
Abstract
Background:
Thyroid disorders are associated with an increased risk of cognitive impairment and Alzheimer's disease. Both small vessel disease and neurodegeneration have a role in the pathogenesis of cognitive impairment and Alzheimer's disease. Thyroid hormone receptor alpha (TRα) is the predominant TR in brain. The circadian clock gene REV-ERBα overlaps with the TRα gene and interferes with TRα expression. Limited data are available on the role of the TRα/REV-ERBα locus in small vessel disease and neurodegeneration. We therefore studied genetic variation in the TRα/REV-ERBα locus in relation to brain imaging data, as early markers for small vessel disease and neurodegeneration.
Methods:
Fifteen polymorphisms, covering the TRα/REV-ERBα locus, were studied in relation to white matter lesion (WML), total brain, and hippocampal volumes in the Rotterdam Study I (RS-I, n=454). Associations that remained significant after multiple testing correction were subsequently studied in an independent population for replication (RS-II, n=607).
Results:
No associations with total brain or hippocampal volumes were detected. A haplotype block in REV-ERBα was associated with WML volumes in RS-I. Absence of this haplotype was associated with larger WML volumes in women (0.38%±0.18% [β±SE], p=0.007), but not in men (0.04%±0.11%, p=0.24), which was replicated in RS-II (women: 0.15%±0.05%, p=0.04; men: 0.05%±0.07%, p=0.80). Meta-analysis of the two populations showed that women lacking this haplotype have a 1.9 times larger WML volume (p=0.001).
Conclusion:
Our results suggest a role for REV-ERBα in the pathogenesis of WMLs.
Introduction
The actions of the active TH T3 (3,5,3′-triiodo-L-thyronine) are mediated through binding to nuclear TH receptors (TRs), thereby regulating gene expression. TRα is the predominant receptor in the brain (4). In mice, a knock-in mutation in TRα leading to a lower affinity to T3 results in, besides a bone and metabolic phenotype, memory impairment in adulthood (5).
On the opposite chromosomal strand of TRα, the circadian clock gene REV-ERBα is located. These genes partially overlap and REV-ERBα expression has been shown to influence splicing of TRα (6 –8). Given that circadian rhythm abnormalities have been associated with cognitive impairment and Alzheimer's disease (9), it is of interest to study genes involved in the circadian clock.
To date, limited data are available on the role of the TRα/REV-ERBα locus in the mature human brain, and in cognitive impairment and Alzheimer's disease in particular. In recent years, it has been shown that both neurodegeneration and small vessel disease have a role in the pathogenesis of cognitive impairment and Alzheimer's disease (10 –13). Therefore, we studied genetic variation in the TRα/REV-ERBα locus in relation to (early) markers of small vessel disease and neurodegeneration derived from MR brain imaging data. White matter lesion (WML) volume was used as a marker for small vessel disease (14), and hippocampal and total brain volumes were used as markers for neurodegeneration (15 –18). The associations of the TRα/REV-ERBα locus with WML, hippocampal, and total brain volumes were studied in a population-based cohort study. Associations that remained significant after multiple testing correction were tested in an independent population for replication.
Materials and Methods
Participants
The Rotterdam Study I (RS-I) is a prospective population-based cohort study from 1990 onward in 7983 Caucasians aged ≥55 years, aimed at investigating determinants of various chronic diseases among elderly persons (19). In 1995, a structured interview, physical examination, blood drawing, and brain magnetic resonance imaging (MRI) were performed in a random subset of 536 nondemented subjects of RS-I.
In 1999, RS-I was expanded (RS-II) with 3011 subjects who had become 55 years of age or moved into the study district. In 2005, a structured interview, physical examination, blood drawing, and brain MRI scans were performed in a random subset of 895 nondemented subjects of RS-II.
The medical ethical committee of the Erasmus MC, University Medical Center, Rotterdam, approved both studies and all participants gave written informed consent.
MRI measures
The Rotterdam Study I
Brain scans were performed on a 1.5 T MRI System (VISION MR; Siemens AG, Erlangen, Germany). In 490 participants we obtained a proton-density, a T2-weighted, and a high-resolution inversion-recovery double contrast 3D HASTE sequence for multi-spectral volumetry (15,20). Image preprocessing and automated measurements of WML and total brain volume have been described in detail previously (15). Hippocampal volumes were measured based on manual segmentations (15).
The Rotterdam Study II
Brain scans were performed in 895 participants on a 1.5 T MRI System (General Electric Healthcare, Milwaukee, WI) (21). For all participants, a T1-weighted, proton-density, and FLAIR sequence were acquired. Preprocessing of these images and the automated measurement of WML, total brain, and hippocampal volume have been described in detail previously (21,22).
