
Abstract
Background:
Dual oxidases (DUOX1 and DUOX2) are NADPH oxidases (NOX) involved in hydrogen peroxide production necessary for thyroid hormonogenesis, but recently, the NOX4 has also been described in the thyroid gland. The prevalence of thyroid disease is higher in women, and the basis for this difference might involve a higher oxidative stress level in the female thyroid gland. Hence, we aimed at evaluating whether the function and the expression of enzymes involved in the thyroid redox balance differ between females and males.
Methods:
DUOX1, DUOX2, NOX4, glutathione peroxidase (GPx), and catalase activities and expression levels were evaluated in the thyroids of prepubertal and adult male and female rats. The mRNA levels of DUOXA1 and DUOXA2, the DUOX maturation factors, and of p22phox and Poldip2 (subunits of NOX4) were also determined.
Results:
A higher calcium-independent H2O2 production was detected in the adult female rat thyroid, being higher in the estrous phase of the cycle. Moreover, the expression of NOX4 and Poldip2 mRNA was higher in the thyroids of adult female rats, as well as in PCCL3 cells treated with 17β-estradiol. The GPx1 mRNA expression was higher in adult female thyroids, while GPx2 and GPx3 mRNA and total GPx activity were not significantly different. Catalase mRNA expression and activity, together with thyroid thiol levels were significantly lower in the adult female rat thyroid.
Conclusions:
Taken together, our results show that the thyroid gland of female rats is exposed to higher oxidative stress levels due both to increased reactive oxygen species (ROS) production through NOX4, and decreased ROS degradation.
Introduction
H2O2 is a reactive oxygen species (ROS) that is continuously generated, but is normally maintained at low intracellular concentrations due to the action of peroxidases, such as glutathione peroxidase (GPx) and catalase, as well as other antioxidant mechanisms (7). It is well established that at low concentrations, H2O2 can act as a signaling messenger regulating many cellular processes, but at high concentration it can be toxic to cells. Low H2O2 concentrations stimulate cell proliferation in different cell types. However, incubation of cells with a higher H2O2 concentration can lead to growth arrest and even cell death by apoptosis or necrosis (8), since it can interact with cellular constituents, such as proteins, lipids, carbohydrates, and DNA, inducing their oxidation and disruption of their normal function (9). Maier et al. showed that the thyroid gland is exposed to higher levels of DNA oxidative damage and spontaneous mutations in comparison to other tissues, and postulated that these findings could be due to the oxidative microenvironment of the thyroid due to the presence of constitutively mediated higher H2O2 production (10).
Thyroid diseases, such as autoimmune disease and cancer, in general, are more prevalent in women than in men; however, the mechanisms involved in this sex difference remain elusive (11,12). Since H2O2 is produced in high levels in the thyroid gland and regulates parameters, such as cell proliferation, migration, survival, and death, an imbalance in the cellular oxidant–antioxidant system in the thyroid could contribute to the greater incidence of thyroid disease among women. Nevertheless, no consistent information about a possible sex-related difference in thyroid oxidative stress is found in the literature. Therefore, our aim was to evaluate the activity and expression of some enzymes involved in the production and degradation of ROS, and to compare the cellular redox status in the thyroid glands of male and female prepubertal and adult rats.
Materials and Methods
Animals
In all experiments, prepubertal (30 days old) or adult (3 months old) male and female Wistar rats were fed a regular diet supplied ad libitum. The Institutional Committee for Use of Animals in Research approved the study, and the procedures were in compliance with the International Guiding Principles for Biomedical Research Involving Animals, the Council for International Organizations of Medical Sciences, and the guiding principles for care and use of animals from the American Physiological Society.
Adult animals was divided into three groups: sham-operated (controls), ovariectomized treated with vehicle (OVX), or treated with estradiol benzoate (Eb; 1,3,5[10] estratriene-3,17β-diol 3-benzoate; Sigma-Aldrich), in a physiological dose of 0.7 μg/100 g body weight per day, administered subcutaneously for 7 or 21 days.
Animals were euthanized; thyroid tissue was collected, immediately frozen in liquid nitrogen, and stored at −80°C or kept at 4°C until further processing.
Cell culture and 17β-estradiol treatment
PCCL3 rat follicular thyroid cells (donated by Prof. Roberto Di Lauro, Stazione Zoologica Anton Dohrn) were maintained in the Coon's modified Ham's F-12 medium supplemented with 5% fetal bovine serum and a six-hormone mixture [1 mU/mL TSH, 10 μg/mL insulin, 5 μg/mL transferrin, 10 nM hydrocortisone, 10 ng/mL somatostatin, and 10 ng/mL glycyl-l-histidyl-l-lysine acetate; complete medium (13)]. Cells were starved for 48 hours using a starvation medium (phenol red-free medium supplemented with 0.2% fetal bovine serum and a six-hormone mixture), followed by 17β-estradiol (17β-E; Sigma-Aldrich) incubation for 72 hours in the starvation medium.
