
Abstract
Background:
Approximately 60% of sporadic medullary thyroid carcinomas (sMTC) remain orphan of a recognized genetic cause. Recently, a high percentage of RAS point mutations have been described in RET-negative sMTC. The aim of this study was to assess the prevalence of RAS point mutations in a large series of MTC collected in four Italian centers.
Methods:
For this purpose, we studied codons 12, 13, and 61 of H-, K-, and N-RAS genes in 188 MTC samples, either hereditary or sporadic, by direct sequencing. Correlations between the RAS mutational status and the clinical-pathological features of MTC patients as well as a meta-analysis of all published data were performed.
Results:
The prevalence of RAS mutations in the present series of MTC was 10.1%, and 17.6% when considering only RET-negative cases. RAS mutations were found in MTC tumoral tissue, but not in peripheral blood indicating their somatic origin. A novel mutation in codon 72 (M72I) was found, but with a low or null transforming potential. No association was found between the presence of RAS mutations and the clinical-pathological features of the patients. Although not statistically significant, a positive association between the presence of RAS mutations and a better outcome was observed. The meta-analysis of all published studies confirmed a prevalence of 8.8% for RAS mutations in MTC.
Conclusions:
The prevalence of RAS mutations in our MTC series was relatively low and consistent with the meta-analysis data. Only somatic RAS mutations were found and only in RET-negative sMTC. Likewise, MTCs that harbor a RAS mutation identify a subgroup of tumors with less aggressive behavior. To our knowledge, this is the largest series of MTCs studied for the presence of mutations in RAS genes and the first meta-analysis on this specific topic.
Introduction
The human RAS genes encode for three distinct but highly homologous 21 kDa proteins (H-RAS, K-RAS, and N-RAS) that serve as transducers of signals from cell membrane receptors to intracellular effectors such as RAS-RAF-MAPK and RAS-PI3K-AKT pathways, which control a wide range of effects on cellular proliferation, differentiation and apoptosis. Activating point mutations in RAS genes confer constitutive activation of multiple downstream effectors and have been extensively described in several tumor types (13,14).
Point mutations of the RAS genes, particularly N-RAS, are usually detected in tumors arising from the thyroid follicular epithelium of the thyroid both in benign tumors, such as in follicular adenoma, and in malignant tumors, such as follicular thyroid carcinoma and the follicular variant of papillary thyroid carcinoma (PTC) (15 –17). Nevertheless, RAS mutations have been also found in MTC, which arise from thyroid parafollicular C-cells, though with discordant results. In particular, point mutations in the RAS genes have been previously studied in small series of sMTC and were either not detected (18 –24) or reported in only a few cases (25 –28). The only considerably high prevalence of K-RAS mutations (40.9%) was previously found in a Greek series (29). Unexpectedly, H-RAS and K-RAS mutations have been recently found in ∼68% of RET-negative sMTC and in only 2.5% of RET-positive cases, suggesting that RAS and RET somatic mutations are likely mutually exclusive in MTC tumorigenesis (30).
The aim of the present study was to evaluate the prevalence of mutations in the three RAS genes in a large Italian multicentric series of MTC, either hereditary or sporadic. The presence of the mutations was also correlated with the presence of the somatic RET mutation and the clinical-pathological features of the patients. Finally, the overall prevalence of RAS mutations in MTC has been calculated with a meta-analysis that included all of the data published thus far together with the present results.
Materials and Methods
MTC patients and clinical data
A total of 188 MTC (13 hereditary and 175 sporadic) cases were enrolled in four different Italian centers: Department of Endocrinology and Metabolism, University of Pisa (n=102), Department of Surgical, Oncological, and Gastroenterological Sciences, University of Padua (n=61), Department of Medical Sciences, University of Milan (n=18), and Experimental Oncology Department-U.O. Molecular Mechanisms, IRCCS Foundation, Istituto Nazionale dei Tumori, Milan (n=7). All 13 hereditary cases were collected at the University of Pisa. Clinical data of MTC patients were retrieved from a computerized database collecting epidemiological, medical, and pathological features of all patients affected by MTC available in the four centers involved in this study. Part of the series from Padua (n=34) was previously reported in another publication (31).