Thyroid hormone measurements
In RS-I, blood samples were collected at the time of MRI. Serum TSH (thyroid-stimulating hormone), FT4 (free 3,5,3′,5′-tetraiodo-L-thyronine), and T3 levels (n=470) were measured with chemoluminescence assays (Vitros ECI Immunodiagnostic System; Ortho-Clinical Diagnostics, Rochester, MI).
Selection and genotyping of polymorphisms
Based on linkage disequilibrium (LD) analysis (
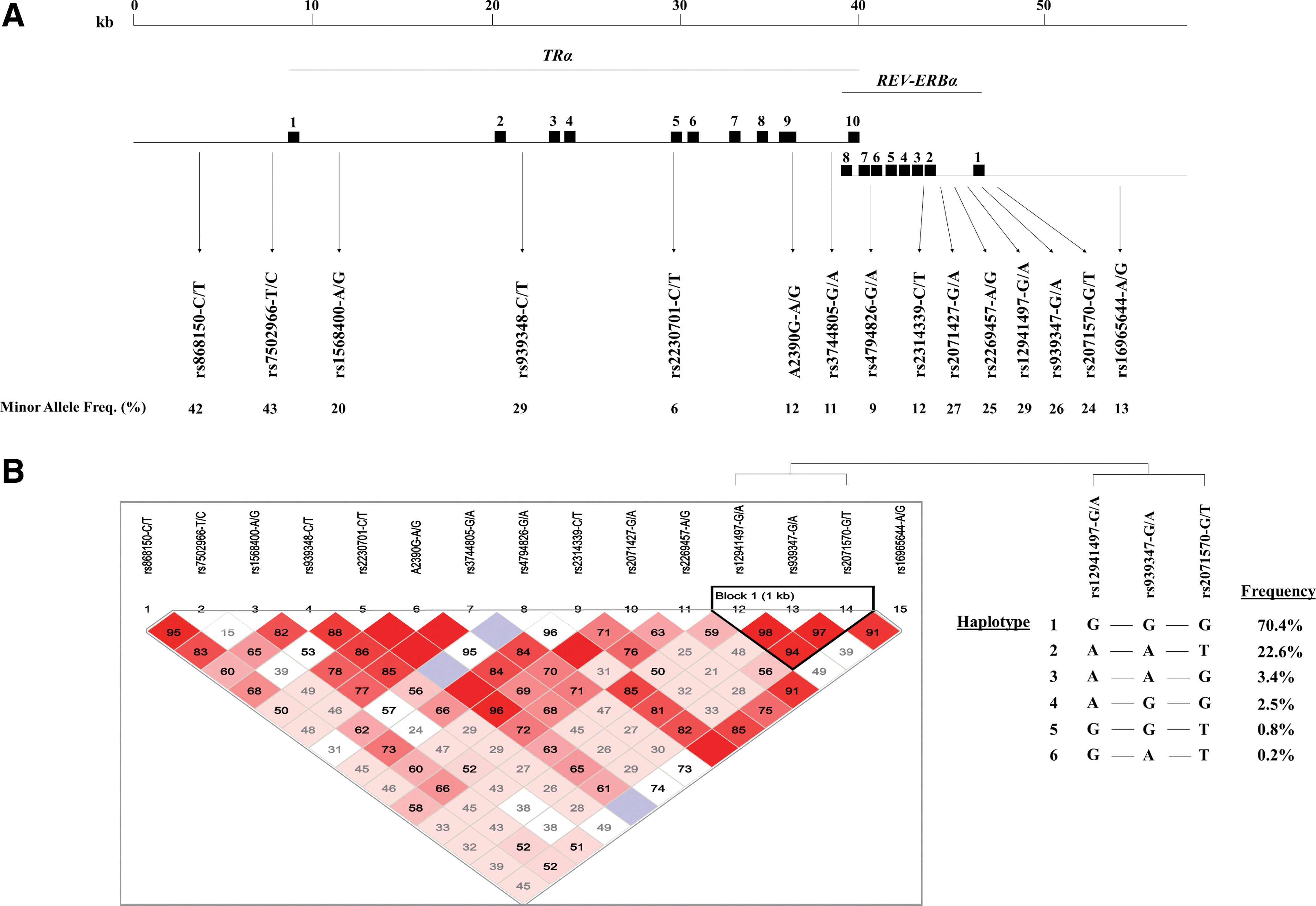
For RS-I and RS-II, genotypes were extracted from the Illumina HumanHap 550K (Duo) array. Genotypes for rs2230701 and A2390G were determined with Taqman Allelic Discrimination (Applied Biosystems, Nieuwerkerk a/d IJssel, the Netherlands). Genotype data were available in 454 (RS-I) and 607 (RS-II) subjects with imaging data.