Thyroid H2O2 production
H2O2 generation was quantified in thyroid particulate fractions by the Amplex red/horseradish peroxidase (HRP) assay (Molecular Probes; Invitrogen), which detects the accumulation of a fluorescent oxidized product. The excised thyroid glands remained at 4°C for 24 hours in a 50 mM sodium phosphate buffer, pH 7.2, containing 0.25 M sucrose, 0.5 mM dithiothreitol, 1 mM ethylene glycol tetra-acid (EGTA), 5 mg/mL aprotinin, and 34.8 mg/mL phenyl methane sulfonyl fluoride (PMSF) before homogenization. Then, the homogenate was centrifuged at 100,000 g for 35 minutes at 4°C and resuspended in 0.5 mL 50 mM sodium phosphate buffer, pH 7.2, containing 0.25 M sucrose, 2 mM MgCl2, 5 mg/mL aprotinin, and 34.8 mg/mL PMSF and stored at −20°C until H2O2 generation measurements (14). This particulate fraction was incubated in 150 mM sodium phosphate buffer (pH 7.4) containing superoxide dismutase (SOD) (100 U/mL; Sigma), HRP (0.5 U/mL; Roche), Amplex red (50 μM; Molecular Probes), 1 mM EGTA, with or without 1.5 mM CaCl2, and, the fluorescence was immediately measured in a microplate reader (Victor X4; PerkinElmer) at 30°C, using excitation at 530 nm and emission at 595 nm (15). H2O2 production was quantified using standard calibration curves.
Specific enzymatic activity was expressed as nanomoles H2O2 per milligram of protein (nmol/mg) per hour. The protein concentration was determined by the Bradford assay (16).
Measurement of H2O2 generation
Extracellular H2O2 generation was quantified by the Amplex Red/HRP assay (Molecular Probes, Invitrogen), which detects the accumulation of a fluorescent oxidized product. 1×105 cells in Dulbecco's phosphate-buffered saline (PBS) with CaCl2 and MgCl2 were incubated with
Immunostaning for NOX4
Immunohistochemistry for NOX4 (anti-NOX4 primary antibody NB 110-5885; Novus Biologicals) was performed in male and female thyroid tissue. Thyroids were fixed in 4% paraformaldehyde, dehydrated in a graded series of ethanol (70%, 95%, and 100%), and embedded in paraffin. Sections of 5 μm thick were mounted on glass slides, baked at 60°C overnight, deparaffinized using xylene, and redydrated using ethanol (100%, 95%, and 80%) and deionized water. Tissue was incubated at 5% sodium tetraborate for 15 minutes and antigen retrieval was done using the citrate buffer 0.01 M, pH 6.0. Then, tissues were blocked with 5% bovine serum albumin at room temperature and incubated with an anti-NOX4 (1:50) antibody. The negative control slide was generated by omitting the primary antibody. The secondary antibody was incubated for one hour at room temperature, followed by a PBS wash. The colorimetric detection was carried out using a diaminobenzamide tetrahydrochloride peroxidase system (Dako; K069), and counterstained for 3 minutes with Harris hematoxilin. Tissue sections were observed and photographed using a Zeiss microscope.
Real-time polymerase chain reaction analysis
Total RNA was extracted from the rat thyroid and PCCL3 cells using the RNeasy® Plus Mini Kit (Qiagen), following the manufacturer's instructions. After DNAse treatment, reverse transcription was followed by real-time polymerase chain reaction (PCR), as previously described (17). The specific oligonucleotides listed in Table 1, were purchased from Applied Biosystems.
Antioxidant enzyme activity
Fresh thyroids were homogenized in 5 mM Tris-HCl buffer (pH 7.4), containing 9 mg/mL NaCl, 1 mM ethylenediaminetetraacetic acid followed by centrifugation at 750 g for 10 minutes at 4°C. The supernatant aliquots were stored at −70°C. Catalase activity was assayed following the method of Aebi and activity is expressed as units per milligram of protein (U/mg) (18). GPx activity was assayed by following NADPH oxidation at 340 nm in the presence of an excess of glutathione reductase, reduced glutathione and tert-butyl hydroperoxide as substrates (19), and expressed as nmol of oxidized NADPH per milligram of protein (nmol/mg).
Measurement of thiol groups
Total thyroid thiols were determined in a spectrophotometer (Hitachi U-3300) using 5,5-dithionitrobenzoic acid (DTNB). Thiols react with DTNB, cleaving the disulfide bond to give 2-nitro-5-thiobenzoate (NTB−), which ionizes to the NTB2− di-anion in water at neutral and alkaline pH. The NTB2− was quantified in a spectrophotometer by measuring the absorbance of visible light at 412 nm (20).
Statistical analysis
All the results are expressed as mean±standard error of the mean, and the differences between male and female groups were compared using the Student t test. The results of thyroid H2O2 generation during the estrous cycle, serum estradiol levels and mRNA levels and H2O2 generation in PCCL3 cells were analyzed by the one-way analysis of variance test followed by the Newman–Keuls multiple comparison test. Statistical analyses were done using the software GraphPad Prism (Version 5, GraphPad; Software Inc.) and the level of significance was established at p≤0.05.