Of the 188 MTC patients, 81 were male and 107 female with a mean age of 54.5±15.6 years (median 57 years, range 19–87 years) at diagnosis. Clinical and pathological data were not available for 12/188 (6.4%) of cases. According to the tumor-node-metastases (TNM) classification (32), 48/176 (27.3%) were at stage I, 38/176 (21.6%) at stage II, 43/176 (24.4%) at stage III, and 47/176 (26.7%) at stage IV.
According to their clinical status, defined on the basis of both serum calcitonin levels and imaging results (i.e., neck ultrasound, computerized tomography scan, and bone scintigraphy) as previously described (11,33), MTC patients were distinguished into two groups: (i) “disease-free” patients [n=89/180 (49.4%)]; (ii) patients with “persistent disease,” either at the biochemical level or with evidence of metastatic disease [n=74/180 (41.1%)] and dead patients [n=17/180 (9.4%)]; in 8/188 (4.3%) of cases, data were not available because the patients were lost to follow-up.
At the time of the present study, the mean follow-up was 78.52±60.87 months, ranging from 0 to 240 months (median, 60 months).
RET gene mutation analysis in MTC tumor samples
Tissue samples were collected at surgery, snap-frozen, and stored at −80°C until usage. The diagnosis of hereditary or sMTC was routinely assessed after clinical evaluation, familial history, and RET gene mutation analysis of peripheral blood and tumor tissue of exons 5, 8, 10, 11, 13, 14, 15, and 16 as described elsewhere (34). Informed consent for the genetic analyses was obtained from the patients. The present study was approved by the internal reviewing boards of the four centers involved.
Mutational analysis for the H-, K-, and N-RAS genes in the MTC samples
The RAS mutational analysis was performed on genomic DNA that was extracted from tissues, amplified by polymerase chain reaction (PCR) and directly sequenced. All the cases were sequenced using both forward and reverse primers previously used for amplification. The PCR conditions were separately designed in each center to amplify exons containing hot spots for mutations (i.e., codons 12, 13, and 61) of each RAS gene. No major differences were found by comparing the PCR conditions used in the four centers. Positive and negative controls were present in each experiment. Each positive case was further confirmed with a second separate PCR. Whenever possible, the mutations detected were also studied in the peripheral blood DNA from the same patient to confirm the somatic nature of the mutation.
In silico analysis of RAS mutations
The in silico analysis of the RAS mutations was performed using the Align-Grantham Variation-Grantham Deviation (Align-GVGD) web-based program (
Meta-analysis of the prevalence of RAS gene mutations in MTC
To perform the meta-analysis of the prevalence of the RAS mutations in MTC, we searched for all articles that were published until 2012 in online databases (PubMed, Google Scholar). After filtering for redundancy, we obtained 13 articles (two abstracts and 11 full-text articles) (18 –30). Since not all series analyzed all RAS isoforms, we calculated the prevalence of mutations in each RAS isoform (H-, K-, and N-RAS) by pulling together data obtained in all series and the overall prevalence of the RAS mutations using the data of the seven studies in which all cases were studied for all RAS isoforms (18,20 –22,26,27,30). A comparison with the prevalence of RAS mutations in our series was also performed.
Statistical analysis
Statistical analysis was performed using StatView 5.0.1. software (SAS Institute Inc., Cary, NC). A Chi-squared (χ 2) test and unpaired Student's t-test were used to compare the clinical and pathological features of the patients and the RAS mutational status. Differences were considered statistically significant when the p-value was less than 0.05.
Results
RET mutational status
A germline RET point mutation was found in 11/13 (84.6%) patients with hereditary MTC, and among these, a mutation at codon C634 was found in 7/13 (53.8%) patients (three C634Y, three C634F, and one C634R); a V804M mutation was present in 2/13 (15.4%) patients; a E768D mutation was revealed in 1/13 (7.7%) patients, and a M918T mutation was found in 1/13 (7.7%) patients. Two out of thirteen (15.4%) patients with hereditary MTC did not show any RET point mutations.
