AMPK and ACC phosphorylation we found a significant increase in glucose uptake (Fig. 4B). To further establish the mechanism by which AMPK activation by AICAR leads to increased glucose uptake even in the absence of TSH signaling, we measured the total protein and mRNA of GLUT 1. The total content of GLUT 1 protein was significantly increased when cells were treated with AICAR, in the presence or absence of TSH (Fig. 5A, B). In contrast, no significant differences were found in the total content of GLUT 1 mRNA in any of the studied experimental conditions (Fig. 5C).
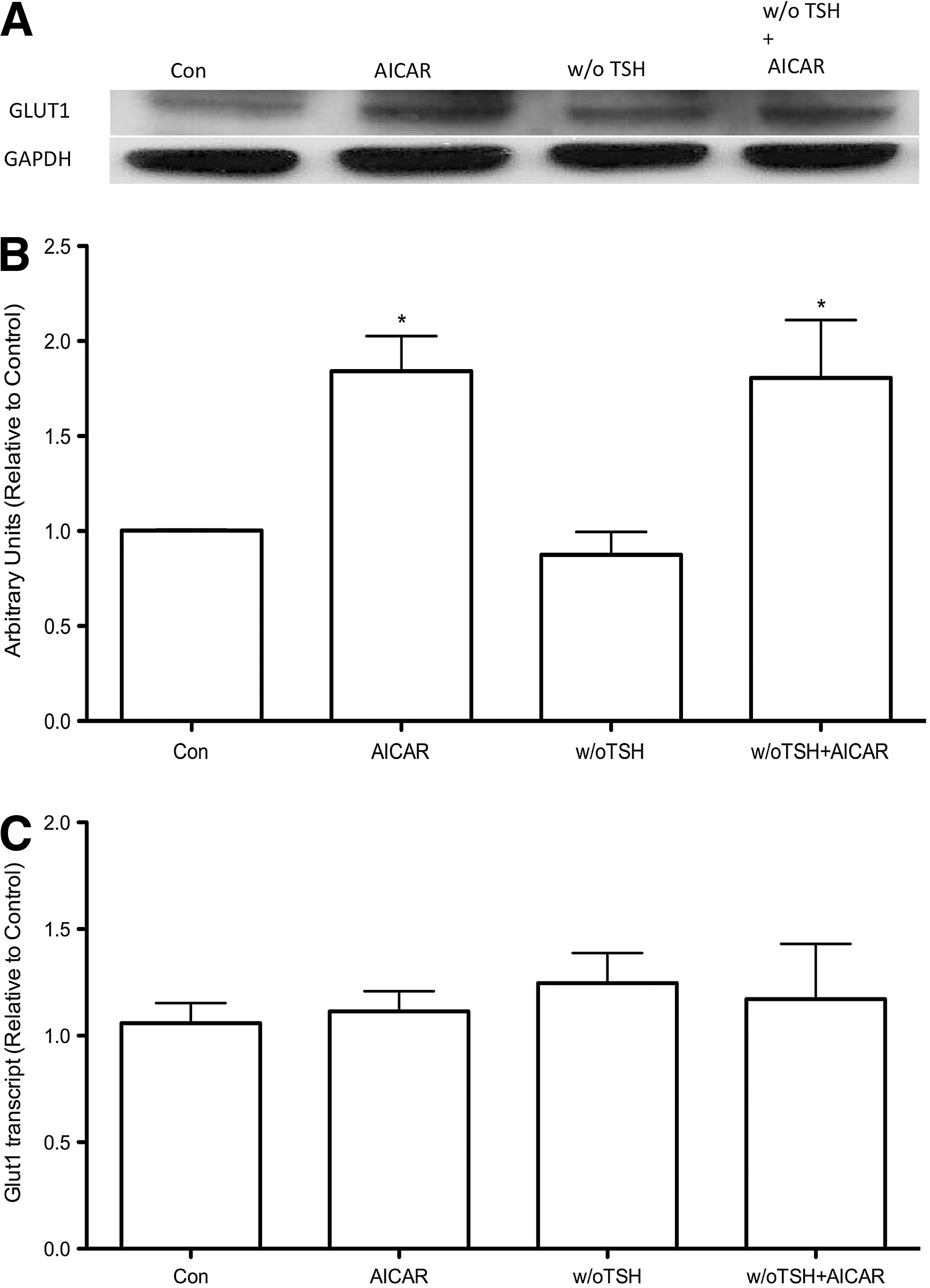
Discussion
The data presented here describe a new intracellular pathway involved in the regulation of glucose uptake by rat thyroid cells. Glucose uptake by PCCL3 cells increases upon AMPK activation by AICAR in a concentration dependent manner, an effect fully reversed by the AMPK inhibitor compound C (Figs. 1 and 2). Previous studies demonstrated that TSH is the main regulator of glucose uptake and metabolism in thyrocytes from different species under physiological conditions (2 –10). The effect of TSH seems to be dependent on the cAMP-PKA and PI3K pathways (3,5 –10). Indeed, Filetti et al. (5) demonstrated that TSH is a potent stimulator of glucose uptake, while Samih et al. (3) demonstrated that the PI3K inhibitor LY294002 blocks the translocation of GLUT 1 to the plasma membrane mediated by TSH and the cAMP membrane permeable analog (Bu)2cAMP. In our study, we confirmed the important role of TSH to induce glucose uptake by thyroid cells. We detected a significant decrease in glucose uptake by PCCL3 cells deprived of TSH for 24 hours, which was accompanied by decreased GLUT 1 protein expression but without changes in HK activity. On the other hand, we describe a novel intracellular pathway that upregulates glucose uptake independent of TSH, since AMPK activation by AICAR significantly increased glucose uptake even in the absence of TSH in PCCL3 cells (Fig. 4). These results are in accordance with our previous report showing that in fact TSH constitutively blocks AMPK activation in thyrocytes through the PKA pathway (11). Thus, the mechanisms underlying AMPK activation in thyrocytes might involve decreased cAMP-PKA signaling pathways; however, AMPK activation through other as yet undefined mechanisms could also account for increased glucose uptake when TSH signaling is downregulated. There are at