Results
Thyroid H2O2 production and NOX mRNA levels
Adult female thyroid particulate fractions produce significantly higher amounts of H2O2 than male ones, as shown in Figure 1A, both in the presence (female: 46.8±2.3, male, 36.1±2.8 nmol H2O2/mg per hour) or absence (female: 31.2±2.6, male: 21.4±1.8 nmol H2O2/mg per hour) of calcium (p<0.01). Nevertheless, the calcium-dependent (DUOX) H2O2 production (activity in the presence deduced by the activity in the absence of calcium) did not differ between males and females. No sex differences were detected in H2O2 production by prepubertal rat thyroids (Fig. 1B). To evaluate which NOX family member was responsible for the higher H2O2 production in the adult female thyroid, DUOX1, DUOX2, NOX4, DUOXA1, DUOXA2, p22phox, and Poldip2 mRNA expression were analyzed by real-time PCR. NOX4 and Poldip2 mRNA expression were 1.5-fold higher, in adult female thyroids, as compared to adult males (Fig. 1C, D; p<0.05), but no significant differences were found in the other genes studied herein. Figure 1E shows a representative immunoperoxidase photomicrography for NOX4 in male and female thyroid tissue. NOX4 staining is present in the cytoplasm of follicular cells as well in the apical plasma membrane. The NOX4 immunostaining observed in slices of female thyroids is markedly stronger in comparison to male thyroids.
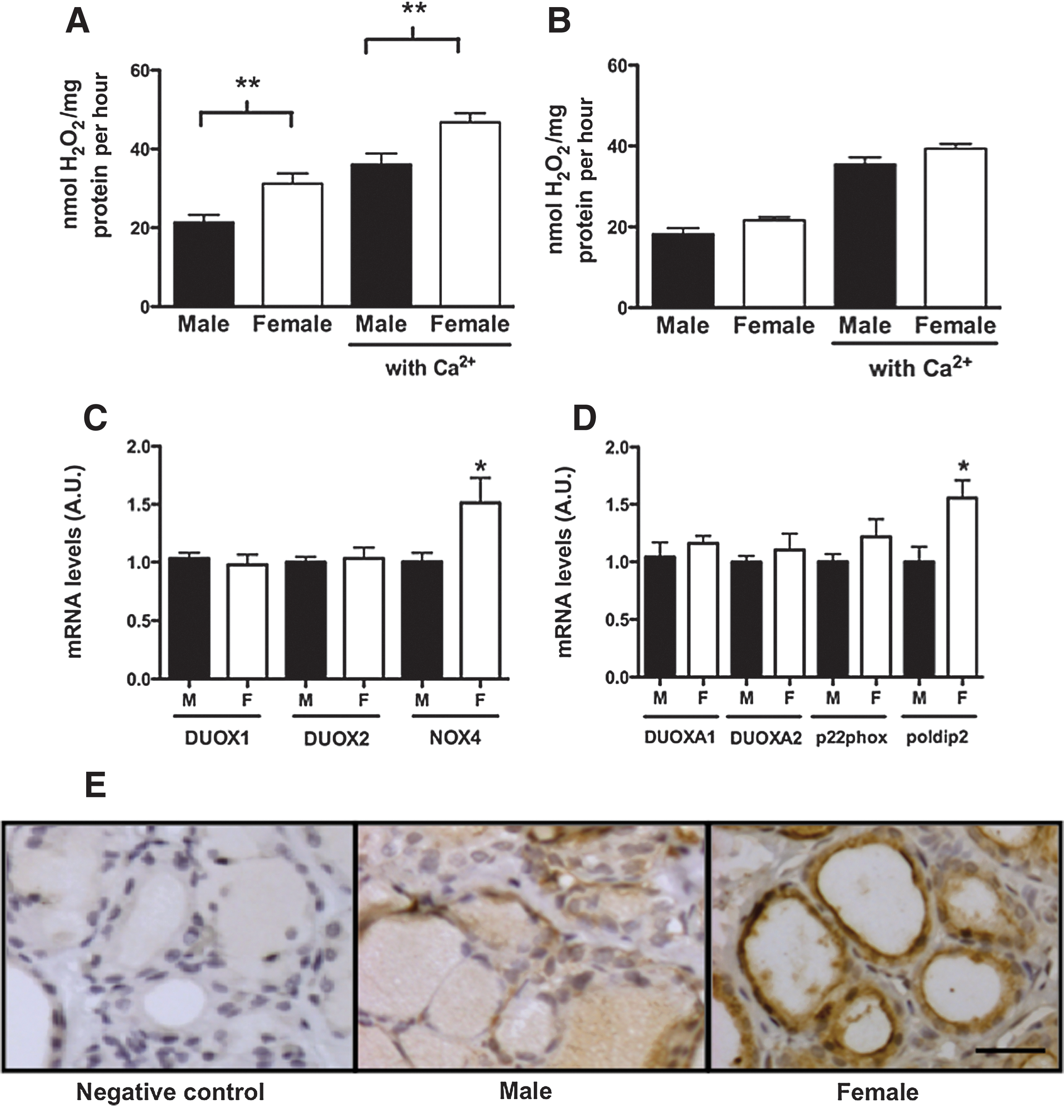
Sexual dimorphism in thyroid hydrogen peroxide (H2O2) production. H2O2 production was determined in male and female thyroid particulate fractions of adult
The greater NOX4 mRNA levels in the thyroid of adult females seem to be dependent on sexual maturation, since no differences in thyroid NOX4 mRNA levels were detected between prepubertal males and females (data not shown), and estradiol levels are significantly higher only in adult females (Table 2). Interestingly, the regulation of NOX4 is tissue specific since, in the liver (Fig. 2A) and kidney (Fig. 2B), NOX4 mRNA levels were higher in males than in females.