A somatic RET point mutation was detected in a total of 69/175 (39.4%) sMTC. Among the cases with a somatic RET mutation, 48/69 (69.7%) harbored the M918T substitution: 15/69 (21.7%) had a mutation at codon C634 (two C634W, four C634Y, seven C634R, one C634S, and one C634G); 3/69 (4.3%) patients showed a complex A883F mutation; 1/69 (1.4%) patients presented contemporary two mutations (Q781R+M918T) and 2/69 (2.9%) patients carried the deletion p.D898_E901del(c.2694_2705del12). Finally, 106/175 (60.6%) patients with sMTC did not show any RET gene somatic mutation in the sequenced exons.
RAS mutational status
An overall prevalence of 10.1% for RAS mutations was found in the series of MTCs studied (19/188; Fig. 1A). The prevalence was similar in the single series belonging to the four different centers (data not shown). None of the germline and somatic RET-positive MTC cases (n=80) showed any mutations in any of the RAS genes. As shown in Figure 1B, mutations in the RAS genes were found in 19/108 (17.6%) RET-negative MTC cases. In detail (Table 1), H-RAS mutations were found in 13/108 (12.1%) cases, with one at codon 12 (G12R), one at codon 13 (G13R), 10 at codon 61 (three Q61K, five Q61R, and two Q61L), and a novel H-RAS mutation, the ATG to ATA substitution, coding for M72I. K-RAS mutations were found in 4/108 (3.7%) cases, with three at codon 12 (G12R) and one at codon 61 (Q61H). Finally, N-RAS mutations were detected in 2/108 (1.8%) cases, with both mutations at codon 61 (1 Q61R and 1 Q61L). Both patients that harbored N-RAS mutations in their MTC presented with a concomitant PTC.
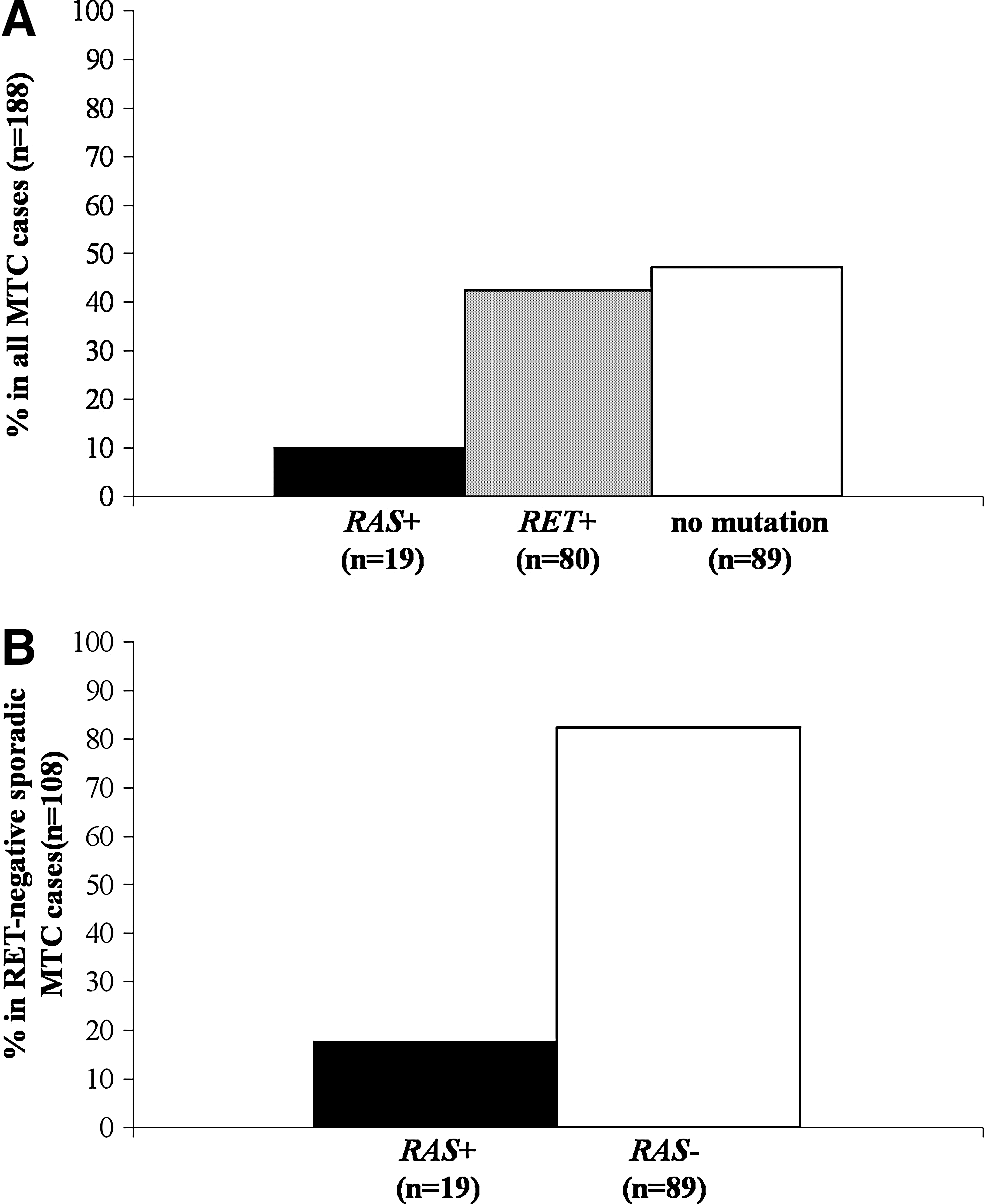