least three intracellular pathways involved with AMPK activation that depend on kinases such as liver kinase B1 homolog (serine/threonine-protein kinase LKB1), calcium/calmodulin-dependent protein kinase kinase (CaMKK), or TGF-beta-activated kinase 1 (TAK1), as described for other cell types (12). It is tempting to speculate whether these kinases might play a role in the regulation of glucose uptake by thyrocytes.
At least two distinct mechanisms could account for the stimulatory effect of the pharmacological agent AICAR in glucose fixation by PCCL3 cells: an enhancement of glucose transport into cells or a higher glucose phosphorylation rate for subsequent intracellular metabolism (1,15,18). In the present study, we demonstrate that AMPK upregulates both GLUT 1 protein content and HK activity. In FRTL-5 cells, Hosaka et al. (6) showed that TSH increased GLUT 1 and GLUT 4 mRNA levels. In our study, however, we did not find a reduction in GLUT 1 mRNA levels in PCLL3 cells deprived of TSH for 24 hours, but the protocols used are quite different. Nevertheless, AMPK-mediated increase in glucose uptake and GLUT 1 protein content occurred even in the absence of TSH, clearly demonstrating that AMPK involves an alternative pathway regulating glucose uptake by thyrocytes.
It is well established in the literature that an increased glucose uptake associated with decreased iodide uptake ability is a strong indicator of thyroid cancer progression and higher aggressiveness (2,15 –17). Recently, we demonstrated that AMPK is expressed in thyroid cells and that AICAR-mediated AMPK activation decreases iodide uptake both in vitro and in vivo. Taken together, our results show that AMPK might play an important role in the regulation of thyroid function, a finding that suggests that it could be involved in some pathophysiological conditions such as tumorigenesis.
From a physiological perspective, the ability of AMPK to modulate glucose uptake and metabolism independent of TSH signaling might be important for the future identification of novel pathways involved in the regulation of thyroid function.
Footnotes
Acknowledgments
We thank Dr. Rolando Ceddia for his contribution in glucose uptake experiments. This work was supported by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Programa de Oncobiologia da UFRJ, Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Disclosure Statement
The authors have nothing to declare.