NOX4 mRNA levels in liver
All data are shown as mean±standard error of the mean.
p<0.01 versus the other groups, analysed by one-way analysis of variance.
The estrous cycle in female rats is characterized by transient changes in serum estrogen levels (21). H2O2 production was significantly higher in the estrus phase when compared with the other phases of the cycle (Fig. 3A), which was associated with higher NOX4 mRNA expression (Fig. 3C) and serum estradiol levels (Fig. 3B) in the proestrus, a phase that precedes the estrus phase.

H2O2 generation in thyroids obtained from female rats in the different phases of the estrous cycle (P=proestrus, E=estrus, D1=diestrus 1, D2=diestrus 2).
Antioxidant enzyme expression and activity
GPx1 mRNA expression (Fig. 4A) was 70% greater in the adult female than in the male rat thyroid (p=0.0001), but catalase mRNA expression (Fig. 4D) was decreased by 60% (p<0.05). There were no differences in the expression of the other genes studied (GPx2, GPx3, SOD1, 2, and 3). GPx and catalase activities were evaluated in thyroid homogenates of adult male and female rats. While catalase activity (Fig. 5A) in females was lower than in males (females: 1471±118.3 U/mg, males: 1913±64.2 U/mg; p=0.01), just as found for mRNA expression, GPx activity was not different between groups (Fig. 5B).
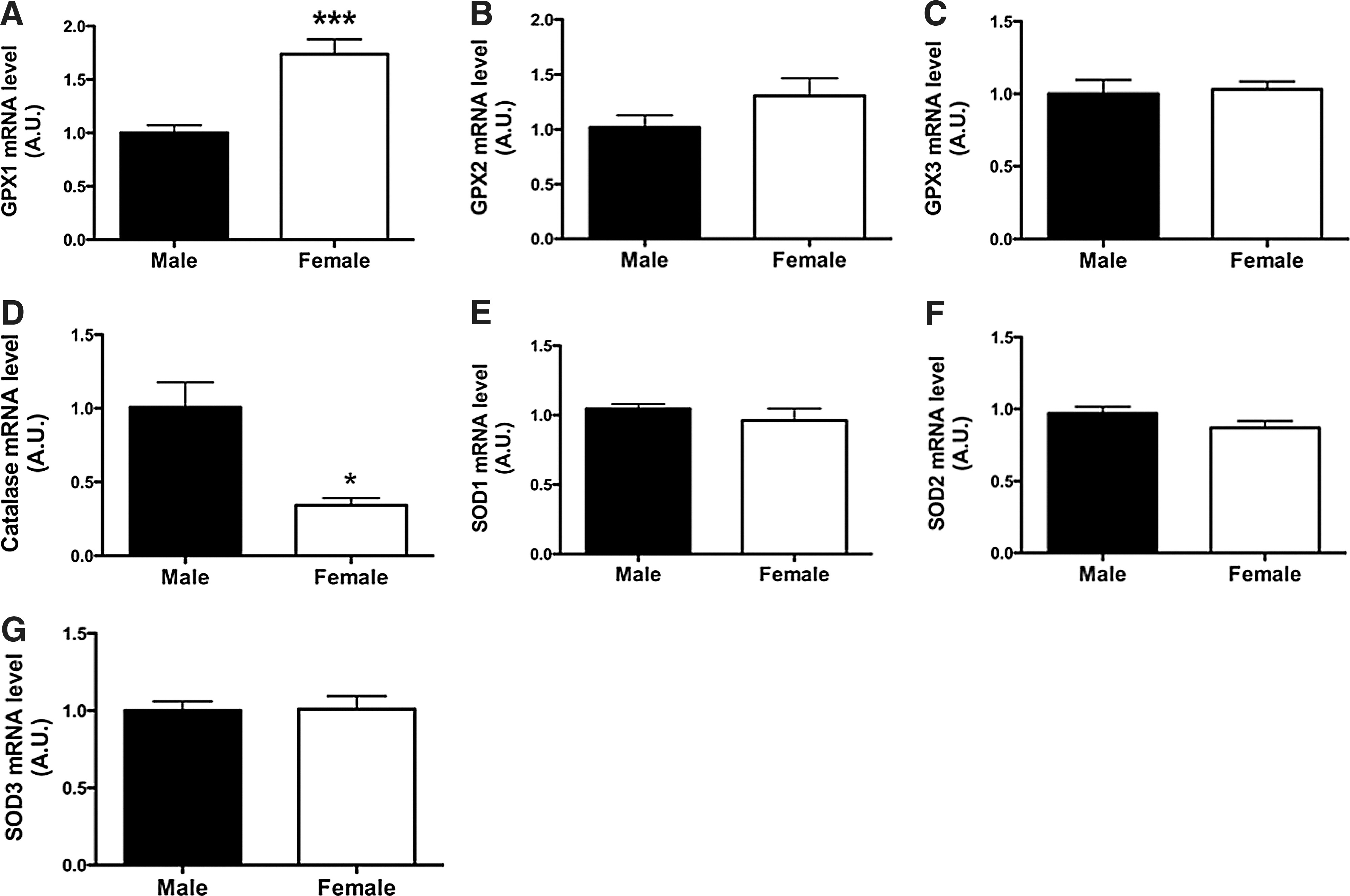
mRNA expression of antioxidant enzymes in the thyroids of male and female rats. Glutathione peroxidase 1 (GPx1)

Activities of antioxidant enzymes in the thyroids of male and female rats. Catalase
Thiol groups
As shown in Figure 6, adult female rat thyroids contain less thiol groups than male rat thyroids (male, 39.3±5.4 nmol reduced DTNB/mg; female, 23.2±3.8 nmol reduced DTNB/mg; p<0.05), confirming that the adult female rat thyroid is under higher oxidative stress levels.

Female rat thyroids are submitted to higher oxidative stress levels. Total sulfhydryl groups were measured by the reaction of thiols with 5,5-dithionitrobenzoic acid (DTNB), evaluated in a spectrophotometer at 412 nm. Results are expressed as mean±SEM (n=5), *p<0.05.
The role of estrogen in the NOX4-dependent thyroid oxidative stress