Overall prevalence of RET and RAS mutations in the entire MTC series
Novel H-RAS mutation.
Sequence analysis also revealed some synonymous nucleotide polymorphisms: 10 at codon 27 of H-RAS (H27H) and two at codons 13 and 56 of K-RAS (one G13G and one L56L).
Correlations between the RAS mutational status and the clinical and pathological features in RET-negative MTC
The RAS mutational status in RET-negative sMTC was correlated with the clinical and pathological parameters of the patients. As shown in Table 2, no statistically significant difference was observed between the presence of a RAS mutation and the age, sex, TNM category, size of the primary tumor, stage of the disease, and outcome of the patients with MTC. However, through an examination of the percentage of disease-free patients who harbor RAS mutations and comparing the percentage to disease-free patients with no RET/no RAS mutations, a higher but not significant prevalence of disease-free patients was found in the mutated RAS group (Fig. 2). Conversely, the outcome of patients with a somatic RET mutation was significantly worse than the outcome of both RAS positive/RET negative and RAS negative/RET negative cases (p<0.0006 and p<0.01, respectively; Fig. 2).
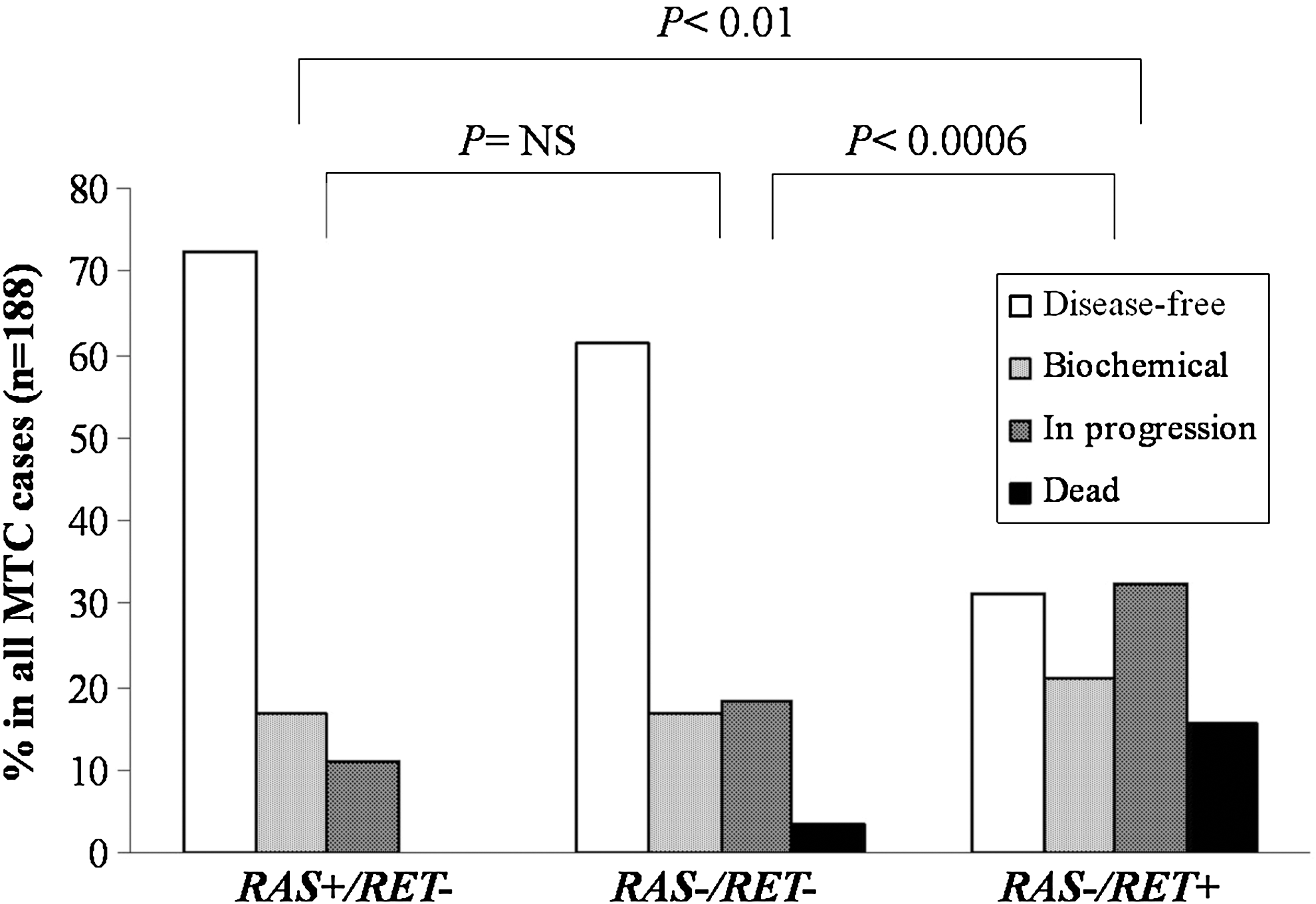
Correlation between the RAS-positive/RET-negative (RAS+/RET−), RAS-negative/RET-negative (RAS−/RET−), and RAS-negative/RET-positive (RAS−/RET+) patients and their outcomes. The p value is indicated.
As continuous variables analyzed with unpaired t test.
Unless stated, values are number/total number (%).
p Values analyzed with χ 2 test.
SD, standard deviation.
In silico analysis of RAS mutations