Using genotype data from 470 subjects from RS-I, the LD structure of the TRα/REV-ERBα locus was analyzed using Haploview 4.1 (24).
Statistical analysis
Genotype and allele frequencies were tested for Hardy–Weinberg equilibrium. Linear regression was used to compare baseline characteristics between genotype groups. WML, total brain, and hippocampal volumes were expressed as percentage of total intracranial volume to adjust for head size differences. WML volume was additionally natural log transformed because of skewness of the untransformed measure. The associations with WML, total brain, and hippocampal volumes were tested using linear regression. All analyses were adjusted for age and gender. To minimize the risk of false-positive findings, multiple testing correction by permutation analysis was performed, thereby taking the LD structure between these polymorphisms into account. Results were obtained after 10,000 permutations, using PLINKv1.07 (25). As REV-ERBα is a circadian clock gene and gender-related differences in circadian rhythm regulation have long been recognized (26 –29), we investigated the gender-specific effects of REV-ERBα polymorphisms that remained significant after multiple testing correction at p=0.05.
Associations that remained significant after multiple testing correction at p=0.05 in RS-I were tested in RS-II for replication. Meta-analyses were conducted using the METAL software package applying inverse-variance weighted fixed-effects methodology (
Power calculations for detectable effect sizes in RS-I and RS-II combined, and in RS-I alone were performed at β=0.80 and α-values corresponding to the multiple testing corrected p-value thresholds.
In RS-I and RS-II, we had power to detect differences in WML, total brain, and hippocampal volumes of 0.27, 0.20, and 0.17 standard deviation (SD), for polymorphisms with an MAF of 10%, 20%, and 30%, respectively. Similarly, in RS-I alone, we had power to detect differences of 0.41, 0.31, and 0.27 SD. One SD WML volume equals 1.53% and 0.66% in RS-I and RS-II, respectively. One SD hippocampal volume equals 0.10% and 0.05% in RS-I and RS-II, respectively. Similarly, 1 SD total brain volume equals 3.66% and 3.41%.
Results
Allele and genotype frequencies of all polymorphisms were in Hardy–Weinberg equilibrium with similar frequencies as reported in literature (23) and established databases, such as HapMap (
Both RS-I and RS-II consisted of 51% women. Mean ages were 73.4±7.9 (mean±SD) and 67.5±5.5 years, respectively.
None of the studied polymorphisms were associated with baseline characteristics, including serum TSH, FT4, and T3 levels.
In RS-I, REV-ERBα-rs939347-A was associated with larger WML volumes (β=0.26%±0.10% (mean±SE), p=0.002), which remained significant after multiple testing correction (p=0.021) (Table 1). As this polymorphism is located in a region of high LD, haplotypes defined by REV-ERBα-rs12941497, −rs939347, and −rs2071570 were created (“Block 1” in Fig. 1). Absence of haplotype 1 was associated with larger WML volumes (β=0.20%±0.10%, p=0.007). We additionally investigated the gender-specific effects of haplotype 1 on WML volumes. This effect was largely driven by women (women: β=0.38%±0.18%, p=0.007; men: β=0.04%±0.11%, p=0.24) (Fig. 2). This effect was replicated in RS-II, which also showed a significant association with larger WML volumes in women (β=0.15%±0.05%, p=0.041), but not in men (β=0.05%±0.07%, p=0.795) (Fig. 2). Meta-analysis of the two populations resulted in β=0.17%±0.05% (p=0.001) in women, and in β=0.05%±0.06% (p=0.42) in men. None of the other studied polymorphisms were associated with WML (Table 1) or total brain volumes (data not shown).