To elucidate the role of estrogen in NOX4 sexual dimorphism, we evaluated H2O2 generation and NOX4 expression in ovariectomized rats treated or not with physiological doses of Eb. No differences in H2O2 generation (Fig. 7A) and NOX4 mRNA levels (Fig. 7B) were detected among the groups treated for 7 days. However, 21-day-old ovariectomized rats had higher NOX4 mRNA expression (Fig. 7D) than sham-operated and OVX+Eb rats (C=1.0±0.11; OVX=1.65±0.16; OVX+Eb=1.43±0.10), but no differences were observed in thyroid particulate fractions from 21 days ovariectomized rats (Fig. 7C). These results indicate that factors other than estradiol might also regulate NOX4 expression in the thyroid. As we have previously shown (22), ovariectomized rats have a different pattern of circulating cytokines than sham-operated animals. To exclude interference by these molecules, we utilized an in vitro model, incubating PCCL3 cells with different 17β-E concentrations for 72 hours We found that PCCL3 cells express estrogen receptor α (ERα) and G-protein–coupled estrogen receptor (GPR30), but not estrogen receptor β (ERβ; data not shown), justifying the use of PCCL3 cells in estrogen-related experiments. Comparing ERα and GPR30 mRNA levels among male and female thyroids and PCCL3 cells, we observed that in PCCL3 cells the content of both receptors is much lower compared to the thyroid glands of both sexes (Fig. 8A, B). NOX4 mRNA levels (Fig. 8C) were four times higher at a 17β-E concentration of 10−7 M compared to control cells, but no significant differences were found for the other concentrations (control=1.0±0.09; 10−9 M 17β-E=1.4±0.29; 10−8 M 17β-E=1.6±0.22; 10−7 M 17β-E=3.8±0.84 A.U.; p=0.002). Poldip2 mRNA levels (Fig. 8D) were 2.5-fold higher in cells treated with 10−9 M 17β-E in comparison to control cells, and no significant differences were detected at the other concentrations used (control=1.0±0.07; 10−9 M 17β-E=2.5±0.54; 10−8 M 17β-E=1.7±0.30; 10−7 M 17β-E=0.8±0.14 A.U.; p<0.05). In PCCL3 cells, no p22phox mRNA was detected. Extracellular H2O2 generation was significantly higher in 10−7 M 17β-E–treated cells compared to the other groups (control=2.5±0.32; 10−9 M 17β-E=2.1±0.12; 10−8 M 17β-E=2.4±0.18; 10−7 M 17β-E=3.8±0.22 nmol H2O2/105 cells per hour; p<0.05; Fig. 8E).