As shown in Table 3, the in silico analysis showed that the H-RAS G12R, G13R, and Q61L substitutions scored in class C65, the H-RAS Q61K substitution scored in class C45, the H-RAS Q61R scored in class C35, and M72I in class C0. The K-RAS G12R and Q61H substitutions scored in class C65 and C15, respectively. Finally, the N-RAS Q61R substitution scored in class C35. All of the mutations fell in the “deleterious 1” category according to the GD/GV prediction (37).
Novel mutation.
GD, Grantham deviation; GV, Grantham variation.
Meta-analysis of the prevalence of RAS mutations in MTC
According to the meta-analysis and when considering all 14 studies (including the current series), the prevalence of mutations in the H-, K-, and N-RAS was 32/393 (8.1%), 26/397 (6.5%), and 2/359 (0.5%), respectively (Table 4). The overall prevalence of RAS mutations calculated based on data obtained from the eight studies (including the current series) reporting a complete analysis of the mutational status was 30/338 (8.8%; Table 5). Moreover, when comparing the overall prevalence of RAS mutations reported in the present study (10.9%) and those derived from the other seven complete series (12.9%), we did not observe any statistically significant difference by χ2-test.
Only codon 61 was studied.
In one case H-RAS analysis was not performed.
Only codons 12 and 13 were studied.
na, not available; ns, not studied; PCR, polymerase chain reaction; DS, direct sequencing; PM, primer-mismatch analysis; SSCP, single strand conformation polymorphism; ASH, allele specific hybridization; PIREMA, primer introduced restriction with enrichment of mutant alleles; RFLP, restriction fragment length polymorphism; PYRO, pyrosequencing.