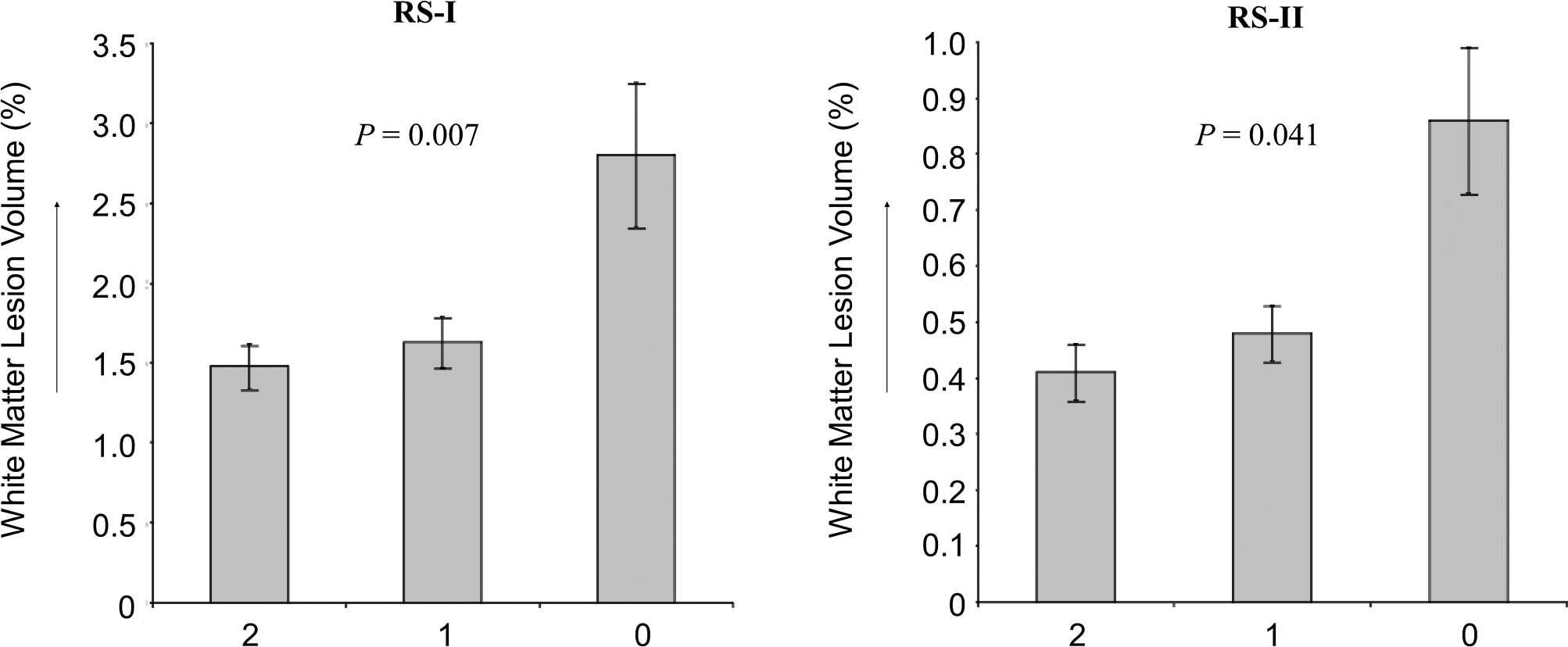
White matter lesion volumes by number of REV-ERBα haplotype 1 copies in 218 women from RS-I and 293 women from RS-II. Volume is expressed as percentage of intracranial volume to adjust for head size differences. Meta-analysis of the two populations resulted in β=0.17%±0.05% (p=0.001).
Expressed as white matter lesion volume percentage. Volume is expressed as percentage of intracranial volume to adjust for head size differences. Effects are adjusted for age and gender.
Obtained after multiple testing correction by permutation analysis (10,000 permutations).
In RS-I, TRα-A2390G-G was associated with smaller hippocampal volumes (β=−0.03%±0.01%, p=0.002), which remained significant after multiple testing correction (p=0.027). However, this effect could not be replicated in RS-II (β=0.01%±0.01%, p=0.28). Meta-analysis of the two populations resulted in β=−0.02%±0.05% (p=0.63). None of the other studied polymorphisms were associated with hippocampal volumes (data not shown).
Discussion
In the present study, we investigated the effects of genetic variation in the TRα/REV-ERBα locus on WML, total brain, and hippocampal volumes. A haplotype block covering exon 1 of the REV-ERBα gene was associated with larger WML volumes. REV-ERBα is a nuclear hormone receptor with a key role in the regulation of the circadian rhythm, which is generated by feedback loops of gene expression (31). In this system, REV-ERBα acts as a constitutive repressive transcription factor, as it has an atypical ligand-binding domain lacking the carboxy-terminal activation function-2, required for recognition of co-activators (32). WMLs, presumed to result from cerebral small vessel disease, range from reduced myelination and edema to gliosis and complete axonal destruction (14). WMLs are associated with a substantial increased risk of cognitive decline, dementia, stroke, and death (33). In the present study, the association of the REV-ERBα haplotype was exclusively driven by its association in women. We show that women lacking REV-ERBα haplotype 1 have a 1.9 times larger WML volume compared with women with 1 or 2 copies of this haplotype (Fig. 2). Gender differences in circadian rhythm regulation have long been recognized (26 –29). Barger et al. found differences in the circadian timing system of body temperature, heart rate, physical activity, and feeding between male and female rhesus monkeys (26). In humans, others have shown gender differences in the circadian rhythms of body temperature and sleep regulation as well (27). Also, at the level of the individual clock components, a number of studies have shown gender differences in circadian rhythm regulation. For example, the type of depression in relation to variants in the clock gene TIMELESS is dependent on gender (34). Recently, Hadden et al. studied the effects of circadian disruption on mouse lung mechanics, and demonstrated that the effects on the lungs, as well as the changes in REV-ERBα expression patterns, were different between men and women (35).