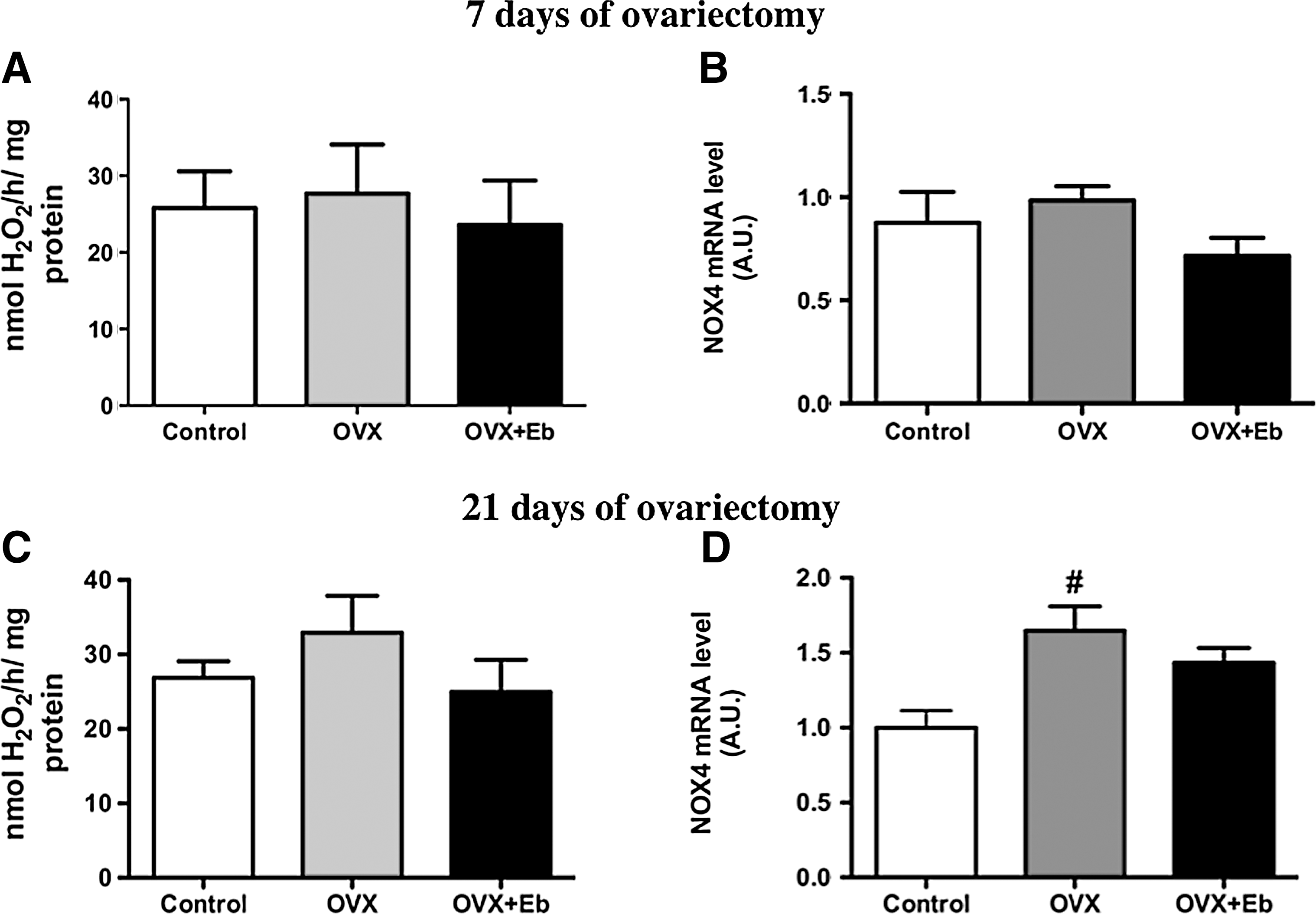
H2O2 generation and NOX4 mRNA expression in thyroids obtained from ovariectomized (OVX) rats treated or not with estradiol benzoate. H2O2 generation without calcium was determined in adult female thyroid particulate fraction of Sham (Control), OVX, and OVX plus estradiol benzoate 0.7 μg/100 g body weight (OVX+Eb) rats treated for 7 days
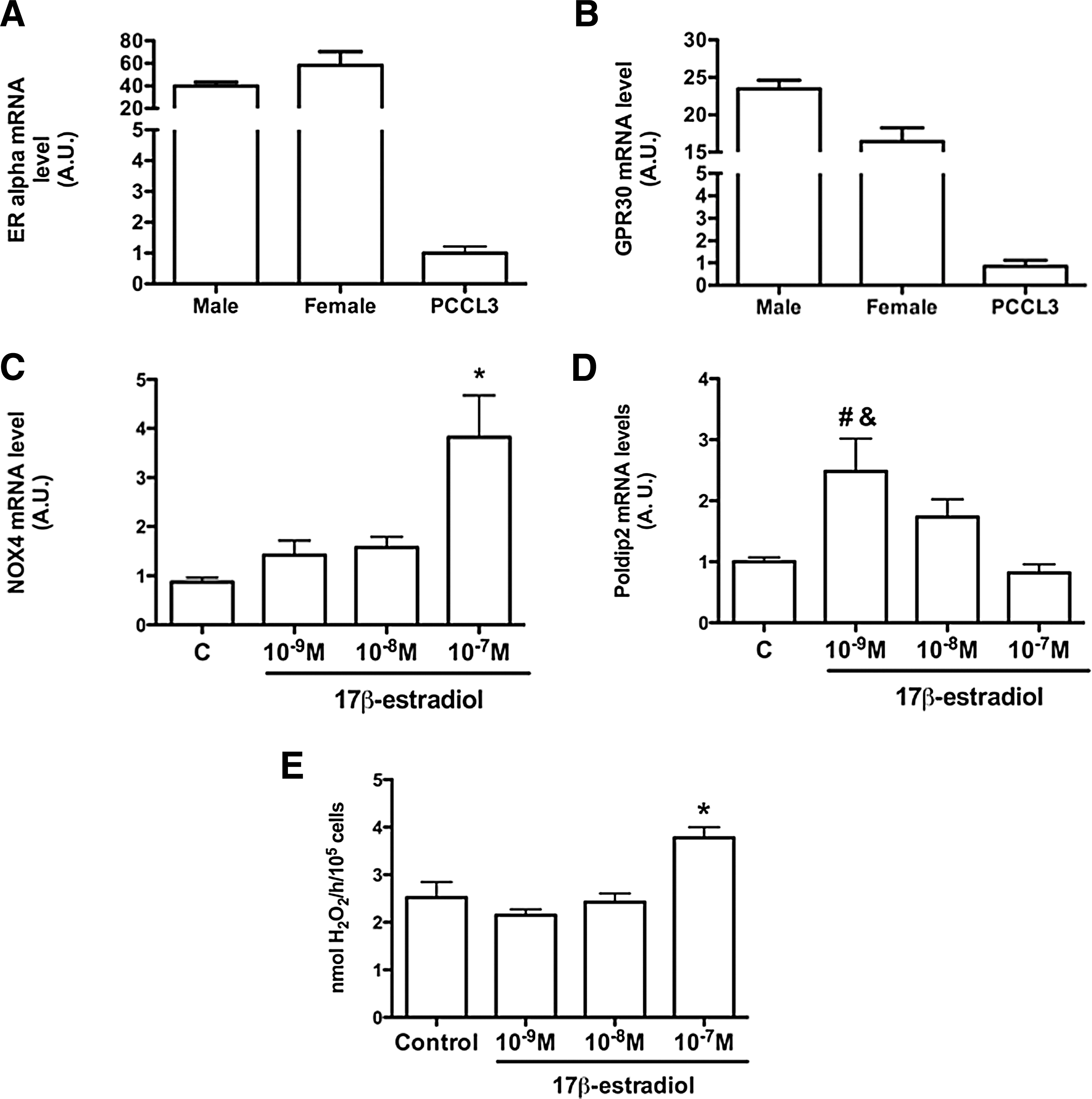
Role of estrogen in the NOX4-dependent thyroid oxidative stress. PCCL3 cells were grown until semiconfluence and incubated with 0.2% fetal bovine serum (FBS) for 48 hours, in the presence of six hormones. After that, the cells were treated with different 17β-estradiol doses in 0.2% FBS for 72 hours. Estrogen receptor α
Discussion
Thyroid diseases affect more than 27 million people in the United States (23). Among these, thyroid cancer is the most common endocrine malignancy accounting for nearly 95% of all endocrine cancers according to the American Cancer Society (24). The prevalence of thyroid dysfunction is five times higher in women than in men (25). Although sexual dimorphism in the prevalence of thyroid disorders is well described, the mechanisms that lead to this marked difference remain unknown.
H2O2 is produced by all cells of living organisms and is able to act in the redox-dependent regulation of different cellular functions, including response to stressors, angiogenesis, cell proliferation, and others (26). To maintain intracellular ROS at adequate levels, antioxidant systems react with these molecules and produce less reactive compounds. Conceptually, an imbalance between pro-oxidant compounds and antioxidant defenses leads to oxidative stress; this concept has recently been redefined as the “disruption of redox signaling and control” (27). At the apical membrane of thyrocytes, H2O2 acts as a TPO cosubstrate in thyroid hormone biosynthesis. The sources of thyrocyte H2O2 related to hormonogenesis are the DUOX enzymes, DUOX1 and 2, which have two calcium-binding sites that are indispensable for their activities (2,3). Another NOX, NOX4, has recently been described in thyrocytes; in contrast with DUOX, it generates ROS in the intracellular compartment and is constitutively active (4). DUOX1 and DUOX2 are active at the cellular plasma membrane only in the presence of the DUOX maturation factors: DUOXA1 and DUOXA2, respectively (5,28). On the other hand, NOX4 seems to depend on the p22phox protein to be fully active.