Discussion
The only recognized genetic alterations responsible for the development of MTC are point mutations of the RET gene (2). Despite this finding, 40–60% of sporadic cases and 2% of familial cases do not have a known genetic driver. Point mutations in the RAS genes have been extensively described in several tumor types (14), including tumors arising from the follicular epithelium of the thyroid (15 –17). RAS mutations have been also studied in MTC, though those studies were performed in a limited number of cases with discordant results, making this topic quite controversial (Table 4). H-RAS and K-RAS mutations have only recently been described in a large percentage of RET-negative sMTC (30) calling attention again to a possible role of these mutations in the development of MTC and when considering future strategies for targeted therapies.
In the present Italian multicentric study, we have investigated the prevalence of H-, K- and N-RAS point mutations in a vast series of hereditary and sMTC collected by four Italian centers to assess the prevalence of these mutations in an Italian large cohort. To this purpose, we studied 188 cases of MTC and found that 19/108 (17.6%) of RET-negative sMTC harbored RAS mutations. None of the RET-positive MTC, either hereditary or sporadic, presented with a concomitant RAS somatic mutation (p<0.0001) thus confirming the previous observation that these two genetic events are mutually exclusive (30).
The prevalence of the RAS mutations was lower than previously reported in the Greek (29) and Portuguese (30) series (40.9% and 68%, respectively). Conversely, our results appear to be more in line with other studies (25 –28) and with the overall prevalence that emerged from the meta-analysis we performed (8.8%), as shown in Table 4. Although the reason for the different results obtained in these series is unclear, we have some hypotheses to explain this. A first explanation could reside in the different methodology that was used for the RAS mutation screening. It has been previously demonstrated that in thyroid follicular neoplasia, the detection of RAS mutations by direct sequencing was less frequent than by using other more sensitive methods (16). This could explain the high prevalence reported by the Greek study, which was performed using enriched PCR followed by fragment length polymorphism analysis (26), but not the data of the Portuguese series (27), which used the same methodology, PCR, and direct sequencing, as used in the present study. Other possible explanations can be offered to explain these differences, such as the different genetic backgrounds among Greek, Portuguese, and Italian individuals, different exposure to environmental factors and carcinogens and the limited number of cases that were analyzed in the previous two series.
To get more insight into the difference in the prevalence of RAS mutations among series, we performed a meta-analysis of the available data in the literature. Unfortunately, some of the published studies did not provide information on RET mutational status (20,22
–25,28,29), while others did not screen for all three RAS genes (23,24,29). For this reason, we calculated the overall prevalence of RAS mutations in sMTC without distinguishing the RET-positive and RET-negative cases, and found an overall prevalence of ∼12.9%, which is slightly higher than ours (10.9%) but much lower than that described in the two previously mentioned series (29,30). In our meta-analysis we kept the prevalence of mutations affecting the three RAS isoforms separated due to lack of homogeneity in the studies. We found that in MTC the percentage of mutations in H-RAS (8.1%) is higher than in K-RAS (6.5%), while mutations in N-RAS remain a rare event (0.5%). Notably, the variability in the prevalence of RAS oncogene mutations in different reports represents a general problem. This issue has already been analyzed for RAS mutations in tumors arising from thyroid follicular lineage (16), for RET/PTC rearrangements (38) and for the BRAF
V600E mutation (39,40) in PTC. To overcome this problem, a computerized database of the overall frequencies of all known oncogenes, which was obtained by calculating the mean of all frequencies reported in the literature, is available on the internet (
Mutations in the N-RAS gene appear to be extremely rare in MTC, which is in contrast to that for tumors arising from thyroid follicular cells (15 –17). However, Takahashi et al. demonstrated that C-cell adenomas occurring in Rb1+/− mice progress to metastatic MTC after loss of N-RAS concluding that in C-cells N-RAS may act more as a tumor suppressor than a proto-oncogene (41). In our series, the two patients with mutations in the N-RAS gene had a PTC in association with the MTC. A frequent association between the two types of tumors has been already described (42,43) and was never fully explained. The identification of N-RAS point mutations in MTC associated with a PTC suggests an interesting scenario that requires further study.