Taken together, various studies have shown that the regulation of circadian clock genes, as well as the effects of dysregulation of those genes, including REV-ERBα, can differ between genders. However, no studies are available on the gender-specific effects of REV-ERBα on the pathogenesis of WML. The exact mechanism behind the gender-specific effects of REV-ERBα on WML therefore needs to be clarified in future studies.
The associated haplotype block in REV-ERBα covers exon 1 and the promoter region of the gene, and may therefore influence splicing or the transcriptional level of REV-ERBα. In addition to a direct effect of the REV-ERBα haplotype, the effects of this haplotype on WML volumes may also be mediated via TRα. As can be expected from the genomic organization of the TRα/REV-ERBα locus (see Fig. 1), REV-ERBα transcription also influences splicing of TRα (6 –8). There are two major TRα isoforms, the T3-binding TRα1 and the non-T3-binding TRα2, which has an antagonistic function. Base pairing with REV-ERBα mRNA blocks splicing of TRα2 mRNA, thereby favoring formation of TRα1 mRNA. In this way, REV-ERBα expression influences the TRα1/TRα2 ratio, thereby regulating local T3 action (6 –8).
Recently, the first three patients with a mutation in TRα have been described (36,37). Patients suffered from growth retardation, as well as from motor and cognitive dysfunction. However, no brain imaging data were available in these patients.
Little is known about the exact role of circadian clock components in the pathogenesis of WMLs. Our results suggest a role for the circadian system, and for REV-ERBα in particular, in the pathogenesis of WMLs, the exact molecular mechanism of which needs to be clarified in future studies. In this context it is interesting to note that circadian rhythm disturbances are frequently observed in patients with Alzheimer's disease, and even in nondemented patients with the earliest signs of Alzheimer's neuropathology (9).
Genetic variation in TRα has previously been studied in relation to Alzheimer's disease, which did not reveal significant associations (38). This is in line with the results of the present study, which do not show an association of genetic variation in TRα with early markers of neurodegeneration or small vessel disease.
Strengths of our study include the high coverage of genetic variation in the studied locus. In addition, due to the relatively large sample size, we were powered to detect at least moderate differences in WML, total brain, and hippocampal volumes. However, we cannot exclude other potential (small) effects of low-frequency polymorphisms.
A point of concern in genetic association studies is the risk of false-positive findings. To minimize this risk, we applied both a multiple testing correction and replicated significant results in an independent population. Further, the relation between the REV-ERBα haplotype 1 and WML volume was similar in RS-I and RS-II: absence of both haplotype 1 copies was associated with higher WML volumes, whereas carriage of only one haplotype 1 copy was not (Fig. 2). It is therefore highly unlikely that these observed effects are false-positive findings.
In conclusion, we have shown that genetic variation in the circadian clock component REV-ERBα is associated with WML volumes in women. Future studies are needed to clarify the exact role of the TRα/REV-ERBα locus, and the circadian rhythm system in general, in the pathogenesis of WMLs. Given the close relation between TRα and REV-ERBα, these studies should identify the independent contributions of REV-ERBα and TRα to the observed effects on WMLs.
Footnotes
Acknowledgments
This work was supported by ZonMw VENI Grant 91696017 (RPP) and an Erasmus MC Fellowship (RPP). The Rotterdam Study was funded by Erasmus Medical Center and Erasmus University, Rotterdam; the Netherlands Organization for Scientific Research (NWO), Netherlands Organization for Health Research and Development (ZonMW); the Research Institute for Research in the Elderly (RIDE); the Ministry of Education, Culture and Science; the Ministry for Health, Welfare and Sports; the European Commission (DG XII); and the municipality of Rotterdam. The authors are grateful to the participants and staff from the Rotterdam Study, the participating general practitioners, and the pharmacists.
Disclosure Statement
No competing financial interests exist.