Recently, it was demonstrated that NOX4 and p22phox expression are upregulated in thyroid cancers, linking NOX4-dependent H2O2 generation to cancer development or progression (4). Herein, we show that in adult female rat thyroid particulate fractions, there is a higher calcium-independent H2O2 production together with a higher NOX4 mRNA expression. This sexual dimorphism was not found in prepubertal thyroids, indicating a direct role of sexual steroids on the NOX4 expression difference found between sexes. Consistent with this hypothesis, we found that in proestrus, the phase of the estrous cycle that is characterized by a peak in serum estrogen, NOX4 nRNA levels were higher in comparison to the other phases, and a higher H2O2 production was detected in the estrus phase. Moreover, NOX4 and Poldip2 mRNA levels were higher in PCCL3 cells incubated with 17β-estradiol, as well as the H2O2 production rate. NOX4 mRNA and H2O2 production were higher only in cells treated with 10−7 M of estrogen, which can be explained by the lower estrogen receptor expression level found in PCCL3 cells. Poldip2 was originally identified as the DNA polymerase δ-interacting protein, implicated in regulation of gene expression, DNA duplication, and DNA repair (29,30). Estrogen has well-described effects in thyrocyte proliferation. Thus, it is probably that Poldip2 could mediate the estrogen effects on cellular division, being more sensitive to estradiol activation than NOX4, once it is regulated in accordance to its functions.
Weyemi et al., (4) described that in normal human thyroid tissue, NOX4 immunostaining was intracytoplasmic. In the present study, we found that NOX4 immunostaining in the thyroid of both male and female rats was intracytoplasmic, but could also be detected at the plasma membrane of thyrocytes. This difference suggests a species-specific regulation of the subcellular distribution of NOX4.
Ovariectomy is usually a good model to evaluate the effects of systemic estrogen deficiency. However, some previous studies published by our group and others show that serum cytokines levels are modulated in ovariectomized rats (22). In addition, ovariectomized rats have a higher body mass 21 days after surgery in comparison to sham-operated rats. As it is well documented that serum TGF-β is increased in obese animals and IL-1β and TGF-β are potent stimulators of NOX4 expression in different tissues, ovariectomy might not be an adequate model to study the regulation of NOX4 expression by estrogen (22,31,32). These concepts are hypothetical as serum cytokines levels were not evaluated in this study.
To minimize the damaging effects of ROS, scavengers react with free radicals to convert them into nonradical oxidants. The enzymes catalase and GPx are specifically involved in H2O2 detoxification, producing H2O directly or in a glutathione-dependent reaction. Catalase has a much higher Michaelis constant (Km) value for H2O2 (25 mM) than the thyrocyte intracellular H2O2 concentration; thus, thyroid GPx seems to be more important for H2O2 degradation under physiological conditions. GPx1 is a cytosolic enzyme, while GPx3 is found in the thyroid colloid, and catalase is located in peroxisomes (5). The discrepancy between GPx mRNA expression and activity can be explained by the interaction between this enzyme and high H2O2 levels, since GPx can be irreversibly inactivated by its own substrate, due to modification of selenocysteine residues at its active site (33). Thus, female thyrocytes could have more GPx mRNA expression than their male counterparts to maintain GPx activity. In our work, we found higher thyroid H2O2 production in female than male rats; although the activity of catalase was decreased in female thyroids and GPx did not differ between groups, indicating sexual dimorphism in redox balance, where female rat thyroids might be exposed to an oxidative stress condition due to increased H2O2 generation and decreased catalase activity.
H2O2 has been linked to some processes related to the development of thyroid nodules and cancer, such as cellular proliferation, DNA damage, and apoptosis inhibition. Normally, H2O2-dependent regulation occurs through redox modification of thiol groups. This process is implicated in the post-translational regulation of protein functions, and thus is involved in macromolecular interactions, regulation of enzymatic activity, and others (34). Our finding of the lower thiol content in the thyroids of adult female than male rats indicate that more H2O2 is available to react with cellular constituents. Salmeen et al. recently showed that NOX4 expression is involved in cell cycle entry through inactivation of the p53-dependent checkpoint pathway, utilizing NOX4-silenced human fibroblasts (35). The authors suggest that the activation of p53 could be due to oxidation of its cysteine residues by NOX4-generated H2O2, decreasing its DNA-binding activity. Moreover, some protein tyrosine phosphatases, including phosphatase and tensin homologue, have cysteine catalytic residues that can be oxidized by ROS, leading to their inactivation (36). All the H2O2 actions described above reinforce the hypothesis that female thyroids are chronically exposed to higher levels of H2O2 than males and, thus, a potential to cause macromolecular damage, a higher cell proliferation rate, and inhibition of apoptosis, all of which might be associated with increased risk for thyroid disorders.
In conclusion, we demonstrate the existence of a sexual dimorphism in the thyrocyte redox balance, characterized by a higher H2O2 production, due to higher NOX4 and Poldip2 expression, and decreased enzymatic antioxidant defense in the thyroids of adult female rats compared to adult male rats. Our novel findings might contribute to an explanation for the molecular basis for the higher prevalence of thyroid disorders in women; increased H2O2 levels might well be one of the factors underlying the increased risk of thyroid disease.
Footnotes
Acknowledgment
This work was supported by Fundação de Amparo à Pesquisa do Rio de Janeiro (FAPERJ).
Disclosure Statement
No competing financial interests exist.