All of the studies present in the literature, except for those not showing the RET mutational status, showed a mutual exclusivity between RET and RAS mutations in MTC. We confirmed this in our study also. In the 80 cases with RET mutations (11 with a germline and 69 with a somatic mutation), we did not find any RAS mutation. A precise role for RAS mutations in the development of RET-negative MTC can be postulated. While the pathogenic role of RET mutations in MTC development has been clearly demonstrated both in vitro and in vivo (2), no similar data have been published for RAS mutations. The transforming ability of the known RAS gene mutations has been definitively demonstrated in follicular cells for thyroid cancer and in several other types of human tumors (13). In this regard, the in silico analysis that we performed in the present study confirmed that all of the RAS mutations described belong to the deleterious Category 1 because they fall in conservative regions of the gene, including the newly reported RAS mutation. Nevertheless, not all of these substitutions show a similar protein-disrupting power, with G12R, G13R, and Q61L of H-RAS and G12R of K-RAS being the most relevant, as expected. Conversely, according to the alignment GD/GV class (37), the newly M72I mutation detected in H-RAS appears to affect protein function to a lesser extent and consequently seems to have a low transforming power.
In one of our previous studies, we hypothesized that the RET-negative MTC, which is more prevalent in small tumors (<2 cm), could represent a subgroup of MTC in which RAS mutations could possibly play a role (44). Nevertheless, in the present study, we were not able to find any statistically significant association between the presence of RAS mutations, tumor size, and other epidemiological and pathological features.
Similarly to what had been observed in the Portuguese series (30), we observed a slightly higher prevalence of RAS mutations in disease-free patients when compared with patients without RAS or RET mutations. This finding suggests that a better prognosis can be expected in the RAS-positive MTC patient group. Therefore, RAS mutations could be responsible for a tumoral transformation of parafollicular C-cells, from which MTC arises, that leads to a less aggressive tumor phenotype; on the other hand, RET mutations could be responsible for a more aggressive phenotype that is associated with a worse prognosis (11). Recently, it has been observed that the doubling time of the serum levels of both carcinoembryogenic antigen and calcitonin can predict the behavior of MTC (45). Although this is a very useful parameter in predicting the outcome of MTC patients, it can change over time, being longer at the beginning of the disease and becoming shorter at later stages. In contrast, the mutational status of the primary tumor is an unchangeable parameter that could represent a prognostic factor for the final outcome.
The RAS analyses performed in peripheral blood in MTC cases harboring a RAS mutation in the tumor tissue was negative, indicating the somatic origin of the mutation. We also studied the genomic DNA of two hereditary RET-negative MTC cases, but both of them were negative for all of the RAS mutations analyzed. Indeed, this result was expected because germline H-RAS mutations are associated with Costello's syndrome, which is a severe disorder affecting many parts of the body and causing an intellectual disability, and none of the members of these two families had this syndrome. Conversely, germline K-RAS mutations are responsible for Noonan's syndrome, which is associated with dwarfism and is more common than Costello's syndrome. The range and severity of its features can vary greatly, and the syndrome is not always identified at an early age (46).
In conclusion, we found that 19/188 (10.1%) MTC were positive for RAS mutations, which were only detected in RET-negative sMTC (17.6%) and confirmed that the two events are mutually exclusive. Moreover, RAS mutations were found in MTC tumoral tissue but not in peripheral blood, indicating their somatic origin. A novel mutation (M72I) was identified in the H-RAS gene, although in silico analysis indicated that the transforming activity of this mutation was likely null. Finally, on the basis of our results, we can hypothesize that MTCs harboring a RAS mutation may identify a subgroup of tumors with a less aggressive behavior. To our knowledge, the present study is the largest series of MTCs analyzed for the presence of RAS gene mutations and the first meta-analysis on this specific topic.
Footnotes
Acknowledgments
This study was supported in part by grants from “Ministero della Istruzione Universitaria e Ricerca Scientifica (MIUR),” “Associazione Italiana per la Ricerca sul Cancro (AIRC),” and “Ministero della Salute, Progetto Ricerca Oncologica RF-CAM 2006353005.” The Department of Endocrinology and Metabolism, University-Hospital of Pisa is a WHO Collaborating Centre for the Diagnosis and Treatment of Thyroid Cancer and other Thyroid Diseases. We thank Dr. M. Moura who kindly provided genomic DNA for RAS mutated controls. We also thank Dr. P. Piaggi who revised the statistical analysis.
Disclosure Statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.